The CYP303A1 is essential in embryonic development of Nilaparvata lugens Stål (Hemiptera: delphacidae)
Yingzhen Liu, Zihan Yan, Di Deng, Huaiqi Wang, Ziyue Mao, Enrui Tang, Junyi Zhong, Xudong Zhao, Linquan Ge

TL;DR
This study shows that CYP303A1 is crucial for the embryonic development of the rice pest Nilaparvata lugens, affecting egg hatchability and embryogenesis.
Contribution
The study reveals a novel role of CYP303A1 in embryonic development, not previously linked to reproduction in this pest species.
Findings
Silencing CYP303A1 prolonged the embryonic period and reduced egg hatchability in N. lugens.
CYP303A1 knockdown caused abnormal embryonic development, including delayed eyespot formation and dispersed yolk granules.
Expression of hatching-related genes was significantly altered when CYP303A1 was silenced.
Abstract
The N. lugens is a highly fecund rice pest that causes severe damage to rice production in Asia. Cytochrome P450 monooxygenases (P450s) play essential roles in insect development, metabolism, and reproduction. In a previous study, we identified and functionally characterized CYP303A1, a member of the CYP2 family, in N. lugens molting and metamorphosis process. However, it is unclear the function of CYP303A1 in N. lugens fecundity. In this study, the expression profiling showed that CYP303A1 is highly expressed in legs and adult fat bodies, with moderate expression in the ovaries. Silencing of CYP303A1 in females had no significant effects on Vg (Vitellogenin) and VgR (Vitellogenin receptor) transcript levels, ovarian morphology, oocyte number, oviposition period, or female lifespan, indicating that CYP303A1 is not involved in vitellogenesis or ovarian development. However, knockdown of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
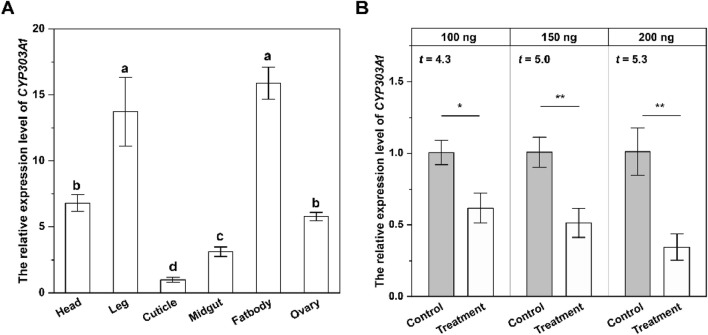 FIGURE 1
FIGURE 1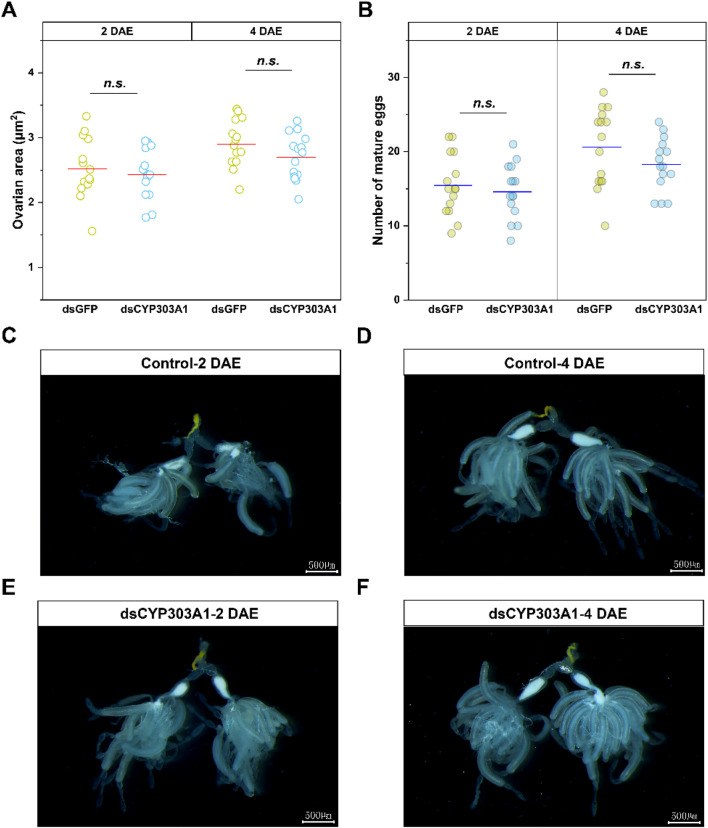 FIGURE 2
FIGURE 2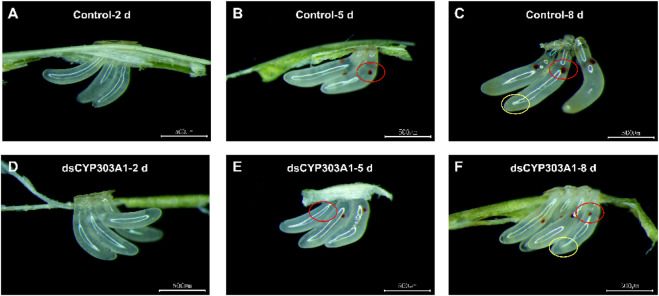 FIGURE 3
FIGURE 3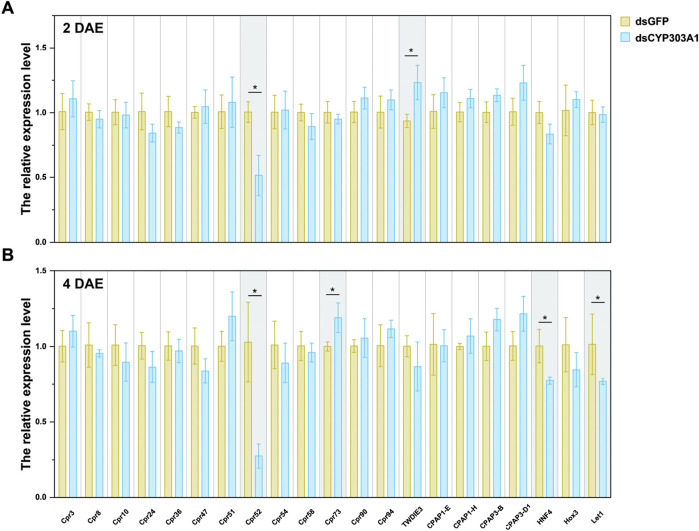 FIGURE 4
FIGURE 4| Reproductive and population parameters | dsGFP | dsCYP303A1 |
|---|---|---|
| Number of eggs laid | 241.2 ± 41.9 | 246.8 ± 47.4 |
| Preovipositon periods | 4.0 ± 0.7 | 4.1 ± 0.7 |
| Ovipositon periods | 15.0 ± 1.3 | 14.3 ± 1.3 |
| Female longevity | 18.7 ± 1.2 | 18.5 ± 1.2 |
| Egg durations | 9.7 ± 0.8 b | 14.3 ± 1.5 a |
| Hatching rate (%) | 83.3% ± 2.0% a | 26.2% ± 10.3% b |
| Sex ratio (female to male) | 1.2 ± 0.1 | 1.2 ± 0.1 |
| Population growth index (PGI) | 100.5 ± 17.8 a | 32.0 ± 3.6 b |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect Resistance and Genetics · Insect-Plant Interactions and Control · Neurobiology and Insect Physiology Research
1 Introduction
Cytochrome genes encoding P450 enzymes are consistently found among the most prominent gene families in plants, animals and fungi (Dermauw et al., 2020; Nauen et al., 2022). Cytochrome P450 enzymes (CYP), commonly called CYP450s, play a crucial role in essential biological processes across a diverse range of organisms (Dermauw et al., 2020). In insects, these enzymes significantly impact physiological processes such as development, reproduction, detoxification, and resistance to pesticides (Nebert and Dalton, 2006; Feyereisen, 2012). Moreover, CYP450s are at the interface of environmental responses, metabolism, and endocrine regulation by catalyzing the transformation of a myriad of exogenous and endogenous substrates by hydroxylation, epoxidation, dealkylations and a great variety of other