Ophiostoma babimostense and Sporothrix europaea (Ascomycota, Ophiostomatales), two new ophiostomatalean species, associated with ambrosia and bark beetles in Norway and Poland

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
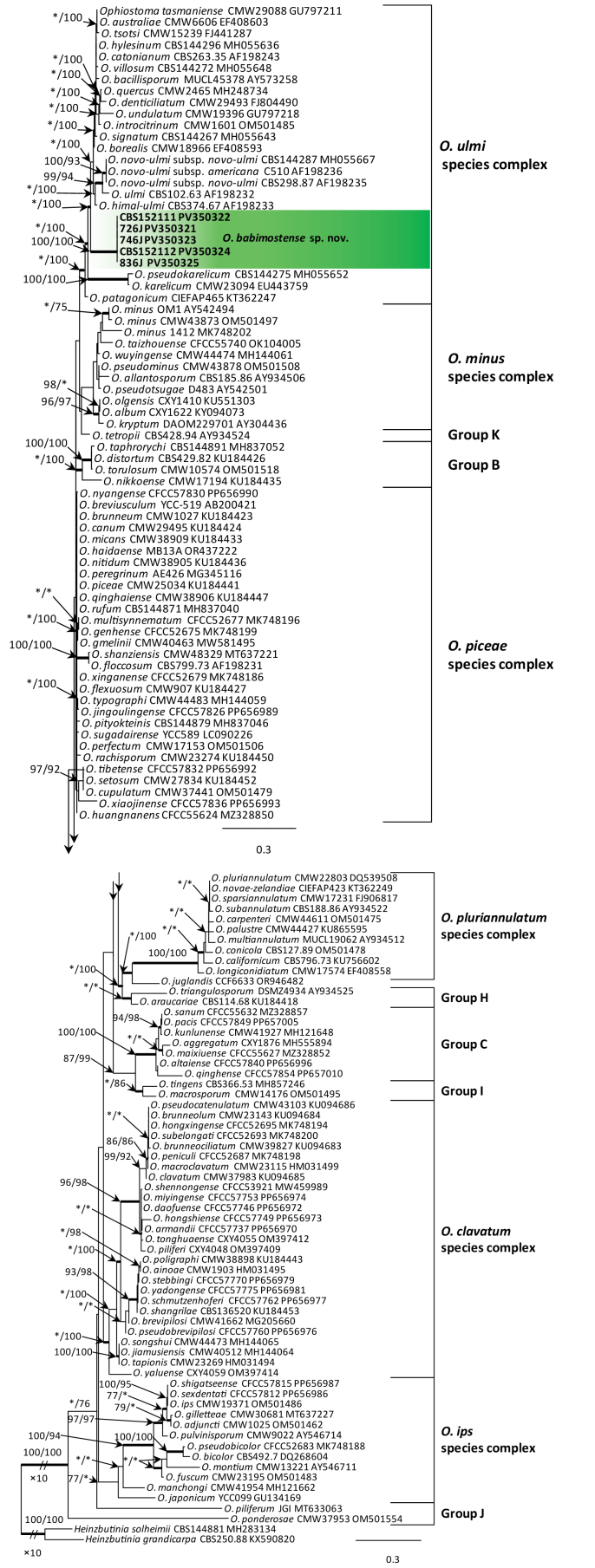 Figure 1
Figure 1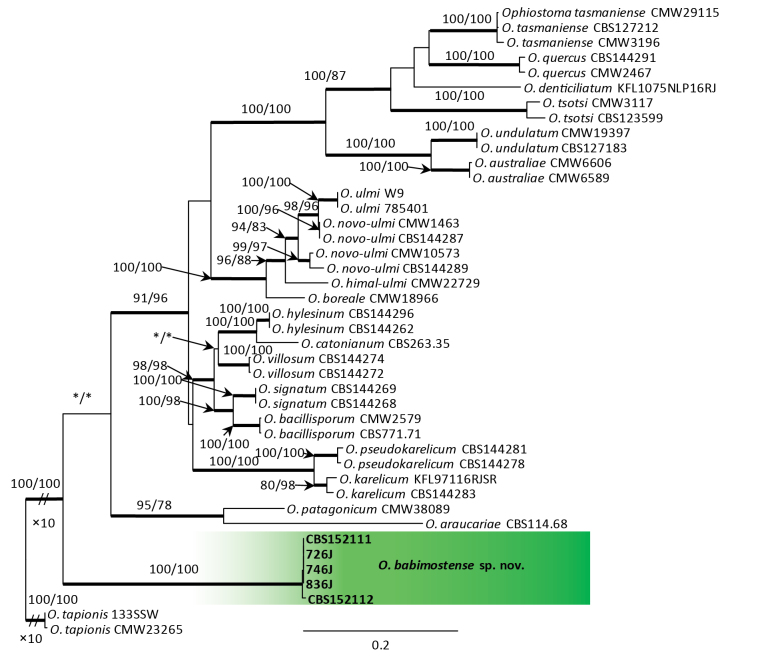 Figure 2
Figure 2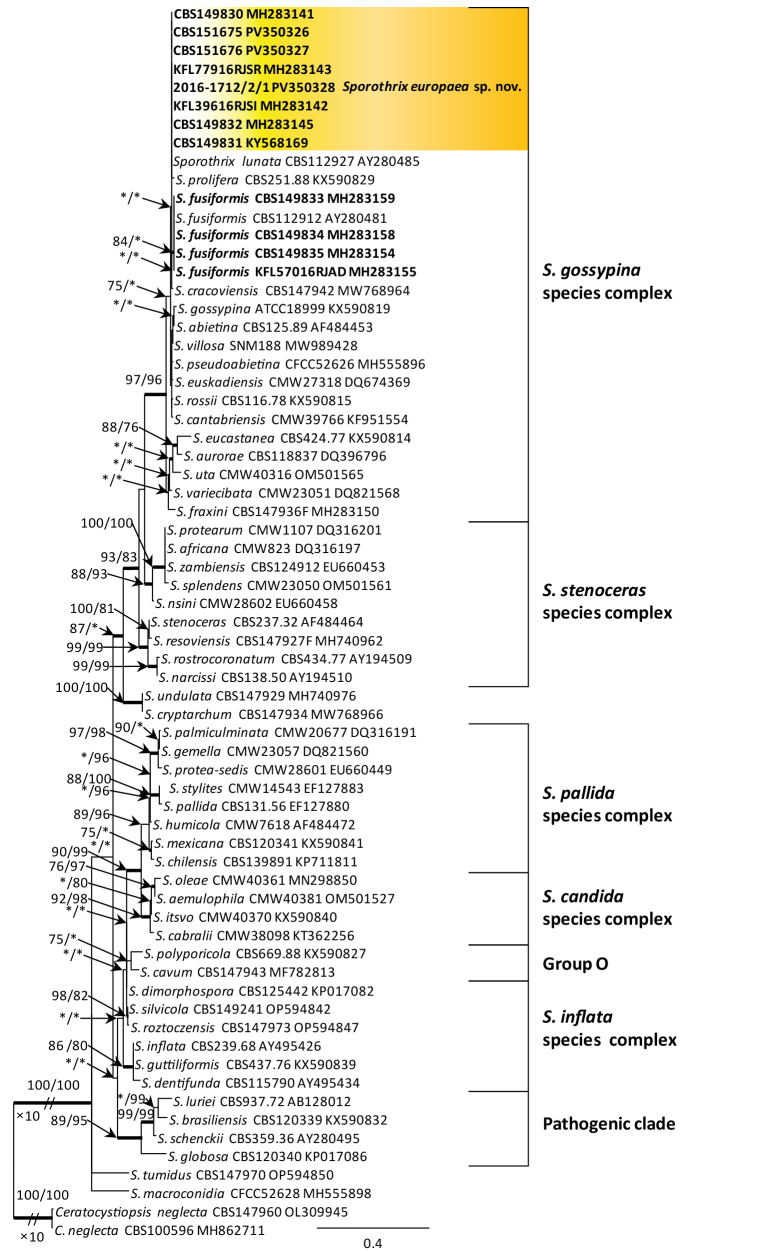 Figure 3
Figure 3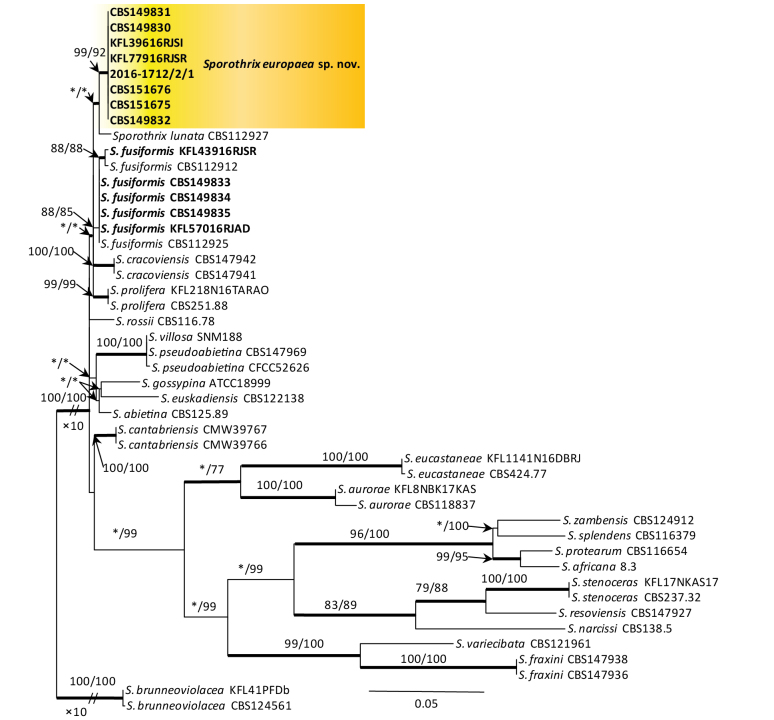 Figure 4
Figure 4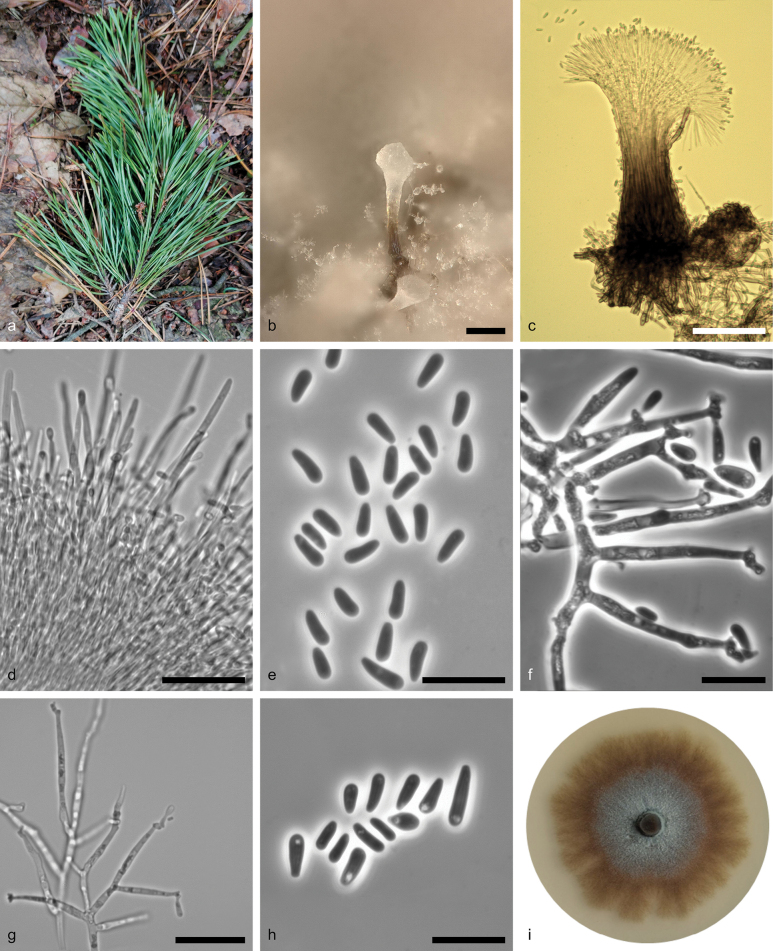 Figure 5
Figure 5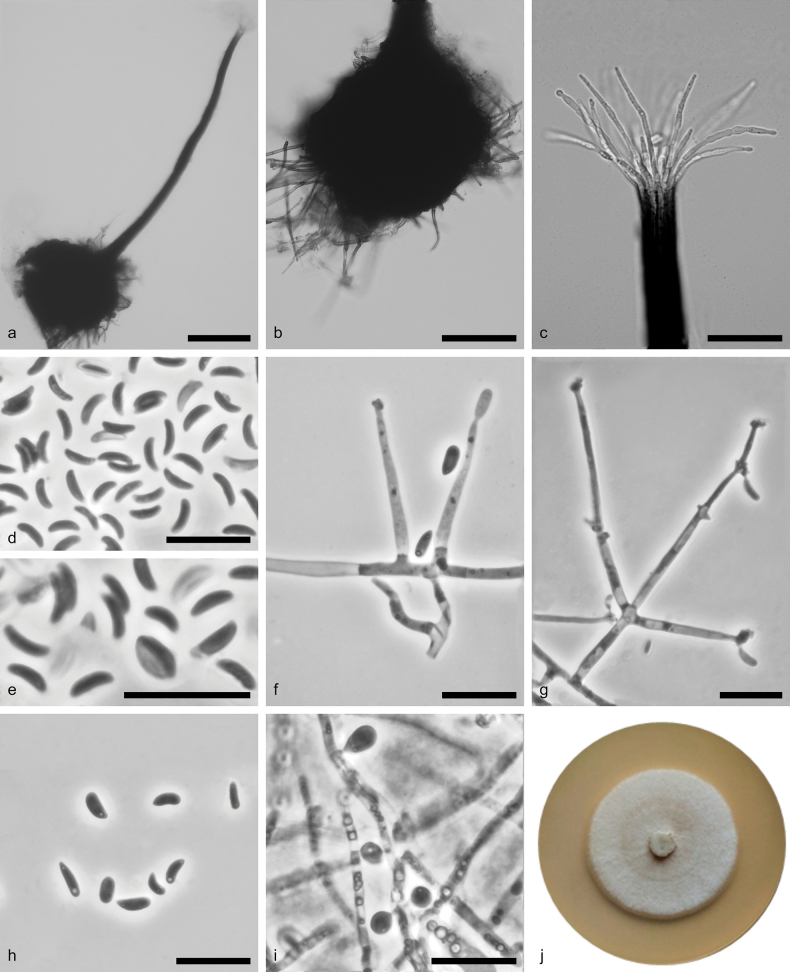 Figure 6
Figure 6| Taxa | Previously labelled | Isolate noA. | Source | Site | GenBank accessionsB | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
| KFL/NIBIO | Host/vector |
|
|
|
|
| |||
| KFL726J | Babimost, Poland |
|
|
|
|
| |||||
| KFL728J | Babimost, Poland |
|
|
|
|
| |||||
| KFL746J | Babimost, Poland |
|
|
|
|
| |||||
| KFL828J | Babimost, Poland |
|
|
|
|
| |||||
| KFL836J | Babimost, Poland |
|
|
|
|
| |||||
| NIBIO2016-1656/3/1 | Ås, Norway |
|
|
|
|
| |||||
| NIBIO2016-1657/2/1 | Ås, Norway |
|
|
|
|
| |||||
| NIBIO2016-1712/2/1 | Ås, Norway |
|
|
|
|
| |||||
| KFL77916RJSRD | Długołęka-Poland |
|
|
|
|
| |||||
| KFL39616RJSID | Wierzchosławice, Poland |
|
|
|
|
| |||||
| KFL12WRJXMD | Prószków, Poland |
|
|
|
|
| |||||
| KFL121LMDD | Rudziniec, Poland |
|
|
|
|
| |||||
| KFL104716RJXSD | Resko, Poland |
|