reactions (Ye et al., 2022; Tzotzos, 2025). In insects, CYP genes are mainly categorized into six clans: clan 2, clan 3, clan 4, and mitochondrial clan with two new additions clan 16 and clan 20, which are restricted to certain Apterygotes and Paleoptera with unknown functions (Dermauw et al., 2020). Most of the single-copy genes belong to the CYP2 clan and the mitochondrial CYP clan, and most of the multiple-copy paralogs belong to the CYP3 and CYP4 clans and are often arrayed in clusters on chromosomes (Feyereisen, 2011; 2012). Many closely paralogous genes are part of lineage-specific family expansions, called P450 blooms (Sezutsu et al., 2013; Hafeez et al., 2022). The P450s implicated in xenobiotic metabolism and pesticide resistance are often found in such blooms.
The CYP3 and CYP4 clans are generally predominant in insects and relate to insect chemical defence. Considerable evidence has shown that P450s of the CYP3 clan, represented by members of CYP6 and CYP9 families, are mainly involved in xenobiotic metabolism by direct detoxification (Sezutsu et al., 2013; Nauen et al., 2022). In addition, members of the CYP4 clan (in the CYP4G subfamily) participate in this process by adjusting cuticle penetration through biosynthesis of cuticular hydrocarbons (Balabanidou et al., 2016; Feyereisen, 2020). In contrast, the evolution and function of most P450s in CYP2 and mito clans are considered highly conserved, and they form many families with few or even single members. Definite evidence has linked P450s in these two clans to biosynthesis or metabolism of endogenous compounds in model insect species. The CYP2 in insects represents a highly conserved group of enzymes that play essential roles in multiple physiological processes. Functionally, CYP2 family members are primarily involved in the biosynthesis and metabolism of endogenous hormones, particularly ecdysteroids and juvenile hormones, which are critical for regulating insect molting, metamorphosis, reproduction, and development (Guittard et al., 2011; Truman, 2019; Jin et al., 2023; Wu et al., 2023). Several key CYP2 genes, such as CYP307A1 (Spook), CYP306A1 (Phantom), CYP302A1 (Disembodied), CYP315A1 (Shadow), and CYP314A1 (Shade), constitute the core of the so-called Halloween gene cluster and catalyze successive steps in the ecdysteroid biosynthetic pathway (Peng et al., 2019; Zhou et al., 2020; Yan et al., 2023). In addition to the previously defined Halloween genes, CYP18A1, which belongs to the CYP2 clan, is involved in 20E inactivation (Rewitz et al., 2010; Guittard et al., 2011). CYP301A1, which belongs to the mitochondrial CYP clan, was described as an important gene involved in the formation of the adult cuticle, but its biochemical function is unknown (Sztal et al., 2012). Another CYP2 clan member, CYP303A1 (nompH) is expressed in the socket cells of sensory bristles in D. melanogaster, and is essential for the development and structure of external sensory organs (Willingham and Keil, 2004). CYP303A1 is a strongly supported clade with generally a single gene for each species, but it is duplicated in the Argentine ant, Linepithema humile, and in the carpenter ant Camponotus floridanus where the two genes are in a tandem array. It is also duplicated in the damselfly C. splendens. CYP303A1 was not found in genomes or transcriptome shotgun assembly (TSA) beyond winged insects, and CYP303A1 is mostly a single copy gene, “stable” in insects (Fallon et al., 2018; Dermauw et al., 2020).
The brown planthopper, N. lugens Stål (Hemiptera: Delpahacide) is one of the most destructive insect pests in Asian rice-growing regions (Wu J. et al., 2020; Sun et al., 2024). It damages rice through direct phloem feeding and transmits Southern rice grassy stunt virus (RGSV), and rice ragged stunt virus (RRSV), leading to plant stunting, reduced tillering, and severe yield losses (Zhang et al., 2022; Zhu et al., 2023). In recent decades, the extensive use of chemical insecticides has not only disrupted natural enemy populations but also accelerated the evolution of insecticide resistance in N. lugens (Wei et al., 2009; Gao et al., 2025). Furthermore, its high reproductive capacity, strong migratory behavior, and adaptability to different rice cropping systems have made its management increasingly challenging (Wu J. et al., 2020; Zhao et al., 2025a). These factors highlight the urgent need to develop sustainable and ecologically sound control strategies against this pest. In our previous study, we found that CYP303A1 was crucial in N. lugens molting and metamorphosis process. Silencing of CYP303A1 disrupted the synthesis of 20E in nymphs, caused downregulation of the 20E signaling pathway, and further affected the transcription of cuticular proteins and chitin metabolism, which ultimately affected the shedding of the old epidermis and the formation of the new epidermis (Wu et al., 2024). Ecdysteroids are considered as the classic insect hormones involved in oogenesis of insects (Song and Zhou, 2020; Benrabaa et al., 2022; Schellens et al., 2022). However, it is unclear whether CYP303A1 affects the embryonic development of N. lugens female adults. In this study, we systematically investigated the key role of CYP303A1 in the reproduction of N. lugens female adults, that it could serve as a potential RNAi target for green control of N. lugens, and making it an important target for both fundamental research and the development of novel pest control strategies.
2 Materials and methods
2.1 Insect culture
The N. lugens populations were originally obtained from the China National Rice Research Institute (CNRRI, Hangzhou, China). The strains were routinely reared on rice seedlings (Wuyunjing 23) under temperature (26 °C ± 2 °C), relative humidity (80% ± 10%), and photoperiod (light: dark, 16:8 h) as previously described by Wu et al. (2024).
2.2 The tissue-specific expression analysis of CYP303A1 in N. lugens females
The tissue samples (including the head, leg, cuticle, fatbody, and ovary) were dissected from virgin female adults at 2 days after emergence under a binocular microscope (Leica EZ4, Germany) by sterilized scalpel and tweezers on ice. Each sample was collected in triplicate from 30 females. The total RNA of each tissue sample was isolated using the RNA Easy Fast Tissue/Cell Kit (Tiangen, Beijing, China) according to the manufacturer’s instructions. The First-strand cDNA was synthesized using the PrimeScriptTM 1st Strand cDNA Synthesis Kit (TaKaRa, Dalian, China) according to the manufacturer’s instructions. The real-time quantitative PCR (RT-qPCR) was performed in CFX96 Touch Real-Time PCR Detection System (Bio-Rad Co., Ltd., CA, United States) in 10 μL reaction mixtures containing 5 μL 2× SYBR Premix EX TaqII Master Mix (TaKaRa, Dalian, China), forward and reverse primers of 0.2 μL (10 nM), 1 μL cDNA template and 3.6 μL ddH_2_O. The RT-qPCR of thermal cycling conditions were as follows: 95 °C for 4 min, 35 cycles of 95 °C for 10 s, 58 °C for 30 s, 72 °C for 20 s, with a final extension of 72 °C for 10 min. Subsequently, a melting curve analysis was conducted in the 60–95 °C temperature range to verify the consistency and specificity of each reaction product. RT-qPCR primers of CYP303A1 were designed online (https://www.primer3plus.com/) and listed in Supplementary Table S1. The β-actin was used as a reference gene (Ge et al., 2020), and the relative gene expression was calculated using the 2^−ΔΔCT^ method (Livak and Schmittgen, 2001).
2.3 The dsRNA synthesis and microinjection
The double-stranded RNA (dsRNA) was synthesized by amplifying a CYP303A1 with a 333 bp fragment by T7 RNA polymerase promoter-linked primers, according to Wu et al. (2024). The Green Fluorescent Protein (GFP) was a negative control. All dsRNAs were synthesized using a T7 RiboMAX™ Express RNAi System (Promega, Madison, WI). Templates for the dsRNA synthesis were reacted in a thermal cycler (BIO-RAD, Hercules, CA, United States) following the procedure: 35 cycles at 95 °C for 30 s, 60 °C for 30 s, and 72 °C for 45 s, with a final extension at 72 °C for 10 min. PCR products were used as templates for dsRNA synthesis through purification kits (Novozymes Biotechnology, Nanjing, China), and the synthesized product was stored at −80 °C until use.