|
|
|
| |||||
|
| CBS112912T |
| Azerbaijan |
|
|
|
|
| |||
| KFL104416RJADD | Resko, Poland |
|
|
|
|
| |||||
| KFL103616RJADD | Resko, Poland |
|
|
|
|
| |||||
| KFL56916RJADD | Resko, Poland |
|
|
|
|
| |||||
| KFL57016RJADD | Resko, Poland |
|
|
|
|
| |||||
|
|
| Austria |
|
|
|
|
| ||||
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsForest Insect Ecology and Management · Plant Pathogens and Fungal Diseases · Yeasts and Rust Fungi Studies
Introduction
Ophiostomatalean fungi (Ophiostomatales, Ascomycota) are characterised by the formation of flask-shaped perithecia and various asexual morphs, including mononematous or synnematous forms (Wingfield et al. 1993; De Beer and Wingfield 2013). These fungi include tree- or wood-infecting species causing a dark bluish discoloration in the sapwood or serious tree diseases and some of them are also the causal agents of human diseases. Ophiostomatalean species are well known to be closely associated with bark- and wood-dwelling beetles and mites (De Beer et al. 2022).
In Europe, surveys for ophiostomatalean fungi from different habitats are constantly improving. Combined with multi-locus molecular phylogenetic analysis has led to the identification of many species of ophiostomatalean fungi, including dozens of novel species (e.g. Linnakoski et al. (2016); Jankowiak et al. (2017, 2019b)). In particular, extensive surveys have been conducted on the diversity and taxonomy of ophiostomatalean and microascalean fungi in Central and Northern Europe. For example, surveys of insect-associated mycobiomes in Norway and Poland, yielded descriptions of almost 30 new species of Ceratocystiopsis (Jankowiak et al. 2022), Graphilbum (Jankowiak et al. 2020), Graphium (Jankowiak et al. 2023a), Ophiostoma (Aas et al. 2018; Jankowiak et al. 2019a) and Sporothrix (Ostafińska et al. 2021). More recent discoveries generated descriptions of a new genus Hausneria (Crous et al. 2024) from a gallery of Dryocoetes alni in Norway and the new species, Ophiostoma juglandis from Dryocoetes himalayensis on Juglans regia in Czechia (Májek et al. 2025). In Poland, the screening of ophiostomatalean fungi was conducted from tree wounds (Jankowiak et al. 2019c), in forest soils (Bilański et al. 2023) and in bird’s nests (Błońska et al. 2021), yielding two new soil-inhabiting Sporothrix species (Bilański et al. 2023), one new species of Hawksworthiomyces from nests of Ciconia ciconia (Crous et al. 2023) and three new species of Leptographium from tree wounds (Jankowiak et al. 2018a). These surveys revealed a plethora of new beetle-fungus associations and led to the discovery and description of many new species, showing there is still a lot of unknown taxonomic and ecological diversity to be uncovered for the ophiostomatalean fungi in Europe.