After dsRNA synthesis, the newly-emerged N. lugens females were anesthetized with CO_2_ and then injected with 100 ng, 150 ng, and 200 ng of dsCYP303A1 or dsGFP in the mid-thorax of female adults using a Naoject II microinjection device (Drummond Scientific, PA, United States) under a microscope. Then, the surviving females were transferred into a plastic cup (15 × 18 cm) containing rice plants at a 4-leaf stage (Zhu et al., 2023). Previous studies have demonstrated that N. lugens is susceptible to RNAi, and the silencing effect of dsRNA to target genes remained stable and highly efficient at 48 h, 72 h, 96 h, and even up to 120 h post-injection (Ge et al., 2015; Li et al., 2025). Therefore, the female adults were collected at 48 h post-injection of dsRNAs as a representative time point to examine the efficiency of RNAi. Each treatment contained at least 5 individuals and three independent biological replicates.
2.4 The isolation and observation of female adult ovaries
The ovaries of N. lugens dsRNAs-injected females were isolated and observed at 2 days (2 DAE) and 4 days (4 DAE) after emergence. The female adults were anesthetized with CO_2_ and subsequently dissected in 0.9% saline solution, and the fat bodies around the ovary were stripped cleanly. The isolated ovaries were washed and photographed using a microscope with a digital camera (Olympus, model SZX23, Japan). The count of ovarioles was documented using microscopic examination. There were 15 females used for each treatment.
2.5 Determine the reproduction and population parameters of N. lugens
The N. lugens reproductive parameters, including pre-oviposition periods, oviposition periods, and the number of eggs laid, were determined, referring to Ge et al. (2020), with slight variations. After 200 ng dsRNAs injected into newly newly-emerged females, the surviving females were paired 1:2 with untreated males (dsCYP303A1♀ × Control♂ or dsGFP♀ × Control♂) and were reared in glass tubes (2.5 cm diameter, 15 cm height) containing tillering rice stems. After the offspring (F1 generation) of N. lugens had reached the 3^rd^ instar, the number of offspring was counted and transferred to new rice stems, which were placed in glass cups and fed until emergence. After that, the unhatched eggs laid by the F0 generation on the rice stems were recorded. The hatching rate was calculated as offspring/offspring + unhatched eggs. The population growth index (PGI) was calculated as F_1_/F_0_, with F_1_ representing the total offspring of the following generation and F_0_ representing the number of parents (F0 = 4) (Zhao et al., 2025b). The rice stems were replaced every 24 h during the pre-oviposition period and every 48 h during the oviposition period until the females died. The pre- and oviposition periods and the number of eggs laid by females were recorded, and 15 replicates were used for each treatment.
2.6 The observation of eggs females laid, and assessment of expression levels of egg hatchability-related genes in N. lugens female adults
After the anesthetizing effect of dsRNAs-injected females vanished, the survived females were transferred into glass tubes (2.5 cm diameter, 15 cm height) containing 2–3 tillering rice stems and paired with untreated males at 1:2 ratio (female to male). The rice stems were replaced every 24 h during the pre-oviposition periods. Embryonic development of eggs can be accurately assessed using morphological landmarks. Previous studies have demonstrated that eyespot formation and yolk distribution patterns are reliable indicators of embryonic developmental progress and can be used to evaluate developmental status or delays in insect eggs (Panfilio, 2008; Fan et al., 2020). In this study, after the females laid eggs, rice stems containing eggs laid by the N. lugens females were collected at 2, 5, and 8 days after oviposition. The eggs laid by females in the rice stems were dissected, and the morphological characteristics of the egg development were observed through a microscope (OLYMPUS CX23 Japan), and photographed and recorded with a DS-Fi2 digital camera (Nikon Tokyo Japan). The newly emerged females were injected with dsRNAs as described in the previous sections, and the surviving female adults were collected at 2 days and 4 days post-emergence qRT-PCR and the procedure as mentioned above were used to determine the expression levels of egg hatchability-related genes (Cpr3, Cpr8, Cpr10, Cpr24, Cpr36, Cpr47, Cpr51, Cpr52, Cpr54, Cpr58, Cpr73, Cpr90, Cpr94, TwdIE3, CPAP1-E, CPAP1-H, CPAP1-I, CPAP3-B, CPAP3-D1, HNF4, Hox3, and Let1) (Pan et al., 2018; Ren et al., 2018; Cheng et al., 2020; Lu et al., 2023). The treatment and control consisted of three independent biological replicates, and each replicate was composed of 10 individuals.
2.7 Data analysis
Student's t-tests were used to compare statistical differences in the gene expression levels and biological parameters of N. lugens between the control and gene-silenced groups. One-way analysis of variance (ANOVA) by the Tukey test was used to compare statistical differences in the gene expression levels of CYP303A1 in different tissues. All statistical tests were conducted in SPSS 22.0 (IBM Inc., Armonk, NY, United States), and plots were generated using Origin 2023 (OriginLab Inc., Northampton, United Kingdom).
3 Results
3.1 The tissue-specific expression profile and RNAi efficiency of CYP303A1 in N. lugens females
To investigate the physiological function of CYP303A1 in N. lugens females, we analysed the expression profiles of CYP303A1 in female different tissues by RT-qPCR. The results showed that the expression level of CYP303A1 was significantly different in various tissues (F = 52.2, P < 0.001), and was predominantly enriched in female fatbodies (Fb) and leg, and showed a lower transcript level in cuticle (Figure 1A).