Ophiostoma is one of the largest genera within the Ophiostomatales and includes tree- or wood-infecting fungi. Currently, more than 170 species are recognised worldwide (De Beer et al. 2022; Bhunjun et al. 2024; Feau et al. 2024; Májek et al. 2025; Wang et al. 2024). Members of Ophiostoma are characterised by pigmented ascomata with slender necks and allantoid, reniform, cylindrical to ossiform in side view ascospores. In addition, Hyalorhinocladiella-, Leptographium-, Pesotum- or Sporothrix-like asexual morphs are produced in culture and nature (De Beer et al. 2022). Ophiostoma is mostly structured into six phylogenetically well-supported lineages or so-called species complex. They are the O. clavatum–, O. ips–, O. minus–, O. piceae–, O. pluriannulatum– and O. ulmi lineages and species complexes. In addition, some species do not form part of these lineages, such as O. piliferum and O. tetropii (De Beer et al. 2022; Wang et al. 2024). Ophiostoma can be found in a wide range of habitats, but most species are commonly associated with forest trees on which they form more or less stable associations together with bark- and wood-dwelling beetles, mites (Six 2012) and, in a few cases, nematodes (Bhunjun et al. 2024). Some members of Ophiostoma, such as O. piliferum and O. minus, cause the economically important blue-stain in freshly exposed sapwood of softwood species (e.g. Seifert (1993); Uzunović and Byrne (2013); Jankowiak et al. (2021)) or sawn timber (Jankowiak et al. 2018b). In addition, O. novo-ulmi is an example of a highly virulent pathogen that has been responsible for Dutch elm disease in Europe, western Asia and North America (Brasier 1991). Ophiostoma has a worldwide distribution, but it is especially abundant in the north temperate to boreal conifer ecosystems, in Asia, Europe and North America. They have been observed from Abies, Larix, Picea, Pinus, Pseudotsuga and Tsuga. Ophiostoma species also grow from hardwood, such as Quercus or Fagus (Jankowiak et al. 2019b, c). Some species, such as Ophiostoma ips, O. piceae, O. piliferum and O. quercus, are globally widespread, possibly due to human activity and the movement of wood products around the globe (Taerum et al. 2018; Bhunjun et al. 2024).
Sporothrix is also a large genus in the Ophiostomatales, with currently 70 recognised species arranged in five phylogenetically well-supported lineages, forming species complex, including the S. candida–, S. inflata–, S. stenoceras–, S. gossypina– and S. pallida species complexes (De Beer et al. 2016, 2022; Wang et al. 2019; Bilański et al. 2023). Other species form part of several unresolved smaller phylogenetic lineages (Groups D-G) or two lineages Sporothrix insertae sedis (XVI & XIX) defined by De Beer et al. (2022). The S. gossypina complex is considered as the largest at present, with 18 accepted species. Members of this complex are characterised by the presence of globose ascomatal base with black necks and allantoid to reniform in side view ascospores. The asexual morphs include simple, micronematous to mononematous conidiophores, with denticulate or not denticulate conidiogenous cells showing sympodial growth. Conidia hyaline to that vary in size and shape, mostly subglobose to oblong, obovoid, clavate to strongly curved, guttuliform to fusiform (De Beer et al. 2022). Similar to Ophiostoma, members of Sporothrix are widely distributed across various climatic zones of the world, colonising diverse environments (De Beer and Wingfield 2013; De Beer et al. 2016). The greatest numbers of species are found on the bark and wood of different forest trees and in the infructescences of Protea spp. (e.g. Roets et al. (2013); De Errasti et al. (2016); Bilański et al. (2023)). Other species have been described from soil, ambrosia and bark beetles, mites and from the fruiting bodies of basidiomycetes (e.g. Constantinescu and Ryman (1989); Marmolejo and Butin (1990); De Meyer et al. (2008)). Several species are also well-known as human and animal pathogens (e.g. Lòpez-Romero et al. (2011); Zhang et al. (2015)).
During a survey of ophiostomatalean fungi on hardwoods in Poland and Norway (Aas et al. 2018; Jankowiak et al. 2019b), isolates of an undescribed Sporothrix species with a sexual state resembling species in the S. gossypina species complex were isolated from different ambrosia and bark beetle species. In addition, isolates of an unknown fungal species with morphological attributes (synnemata) matching those described for species of the Ophiostoma were isolated from Scots pine (Pinus sylvestris L.) shoots infested by an unknown species of Tomicus. The purpose of this study is to characterise these fungi using the morphology, phylogenetic analyses and to formally describe them as novel species of Sporothrix and Ophiostoma.
Materials and methods
Isolates and herbarium specimens
A collection of ten Polish and three Norwegian isolates were used in this study. Isolates of Ophiostoma sp. were collected in pure Scots pine stands in western Poland (Babimost: 52°09′11″N, 15°50′33″E) in October 2023. Isolations were made from fallen shoots of Scots pine pruned by Tomicus spp. as described by Jankowiak and Kolařík (2011). Isolates of Sporothrix sp. were collected during surveys of hardwood-infesting bark and ambrosia beetles in Poland and Norway (Aas et al. 2018; Jankowiak et al. 2019b). The cultures are maintained in the culture collection of the Department of Forest Ecosystems Protection, University of Agriculture in Krakow, Poland and in the culture collection of Norwegian Institute of Bioeconomy. The ex-type isolates and representative isolates of the new species described were deposited in the culture collection (CBS) of the Westerdijk Fungal Biodiversity Institute, Utrecht, The Netherlands and in the culture collection (CMW) of the Forestry and Agricultural Biotechnology Institute (FABI), University of Pretoria, Pretoria, South Africa. Dried cultures were deposited as holotype specimens in the National Biodiversity Collection – Herbarium KRAM, the W. Szafer Institute of Botany Polish Academy of Science, Kraków, Poland. Two reference strains were also obtained from collections of CBS. These included a living culture of Sporothrix fusiformis (CBS 112912) and Sporothrix lunata (CBS 119444) (Table 1).
PCR, sequencing and phylogenetic analyses
DNA extraction, PCR and sequencing were performed as described by Jankowiak et al. (2019a). For DNA sequencing and phylogenetic analyses, five loci were amplified as follows: the internal transcribed spacer regions ITS1-5.8S-ITS2 (ITS), the partial 28S ribosomal large subunit (LSU), the partial beta-tubulin (TUB2), the partial calmodulin (CAL) and the partial translation elongation factor 1-alpha (TEF1). The primers used for polymerase chain reaction (PCR) and sequencing of the various gene regions were as follows: ITS1-F (Gardes and Bruns 1993) and ITS4 (White et al. 1990) for ITS, LR5 and LR0R (Vilgalys and Hester 1990) for LSU, Bt2a and Bt2b (Glass and Donaldson 1995) for TUB2, EF2F (Marincowitz et al. 2015) and EF2R (Jacobs et al. 2004) or F-728F (Carbone and Kohn 1999) and EF2 (O’Donnell et al. 1998) for TEF1 and CL1 and CL2a (O’Donnell et al. 2000) or CL3F and CL3R (De Beer et al. 2016) for CAL.
BLAST searches using the BLASTn algorithm were performed to retrieve similar sequences from GenBank (http://www.ncbi.nlm.nih.gov) and accession numbers for these sequences are presented in the corresponding phylogenetic trees (Figs 1–4). Datasets were curated using Molecular Evolutionary Genetic Analysis (MEGA) 6.06 (Tamura et al. 2013). The ITS datasets included all available sequences for reference species of Ophiostoma and Sporothrix that could be retrieved from GenBank to resolve the placement of the isolates within the mentioned genera.
Phylogram from Maximum Likelihood (ML) analysis of ITS data for Ophiostoma spp. Polish isolates used in this study are in bold. Polish isolates used in this study are in bold. Bootstrap values (if ≥ 75%) for ML and Maximum Parsimony (MP) analyses are presented at the nodes as follows: ML/MP. Bold branches indicate posterior probabilities values ≥ 0.95 obtained from Bayesian Inference (BI) analysis. * Bootstrap values < 75%. The tree is drawn to scale (see bar) with branch lengths measured in the number of substitutions per site. The species complexes were designated, based on the classification of De Beer et al. (2022) and Wang et al. (2024). Heinzbutinia solheimii and H. grandicarpa represent the outgroup.
Phylogram from Maximum Likelihood (ML) analysis of the combined datasets of TUB2+TEF1 for the Ophiostoma ulmi species complex. Polish isolates used in this study are in bold. Bootstrap values (if ≥ 75%) for ML and Maximum Parsimony (MP) analyses are presented at the nodes as follows: ML/MP. Bold branches indicate posterior probabilities values ≥ 0.95 obtained from Bayesian Inference (BI) analysis. * Bootstrap values < 75%. The tree is drawn to scale (see bar) with branch lengths measured in the number of substitutions per site. Ophiostoma tapionis represents the outgroup.
Phylogram from Maximum Likelihood (ML) analysis of ITS data for Sporothrix spp. Norwegian and Polish isolates used in this study are in bold. Bootstrap values (if ≥ 75%) for ML and Maximum Parsimony (MP) analyses are presented at the nodes as follows: ML/MP. Bold branches indicate posterior probabilities values ≥ 0.95 obtained from Bayesian Inference (BI) analysis. * Bootstrap values < 75%. The tree is drawn to scale (see bar) with branch lengths measured in the number of substitutions per site. The species complexes were designated, based on the classification of De Beer et al. (2016). Ceratocystiopsis neglecta represents the outgroup.
Phylogram from Maximum Likelihood (ML) analysis of the combined datasets of TUB2+CAL for the Sporothrix stenoceras and S. gossypina complexes. Norwegian and Polish isolates used in this study are in bold. Bootstrap values (if ≥ 75%) for ML and Maximum Parsimony (MP) analysis are presented at the nodes as follows: ML/MP. Bold branches indicate posterior probabilities values ≥ 0.95 obtained from Bayesian Inference (BI) analysis. * Bootstrap values < 75%. The tree is drawn to scale (see bar) with branch lengths measured in the number of substitutions per site. Sporothrix brunneoviolacea represents the outgroup.
Sequence alignments were performed using the online version of MAFFT 7 (Katoh and Standley 2013) using the E-INS-i strategy with a 200PAM/κ = 2 scoring matrix, a gap opening penalty of 1.53 and an offset value of 0.00. The alignments were checked manually with BioEdit 2.7.5 (Hall 1999). The resulting alignments and trees were deposited in TreeBASE (http://purl.org/phylo/treebase/phylows/study/TB2:S32067).
All phylogenetic analyses were performed independently for each gene regions (ITS, TUB2, TEF1 for Ophiostoma and ITS, CAL, TUB2 for Sporothrix). Sequences of the LSU (for Ophiostoma, Sporothrix), CAL (for Ophiostomа) and TEF1 (for Sporothrix) genes were generated for future reference. Gene regions showing no conflicts in the grouping of isolates at the terminal clades (i.e. TUB2 and TEF1 for Ophiostoma and TUB2 and CAL for Sporothrix), were combined and analysed as concatenated datasets.
Phylogenetic trees were inferred for each of the datasets using three different methods: Maximum Likelihood (ML), Maximum Parsimony (MP) and Bayesian Inference (BI). For ML and BI analyses, the best-fit substitution models for each dataset were determined using the corrected Akaike Information Criterion (AICc) in jModelTest 2.1.10 (Guindon and Gascuel 2003; Darriba et al. 2012). ML analyses were carried out using PhyML 3.0 (Guindon et al. 2010), utilising the Montpelier online server (http://www.atgc-montpellier.fr/phyml/). The ML analysis included bootstrap analysis (1000 bootstrap pseudoreplicates) in order to assess node support values.
MP analyses were performed using PAUP* 4.0b10 (Swofford 2003). Gaps were treated as the fifth state. Bootstrap analysis (1000 bootstrap replicates) was conducted to determine the levels of confidence for the nodes within the inferred tree topologies. Tree bisection and reconnection (TBR) was selected as the branch swapping option. The tree length (TL), consistency index (CI), retention index (RI), homoplasy index (HI) and rescaled consistency index (RC) were recorded for each analysed dataset after the trees were generated.