*The tissue-specific expression level (A) and silencing efficiency of CYP303A1 (B) in females. All data are means ± SE. The different lowercase letters represent statistically significant differences in the gene expression levels of CYP303A1 in different tissues by the Tukey test. The asterisks (*P < 0.05, *P < 0.01) represent statistically significant differences between control and treatment by Student’s t-test.
We further determined the RNAi efficiency of CYP303A1 in females. Compared with the dsGFP-injected females, the gene expression levels of newly emerged females injected with 100 ng, 150 ng and 200 ng of dsCYP303A1 decreased by 38.8% (t = 4.3, P < 0.05), 49.3% (t = 5.0, P < 0.01) and 66.6% (t = 5.3, P < 0.01), respectively, at 2 days. Based on the silencing efficiency, 200 ng of dsCYP303A1 was selected for subsequent experiments (Figure 1B).
3.2 Effect of silencing CYP303A1 on ovarian development in female adults
We further analyzed the effect of silencing CYP303A1 on the ovarian development of N. lugens females. The results showed that the mean ovarian area (Figures 2A DAE: t = 0.6, P > 0.05; 4 DAE: t = 1.4, P > 0.05) as well as the number of mature eggs (Figures 2B DAE: t = 0.4, P > 0.05; 4 DAE: t = 1.0, P > 0.05) were reduced in CYP303A1-suppressed females than dsGFP-injected females at 2, and 4 days post-emergence, but there were no statistical differences. Moreover, the morphological observations showed that the ovaries of both dsGFP-injected and dsCYP303A1-injected females were completely filled with regular banana-shaped oocytes, which were closely arranged in the ovarioles at 2 and 4 DAE (Figures 2C–F). These results indicated that suppression of CYP303A1 might be involved in the cuticle barrier construction. These results indicated that suppression of CYP303A1 would not disrupt ovarian development in N. lugens females.
Effects of dsCYP303A1 on ovarian area (A), number of mature eggs (B), and ovary development (C–F) of N. lugens females. The ns represent no significant differences in expression level between control (dsGFP) and treatment (dsCYP303A1) by Student's t-test.
3.3 Silencing of CYP303A1 decreased the number of offspring in N. lugens
Subsequently, we statistically examined the reproductive and population parameters of N. lugens after CYP303A1 silencing in females. The results showed that reproductive parameters, including number of eggs laid (Table 1, t = 0.34, P > 0.05), pre-oviposition (t = 0.27, P > 0.05), oviposition periods (t = 1.47, P > 0.05), and female longevity (t = 0.44, P > 0.05) were not significantly different between dsCYP303A1-injected N. lugens and controls (Table 1). However, silencing CYP303A1 significantly affected the population parameters of N. lugens. Inhibition of CYP303A1 significantly prolonged the egg durations by 47.3% and significantly reduced egg hatchability by 68.6% compared to the controls (Table 1). Ultimately, the population parameters of dsCYP303A-injected N. lugens were significantly reduced by 68.1% compared to the control (Table 1). These results demonstrate that inhibition of CYP303A1 will not affect female ovary development, but will reduce the number of offspring by decreasing egg hatchability of N. lugens.
3.4 Silencing of CYP303A1 delayed the egg development of N. lugens
The silencing of CYP303A1 significantly delayed the egg development of N. lugens. The eggs laid by dsCYP303A1- and dsGFP-treated females showed no obvious phenotypic differences at 2 days after oviposion (Figures 3A,D). Compared with the dsGFP-treated females, some eggs did not develop eyespots after 5 days of egg laying in dsCYP303A1-treated newly emerged females (Figures 3B,E). In dsGFP-treated newly emerged females, embryonic eye spots developed normally after 8 days of egg laying, and a distinct yellow substance was produced and accumulated at the egg tip (Figure 3C). In contrast, in dsCYP303A1-treated newly emerged females, embryonic eye spot development was delayed after 8 days of egg laying, and the egg tip exhibited a transparent substance (Figure 3F).
Effects of silencing CYP303A1 on embryonic development in N. lugens. Note: (A): Eggs laid for 2 days by dsGFP-treated females; (B): Eggs laid for 5 days by dsGFP-treated females; (C): Eggs laid for 2 days by dsGFP-treated females; (D): Eggs laid for 2 days by dsCYP303A1-treated females; (E): Eggs laid for 5 days by dsCYP303A1-treated females; (F): Eggs laid for 8 days by dsCYP303A1-treated females. Red circle in the figure mark embryo eyespots, yellow circle in the figure mark aggregated substance in the eggs.
3.5 Suppression of CYP303A1 reduced the transcript level of hatchability-related genes in N. lugens female adults
Finally, we investigated the effects of CYP303A1 knockdown on the expression of egg-hatching-related genes in female N. lugens. These genes included Cpr3, Cpr8, Cpr10, Cpr24, Cpr36, Cpr47, Cpr51, Cpr52, Cpr54, Cpr58, Cpr73, Cpr90, Cpr94, TwdIE3, CPAP1-E, CPAP1-H, CPAP1-I, CPAP3-B, CPAP3-D1, as well as transcription factors HNF4, Hox3, and Let1. The results showed that injection of dsCYP303A1 significantly downregulated the expression level of Cpr52 by 49.0% (Figure 4A, t = 4.2, P < 0.05) and upregulated TwdIE3 expression by 32.0% (Figure 4A, t = 3.1, P < 0.05) in females at 2 days post-eclosion. At 4 days post-eclosion, the expression levels of Cpr52, HNF4, and Let1 were reduced by 75.2% (Figure 4B, t = 4.1, P < 0.05), 22.8% (Figure 4B, t = 3.03, P < 0.05), and 32.0% (Figure 4B, t = 2.87, P < 0.05), respectively, in dsCYP303A1-treated females compared to the dsGFP controls. These findings suggest that silencing CYP303A1 markedly suppresses the expression of genes associated with egg hatching, thereby impairing embryonic development and hatching rate.