BI analyses using Markov Chain Monte Carlo (MCMC) methods were carried out using MrBayes 3.1.2 (Ronquist and Huelsenbeck 2003). Four MCMC chains were run for 10 million generations, applying the best-fit model for each dataset. Trees were sampled every 100 generations, resulting in 100,000 trees. Tracer 1.4.1 (Rambaut and Drummond 2007) was utilised to determine the burn-in value for each data-set. The remaining trees were utilised to generate a 50% majority rule consensus tree, which allowed for calculating posterior probability values for the nodes.
Morphology and growth rate
Morphological characters were examined for selected isolates as well as for the herbarium specimens selected as types. Cultures were grown on 2% malt extract agar (MEA) made up of 20 g malt extract, 20 g bacteriological lab-agar (Biomaxima S.A., Lublin, Poland) in 1 litre deionised water. Fungal cultures were derived from single conidia spores. The cultures were incubated at 25 °C and monitored regularly for the appearance of sporulating structures.
Attempts were made to induce the formation of sporulating structures, by placing autoclaved twigs of host trees at the centres of agar plates containing 2% MEA. To promote the production of ascomata, all isolates of each species were crossed in all possible combinations, following the technique described by Grobbelaar et al. (2009). These cultures were incubated 25 °C in darkness and monitored regularly for the appearance of sporulating structures.
Morphological features were examined by mounting fungal tissue in 80% lactic acid on glass slides and fruiting structures were observed using a Nikon Eclipse 50i microscope (Nikon, Tokyo, Japan) with an Invenio 5S digital camera (DeltaPix, Maalov, Denmark) to capture photographic images. Colour designations were based on the colour charts of Kornerup and Wanscher (1978).
For each taxonomically relevant structure, 50 measurements were made, when possible, using Coolview 1.6.0 (Precoptic, Warsaw, Poland). Averages, ranges and standard deviations were calculated for the measurements and these are presented in the format “(min–)(mean − SD)–(mean + SD)(–max)”.
Growth characters for the novel species were determined by analysing the growth for four isolates, including two for each species (Table 1). Agar disks (5 mm diam.) were cut from the actively growing margins of fungal colonies and these discs were placed at the centres of plates containing 2% MEA. Four replicate plates for each of the two putative new species were incubated at temperatures between 5 and 35 °C, at 5 °C intervals. The mean colony diameter and the radial growth (two measurements perpendicular to each other per plate) were determined 7 and 14 d after inoculation and growth rates were calculated as mm/d.
Results
DNA sequence data and phylogenetic analysis
DNA sequence data were generated for 13 isolates considered in this study (Table 1). BLAST analyses of the ribosomal DNA sequences and phylogenetic analyses of the ITS sequences placed five isolates within Ophiostoma and eight isolates within Sporothrix (Figs 1, 3).
The best evolutionary substitution models identified for the Ophiostoma datasets were as follows: GTR+I+G for ITS (−lnL = 7360.57), HKY+I+G for TUB2 (−lnL = 2541.70), GTR+I+G for TEF1 (−lnL = 5638.21) and GTR+I+G for the combined TUB2+TEF1 dataset (−lnL = 8370.00). For the Sporothrix datasets, the best substitution models were as follows: GTR+I+G for ITS (−lnL = 3664.91), GTR+G for TUB2 (−lnL = 1474.17), GTR+I+G for CAL (−lnL = 3780.98) and GTR+G for the combined TUB2+CAL dataset (−lnL = 5380.69).
Ophiostoma
Alignments for the ITS dataset contained 796 characters, while TUB2 and TEF1 had 396 and 731 characters, respectively. The combined TUB2+TEF1 dataset included 1127 characters. Of these, 297 characters in the ITS dataset, 150 in TUB2, 394 in TEF1 and 554 in the TUB2+TEF1 dataset were parsimony-informative. The aligned the TUB2 data used for phylogenetic analyses included exons 4, 5 and 6, interspersed by introns 3 and 4. The aligned the TEF1 data used for phylogenetic analyses included exons 4 and 5, preceded by intron 3.
The analyses of ITS sequences (Fig. 1) showed that five isolates grouped closest to the members of the O. ulmi species complex and were distinct from all other species in this species complex. These isolates formed distinct clade with high statistical supports. In the TUB2 and TEF1 trees (Suppl. material 1: figs S1, S2), these isolates formed well-supported lineages that clearly separated this newly-proposed species from all the other known species in the O. ulmi species complex. The combined analyses of the TUB2 and TEF1 datasets (Fig. 2) also showed that these isolates formed a distinct and well-supported clade within the O. ulmi complex.
Sporothrix
The ITS dataset consisted of 631 characters. For the individual gene regions, TUB2 had 364 characters and CAL had 825 characters. When TUB2 and CAL were combined (TUB2+CAL), the dataset contained 1189 characters. Regarding parsimony-informative characters, the breakdown was as follows: 210 in the ITS dataset, 87 in TUB2, 266 in CAL and 353 in the combined TUB2+CAL dataset. Alignments for the ITS dataset contained 631 characters and for TUB2+CAL 1189 characters, of which 210 and 353 were parsimony informative, respectively. The aligned the TUB2 data used for phylogenetic analyses included exons 4, 5 and 6, interspersed by introns 4 and 5. The aligned the CAL data used for phylogenetic analyses included exons 3, 4, 5 and 6, interspersed by introns 3, 4 and 5.
The analyses of ITS sequences showed that 13 isolates, labelled as Sporothrix sp. 4 and Sporothrix sp. 9 by Jankowiak et al. (2019b), resided in the S. gossypina species complex and were not distinct from some other species in this species complex (Fig. 3). Based on the TUB2 phylogeny, eight isolates of Sporothrix sp. 4 grouped closely with S. fusiformis. Based on the CAL phylogeny, isolates of Sporothrix sp. 4 formed a distinct and well-supported clade which was close to, but distinct from S. lunata (Suppl. material 1: figs S3, S4). The combined analyses of the TUB2 and CAL datasets showed that eight isolates of Sporothrix sp. 4 formed a well-resolved clade and this formed a sister clade with S. lunata (Fig. 4). Analyses of TUB2 and CAL sequences and the combined analyses of these both genes clearly showed that four isolates of Sporothrix sp. 9 belonging to known species, S. fusiformis (Fig. 4).