*Effects of silencing CYP303A1 on the expression levels of genes involved in embryonic development in N. lugens. Note: (A) The expression levels of hatchability-related genes in females at 2 days post-eclosion. (B) The expression levels of hatchability-related genes in females at 4 days post-eclosion. All data are means ± SE. The asterisks (P < 0.05) represent statistically significant differences between control and treatment by Student’s t-test.
4 Discussion
In oviparous insects, reproduction is a complex process involving multiple sequential stages, including previtellogenesis, vitellogenesis, chorion formation, oocyte maturation, and egg hatching (Cheng et al., 2020). To date, most cytochrome P450 (CYP) genes known to regulate insect reproduction are members of the Halloween gene family, which are essential for the biosynthesis of 20E (Lao et al., 2015; Truman, 2019; Hafeez et al., 2022). Functional studies have shown that silencing CYP307A2, CYP302A1, or CYP314A1 in N. lugens results in defective ovarian and egg development and markedly reduced fecundity (Zhou et al., 2020). Similarly, mutations in CYP306A1 or CYP302A1 in Drosophila melanogaster cause embryonic lethality (Chávez et al., 2000; Niwa and Niwa, 2016). In Schistocerca gregaria, RNAi knockdown of CYP307A1, CYP306A1, or CYP314A1 disrupts oocyte development and leads to reduced egg production and hatching success (Schellens et al., 2022). Ecdysteroids are aslso considered as the classic insect hormones involved in oogenesis of insects; however, their function and importance differ between distinct insect orders (Song and Zhou, 2020; Khalid et al., 2021). Ecdysteroids are needed to establish and maintain the stem cell niche (Gancz et al., 2011), to stimulate follicle cell formation and border cell migration (Morris and Spradling, 2012), to coordinate the onset of vitellogenesis with the availability of nutrients, to stimulate yolk polypeptide synthesis in the fat body, and finally, to induce choriogenesis (Buszczak et al., 1999; Carney and Bender, 2000; Terashima and Bownes, 2004). In previous study, we demonstrated that CYP303A1 plays a critical role in molting and metamorphosis of N. lugens. CYP303A1 was highly expressed during the pre-molt stage and primarily localized in tissues associated with cuticle formation. RNAi-mediated silencing of CYP303A1 significantly reduced the titer of 20E and downregulated key genes in the 20E signaling pathway, leading to impaired transcription of cuticular proteins and disrupted molting and metamorphic processes. These results indicate that CYP303A1 exerts a central role in molting and metamorphosis by regulating 20E signaling. Notably, the functions of CYP303A1 in other physiological processes of N. lugens have not been reported. Given the critical role of 20E in female reproduction and ovary development, CYP303A1 is likely to influence reproductive capacity and ovarian development through modulation of 20E biosynthesis.
N. lugens is a highly fecund r-strategy insect, represents a persistent threat to rice production and agroecosystem sustainability (Hu et al., 2017; Gou et al., 2024). To date, 67 CYP450 genes have been identified in N. lugens (Xue et al., 2014). Among these, conserved CYP genes from the CYP2 family and mitochondrial CYP clan have been implicated in reproductive regulation (Li et al., 2015; Zhou et al., 2020). Previous studies have demonstrated that Halloween genes play key roles in regulating Vg and VgR expression, oocyte development, and embryogenesis (Peng et al., 2019; Wu Z. et al., 2020; Benrabaa et al., 2022; Jin et al., 2023). Intriguingly, recent findings suggest that CYP genes outside the Halloween family may also contribute to insect reproduction. For instance, CYP303A1 has been implicated in embryonic development in D. melanogaster, where its mutation leads to impaired dorsal vessel formation and late-stage embryonic arrest, resulting in lethality (Wu et al., 2019). However, whether CYP303A1 is involved in embryogenesis and reproductive regulation in hemipteran pests such as N. lugens remains unclear. In the current study, CYP303A1 is abundantly expressed in the fat body of adult females and exhibits moderate expression in the ovary, suggesting a potential role in ovarian or embryonic development. Interestingly, although CYP303A1 was highly expressed in the fat body, RNAi knockdown of CYP303A1 did not significantly affect the transcript levels of Vg or VgR, nor did it impact ovarian morphology, the number of mature oocytes, ovarian size, oviposition parameters, female longevity, or progeny sex ratio. This discrepancy suggests that CYP303A1 may execute tissue-specific roles distinct from yolk protein synthesis. In the fat body, CYP303A1 might participate in processes such as ecdysteroid biosynthesis, lipid mobilization, or nutrient homeostasis, thereby indirectly contributing to reproductive fitness. By contrast, in embryos, its role appears to be directly associated with embryogenesis and successful hatchability through the regulation of developmental gene networks, and previous studies in N. lugens have shown that CYP303A1 is highly expressed in eggs (Wu et al., 2024). In D. melanogaster, CYP303A1 was predominantly enriched in the ring gland, and was essential in embryonic development (Wu et al., 2019). It also indicated the functional diversification of P450 genes.
Moreover, we further found that silencing CYP303A1 significantly prolonged the egg developmental period and markedly reduced egg hatchability. The embryonic development of N. lugens eggs exhibits clear stage-specific characteristics and is closely temperature-dependent (Fan et al., 2020). At room temperature, the egg period lasts ∼192 h from oviposition to first-instar nymph hatching. In early embryogenesis (0–6 h after egg laying, AEL), eggs appear oyster white and gradually turn yellow, with synchronous nuclear divisions forming a syncytial stage, providing an optimal window for RNAi or genome-editing (Huang et al., 2016). By 30 h AEL, the germ band forms, the embryo elongates and segments (intermediate germ development). Eyespots appear at 96 h AEL, darken at 120 h, abdominal segments form sequentially, appendages complete by 168 h, and nymphs hatch at 192 h AEL (Fan et al., 2020). Egg length and width increase throughout development, reflecting rapid cellular proliferation and morphological remodeling. In the current study, the eggs from dsCYP303A1-treated females exhibited delayed eyespot development and transparent terminal yolk structures that failed to aggregate and appear yellow, strongly suggesting that CYP303A1 plays a critical role in embryonic development in N. lugens. In Locusta migratoria, CYP303A1 is mainly expressed at fourth- and fifth-day of the egg stage (Zhang et al., 2018). Interestingly, although CYP303A1 is conserved across insects, its knockdown produces divergent phenotypes between different species. In N. lugens, eggs are laid externally on plant tissues, and early embryogenesis requires the formation of a serosal cuticle to protect the embryo and support nutrient allocation (Fan et al., 2020). Consequently, CYP303A1 may primarily influence embryonic morphogenesis. In contrast, in D. melanogaster, the developmental arrest occurred in the late embryonic development in the CYP303A1 mutants, showing an abnormal dorsal vessel (Wu et al., 2019). In holometabolous insects such as Drosophila, embryogenesis occurs within a more protected eggshell environment (Donoughe, 2022), and CYP303A1 may play a greater role in dorsal vessel development or hormonal regulation. These observations suggest a life-history–dependent functional diversification of CYP303A1. Compared to the known ecdysteroidogenic genes exhibiting a typical Halloween-class embryonic phenotype, the CYP303A1 mutants have a later stage of arrest in embryonic development after near completion of dorsal closure (Wu et al., 2019). In a previous study, we demonstrated that the knockdown of CYP303A1 significantly downregulated thetranscript levels of ecdysteroid biosynthesis-related genes, CYP307A1 and CYP314A1, as well as reducing the 20E titers in N. lugens (Wu et al., 2024; Du et al., 2025). 20E is a key insect steroid hormone that plays vital roles in female reproduction, particularly in regulating oogenesis and embryogenesis. During oogenesis, 20E promotes vitellogenin synthesis in the fat body and facilitates its uptake into oocytes via vitellogenin receptors, thereby supporting yolk accumulation and oocyte maturation. In embryogenesis, maternally deposited 20E participates in coordinating crucial developmental events such as germband extension and cuticle formation (Swevers, 2019; Song and Zhou, 2020; Khalid et al., 2021; Schellens et al., 2022). In S. gregaria, depleting the expression of SchgrSpo (CYP307A1), SchgrSad (CYP315A1) and SchgrShd (CYP314A1) had a significant impact on oocyte development, oviposition and hatching of the eggs. Moreover, the shape of the growing oocytes, as well as the deposited eggs, was very drastically altered by the experimental treatments (Schellens et al., 2022). In Diaphorina citri, inhibition of Halloween gene expression in adults impeded the growth of the female ovary, diminished yolk formation, lowered vitellogenin transcription levels, and impaired female fecundity (Zhang et al., 2023). In Bombyx mori, knockdown of a dephosphorylation enzyme of 20E delayed development at early embryogenesis, whereas knockdown of an ecdysteroidogenic enzyme delayed development at early-middle embryogenesis (Fujinaga et al., 2020). These results demonstrated the essential role of 20E in insect embryonic development and further suggest that CYP303A1 might disturb the embryonic development of N. lugens by modulating the 20E signaling pathway.