Taxonomy
Ophiostoma
babimostense
Taxon classificationFungiOphiostomatalesOphiostomataceae
R. Jankowiak sp. nov.
82804642-68E5-54AA-98C6-A5862372A6F8
858470
Etymology.
The specific epithet “babimostense” refers to the name of locality in Poland, Babimost, where the fungus was isolated for the first time.
Ophiostoma babimostense sp. nov. (CBS 152112). a. Fallen shoot of Scots pine pruned by Tomicus spp.; b, c. Synnemata in culture; d. Conidiogenous apparatus showing conidiogenous cells; e. Conidia; f, g. Mononematous, macronematous conidiophores with apical denticles; h. Conidia; i. Fourteen-day-old culture on MEA. Scale bars: 50 μm (b–c); 25 μm (d); 10 μm (e–f); 25 μm (g); 10 μm (h).
Type.
Poland • Lubuskie Province, Babimost, fallen shoots of Pinus sylvestris pruned by pine shoot beetles, Tomicus sp., October 2023, coll. R. Jankowiak (holotype KRAM F-60035, ex-type culture CBS 152112).
Description.
Sexual morph: not observed. Asexual morphs synnematous, pesotum-like and mononematous to micronematous (sporothrix-like type). Synnemata abundant on MEA and sterilised pine twigs, determinate, erect, single or in groups, arising from the agar, pine twigs or aerial mycelium and attached to substratum by brown, rhizoid-like hyphae, dark brown to black and becoming subhyaline to hyaline towards the conidiogenous apparatus, (127–)145.5–239(–315) μm long including the capitulum. Stipe dark brown to black at the bases, light brown or yellowish-brown at the centre and hyaline at the apex, (60–)86–153.5(–210) μm long, broadest towards the base, (16.5–)24–48.5(–81.5) µm down to (11.5–)18.5–44(–91) µm wide at the apex, cylindrical, smooth. Conidiophores branching divaricate or dichotomous with 2 (mostly) or 3 conidiogenous cells per branch point. Conidiogenous cells annellated, discrete, terminal, cylindrical, tapering towards apex, hyaline, smooth, (16.5–)27–42.8(–50) × (0.8–)0.9–1.5(–2) μm with. Conidia aseptate, hyaline, curved, obovate, (3–)3.5–4.5(–6) × (1–)1.5–2(–2.5) µm, accumulating in a terminal mucilagenous mass, hyaline, transparent and glassy when young, becoming white with age. Sporothrix-like type: conidiophores macronematous to micronematous, hyaline, produced as aerial mycelia, simple or irregularly or dichotomous branched, producing conidia from denticles in a sporothrix-like fashion. Conidiogenous cells hyaline, smooth, straight or curved, integrated or discrete, terminal or intercalary, cylindrical, tapering towards apex, (18.5–)20–43.5(–73.5) × (1–)1.5–2.5(–3) µm, apex becoming nodose from numerous denticles, often proliferating at the apex and giving rise to another nodose or a conidiogenous cell with nodose at the apex. Conidia solitary, abundant in cultures, more varied to the size and shape of conidia produced in synnemata, hyaline, aseptate, smooth, oblong, obovate, (3–)4–5.5(–8) × (1–)1.5–2(–3) µm.
Culture characteristics.
Colonies with optimal growth at 25 °C on 2% MEA, reaching 66 mm (± 0.59 mm) diam. in 14 d, with a radial growth rate of 0.24 mm/d, followed by 20 °C (55 mm, ± 0.21 mm) diam. Colony olive-brown (1E5), with pale grey (1B1) aerial mycelia at the centre; with age, colonies become olive-grey (1E2); flat, with undulate margin, revers olive (1F4). Hyphae pale yellow (1A3) to olive-yellow (3E6) in colour (Kornerup and Wanscher 1978), smooth, with or without granules, submerged in the medium and aerial mycelium abundant, often constricted at the septa, (1–) 1.5–2.5 (–3) µm wide.
Associated insect.
Tomicus spp.
Host tree.
Pinus sylvestris *
Distribution.
Poland
Additional material examined.
Poland • Lubuskie Province, Babimost, from fallen shoots of Pinus sylvestris pruned by pine shoot beetles, Tomicus sp., October 2023, coll. R. Jankowiak (culture CBS 152111).
Notes.
The newly-described O. babimostense resides in the O. ulmi species complex. Ophiostoma babimostense is morphologically similar to many species of the O. ulmi complex. However, O. babimostense does not form a monophyletic clade with other species in the O. ulmi complex (Figs 1, 3, Suppl. material 1: figs S1, S2). This species fails to group consistently with any one member of the O. ulmi complex within the ITS- (Fig. 1), TUB2- (Suppl. material 1: fig. S1) and TEF1- (Suppl. material 1: fig. S3) based trees. In addition, O. babimostense formed a distinct lineage basal to all other species in the O. ulmi complex in the combined analyses of the TUB2 and TEF1 datasets (Fig. 3).
Sporothrix
europaea
Taxon classificationFungiOphiostomatalesOphiostomataceae
R. Jankowiak & H. Solheim sp. nov.
DA31E7C6-F903-5CE7-B64E-43D831626355
Mycobank No: 858471
Etymology.
The specific epithet “europaea” (Latin) refers to the European continent, where this fungus was isolated in Norway and Poland.
Sporothrix europaea sp. nov. (CBS 151676). a. Ascoma; b. Ascomatal base; c. Ostiolar hyphae; d, e. Ascospores; f, g. Conidiogenous cell with an inflated cluster of denticles at the apex; h. Conidia; i. Globose conidia arising directly from hyphae; j. Fourteen-day-old culture on MEA. Scale bars: 100 (a); 50 μm (b); 25 μm (c); 10 μm (d–i).
Type.
Norway • Ås, from Anisandrus dispar infesting Quercus robur, June 2016, coll. T. Aas, (holotype KRAM F-60036, culture ex-type CBS 151676).
Description.