Furthermore, we found that the expression of several genes associated with eggshell formation and embryonic development (Cpr52, TwdIE3, HNF4, and Let1) was significantly altered following CYP303A1 knockdown. These findings suggest that CYP303A1 plays a crucial role in coordinating late-stage embryogenesis, likely by regulating hatching-related gene expression. Cuticular proteins (CPRs) are the major structural components of the insect cuticle and play indispensable roles in cuticle formation, mechanical support, and protection (Pan et al., 2018; Mallick and Eleftherianos, 2024). During embryogenesis, the expression of specific CPR genes is tightly regulated both spatially and temporally to coordinate the formation of embryonic cuticular structures such as the serosa cuticle, embryonic epidermis, and chorion (Pan et al., 2018). In the late stages of embryogenesis, CPRs contribute to key morphogenetic events, including dorsal closure, head involution, and the formation of body segmentation (Charles, 2010). Disruption of cuticular protein genes can impair these processes, leading to defects in embryo elongation, desiccation resistance, and ultimately hatching failure (Moussian et al., 2006). Many holo- and hemimetabolous insects enhance their eggshells during embryogenesis by forming a serosal cuticle. Previous work in N. lugens, five cuticle protein coding genes, Cpr1/2/3/8/90, were specifically or highly expressed during the serosal cuticle formation period. TEM observations of the SC following parental RNAi against NlugCpr1/2/3/8/90 demonstrated that NlugCpr3/8/90 were essential for serosal cuticle formation (Lu et al., 2022). The transcription factors, HNF4, and Let1 have been reported that were essential for embryonic development (Ren et al., 2018; Cheng et al., 2020). In N. lugens, HNF4 was highly expressed in the fat body and ovary of females, and knockdown of HNF4 resulted in a dramatic reduction in egg hatching rate (Cheng et al., 2020). The Let1 accumulates during the serosal cuticle formation period in N. lugens, and is located in the serosal endocuticle. RNAi-mediated silencing of Let1 disrupted the serosal cuticle structure, accompanied by a loss of the outward barrier and 100% embryo mortality (Lu et al., 2023). These results suggest that silencing CYP303A1 altered the expression of several genes related to chorion formation and embryonic morphogenesis, further affecting embryonic development.
In conclusion, this study provides new insights into the role of CYP303A1, a non-Halloween cytochrome P450 gene, in the embryonic development of the N. lugens, and expands our understanding of P450 gene family diversity in insect reproductive biology beyond the well-studied Halloween genes. Although CYP303A1 is not involved in vitellogenesis or ovarian maturation, its high expression in eggs and fat body, along with RNAi-based functional analysis, indicates its essential role during embryogenesis. Knockdown of CYP303A1 significantly reduced hatchability and delayed embryonic development, accompanied by abnormal eyespot formation and yolk distribution. These findings indicate that CYP303A1 is indispensable for proper embryogenesis, likely through the regulation of hatching-related genes, and highlight its potential as a molecular target for RNAi-based pest control. However, the translation of such molecular targets into field-deployable RNA pesticides is constrained by delivery challenges. Naked dsRNA is inherently unstable under field conditions, rapidly degrading by UV radiation, rainfall, and nucleases, which greatly limits its persistence. Moreover, phloem-feeding insects such as planthoppers exhibit low uptake efficiency of exogenous RNA, making it difficult to reproduce laboratory efficacy in the field. Despite these hurdles, significant opportunities are emerging: spray-induced gene silencing (SIGS) provides a practical, non-transgenic approach for large-scale deployment, while nanocarrier-based systems can markedly improve dsRNA stability, cellular uptake, and trans-barrier transport (Hoang et al., 2022; Qiao et al., 2024). Integrating these delivery innovations with key developmental targets such as CYP303A1 may enable the transition of RNAi from laboratory proof-of-concept to effective field application, offering a sustainable strategy for N. lugens management.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Balabanidou V.Kampouraki A.Maclean M.Blomquist G. J.Tittiger C.Juárez M. P. (2016). Cytochrome P 450 associated with insecticide resistance catalyzes cuticular hydrocarbon production in Anopheles gambiae . P Natl. Acad. Sci. 113, 9268–9273. 10.1073/pnas.1608295113 27439866 PMC 4995928 · doi ↗ · pubmed ↗
- 2Benrabaa S. A.Orchard I.Lange A. B. (2022). The role of ecdysteroid in the regulation of ovarian growth and oocyte maturation in rhodnius prolixus, a vector of chagas disease. J. Exp. Biol. 225, jeb 244830. 10.1242/jeb.244830 36268612 · doi ↗ · pubmed ↗
- 3Buszczak M.Freeman M. R.Carlson J. R.Bender M.Cooley L.Segraves W. A. (1999). Ecdysone response genes govern egg chamber development during mid-oogenesis in Drosophila. Development 126, 4581–4589. 10.1242/dev.126.20.4581 10498692 · doi ↗ · pubmed ↗
- 4Carney G. E.Bender M. (2000). The Drosophila ecdysone receptor (Ec R) gene is required maternally for normal oogenesis. Genetics 154, 1203–1211. 10.1093/genetics/154.3.1203 10757764 PMC 1461007 · doi ↗ · pubmed ↗
- 5Charles J.-P. (2010). The regulation of expression of insect cuticle protein genes. Insect Biochem. Mol. Biol. 40, 205–213. 10.1016/j.ibmb.2009.12.005 20060042 · doi ↗ · pubmed ↗
- 6Chávez V. M.Marqués G.Delbecque J. P.Kobayashi K.Hollingsworth M.Burr J. (2000). The Drosophila disembodied gene controls late embryonic morphogenesis and codes for a cytochrome P 450 enzyme that regulates embryonic ecdysone levels. Development 127, 4115–4126. 10.1242/dev.127.19.4115 10976044 · doi ↗ · pubmed ↗
- 7Cheng Y.Li Y.Li W.Song Y.Zeng R.Lu K. (2020). Effect of hepatocyte nuclear factor 4 on the fecundity of Nilaparvata lugens: insights from RNA interference combined with transcriptomic analysis. Genomics 112, 4585–4594. 10.1016/j.ygeno.2020.08.002 32763353 · doi ↗ · pubmed ↗
- 8Dermauw W.Van Leeuwen T.Feyereisen R. (2020). Diversity and evolution of the P 450 family in arthropods. Insect Biochem. Mol. Biol. 127, 103490. 10.1016/j.ibmb.2020.103490 33169702 · doi ↗ · pubmed ↗