Sexual and asexual morphs produced on the sterilised beech twigs and on the surface of malt agar in Petri dishes. Sexual morphs: ascomata perithecial, abundant, superficially or partly embedded in the agar, single or in groups. Perithecial bases globose, (86–) 105–160 (–193) μm diam., black, with brown hyphal hairs, 30–84 μm long and 1.3–2.4 μm wide at the base. Perithecial necks black, straight or curved, (257–)336–456(–530) μm long, (8.5–)9.5–13(–14.5) μm diam. at the apex and (22–)26–39(–57.5) μm at the base. Ostiolar hyphae present, pale brown, septate, straight or curved, simple, tips blunting or strongly thickened, 10–19 in number, (14–)29–42.5(–48.5) μm long, 0.5–1.5 μm at the apex and 1–2.5 μm at the base. Asci evanescent. Ascospores one-celled, hyaline, allantoid in side view (3–)3.5–4(–5) x (0.5–)1–1.5(–2) μm, elliptical in front view (2.5–)3–4(–5) × (1–)1–1.5(–1.5) μm, sometimes with residual sheath up to 2 μm thick, accumulated in white-colour mass at the tip of the neck. Asexual morphSporothrix-like: conidiophores hyaline, micronematous, simple or branched and bearing several conidiogenous cells, borne on upright undifferentiated hyphae. Conidiogenous cells cylindrical, terminal or intercalary, straight or curved, tapering towards the apex, (3.5–)15–36(–50.5) μm long, (0.5–)1–2(–2) μm wide at the base, the apical part swollen, (1–)2–3(–3) μm long, (1.5–)2–4(–5.5) μm wide, with multiple conidiogenous loci as denticles, born by sympodial proliferation. Conidia of two types: 1) abundant in cultures, hyaline, unicellular, smooth, variable in shape and size, guttuliform to fusiform, curved, often asymmetric, pointed at the base, (3–)3.5–5(–7) × (0.5–)1–1.5(–2.5) μm, formed directly on denticles; 2) sparse in cultures, directly on the side of submerged hyphae in malt agar, subhyaline to lightly pigmented, unicellular, smooth, subglobose to broadly obovate, (2.5–)3–4(–5.5) × (2–)2–3(–4) μm diam., formed singly.
Culture characteristics.
Colonies with optimal growth at 25 °C on 2% MEA reaching an average of 47 mm (± 0.07 mm) after 14 days, with radial growth rate 1.68 (± 0.24) mm/d, growth somewhat slower at 30 °C (40 mm diameter); white (3A1), flat, floccose, growing in a circular pattern with entire margins, reverse yellowish-white (3A2). Hyphae greenish-grey (1B7) in colour, smooth, with granules, submerged in the medium and aerial mycelium abundant, (0.5–)1–1.5(–3) µm wide.
Associated insects.
Anisandrus dispar, Ips cembrae, Scolytus intricatus, Scolytus rugulosus, Xyleborinus saxesenii, Xyleborus monographus.
Host trees.
Fagus sylvatica, Larix decidua, Prunus domestica, Quercus robur
Distribution.
Norway, Poland
Additional specimen examined.
Poland • Prószków, from Xyleborus monographus infesting Quercus robur, May 2013, coll. P. Wieczorek, (culture CBS 149830=CMW 60554); Norway • Ås, Anisandrus dispar on Quercus robur, June 2016, coll. T. Aas, (culture CBS 151675).
Notes.
This species is phylogenetically distinct from the other Sporothrix species, based on phylogenetic analysis of combined TUB2 and CAL sequence data (Fig. 4). Sporothrix europaea is phylogenetically closely related to S. fusiformis and S. lunata described by Aghayeva et al. (2004). However, S. europaea has smaller ascomatal bases and necks compared to S. fusiformis (86–193 μm and 257–530 μm vs. 121–273 μm and 301–1168 μm). In turn, S. europaea has larger ascomatal bases (86–193 μm vs. 60–178 μm) and longer necks (257–530 μm vs. 162–700 μm) compared to S. lunata (Aghayeva et al. 2004). In addition, S. europaea has two conidial types, guttuliform to fusiform and subglobose to obovate, whereas those in cultures of S. fusiformis are only guttuliform to fusiform.
Spotothrix lunata has lunate conidia flattened at one side or curved with a blunt base (Aghayeva et al. 2004). The cultures of S. europaea grow on 2% MEA between 5 °C and 35 °C, whereas S. fusiformis and S. lunata did not growth below 10 °C and above 30 °C (Aghayeva et al. 2004).
Sporothrix europaea was represented by five isolates collected from Poland. It corresponds to Sporothrix sp. 4 in the study of Jankowiak et al. (2019b). Sporothrix europaea was isolated from hardwoods in association with different bark beetles.
Discussion
In this study, a set of ophiostomatalean isolates from Norway and Poland were shown to represent two new taxa in the Ophiostomatales (Ascomycota), Ophiostoma babimostense and Sporothrix europaea.
Morphological comparisons and multigene phylogenies placed O. babimostense as an additional species within the O. ulmi species complex. The new species described herein exhibit morphological resemblance to the other members of the O. ulmi complex producing pesotum- and sporothrix-like morphs. Most of the members of the complex produce ascomata with long necks and allantoid ascospores that lack sheaths (De Beer and Wingfield 2013; De Beer et al. 2022). Ophiostoma babimostense is not easily distinguished morphologically from other members of the O. ulmi species complex, especially regarding morphology of the asexual morph. However, it is different from most members of the O. ulmi complex by the lack of a sexual morph in culture. Currently no sexual morphs are known for O. hylesinum and O. australiae. The first species was described from Fraxinus excelsior L. infested by bark beetles (Aas et al. 2018), while O. australiae was isolated from wounds on Acacia mearnsii De Wild. trees in Australia (Kamgan Nkuekam et al. 2008).
Twenty-one known species were included previously in the O. ulmi complex (De Beer et al. 2022). The O. ulmi complex includes species what was previously referred to as the ‘hardwood clade’ in the O. piceae complex (Harrington et al. 2001; Linnakoski et al. 2010). It was later considered as the O. quercus complex (Kamgan Nkuekam et al. 2011). Finally, the O. ulmi species complex has been defined by De Beer and Wingfield (2013). The discovery of O. babimostense known hitherto only from conifers was unexpected because members of this complex had been, so far, only found in association with a variety of bark beetles on hardwoods. Nonetheless, recently, two other members of the O. ulmi complex, namely O. borealis and O. pseudokarelicum, have also been recorded from conifer hosts in the western Carpathian (Jankowiak et al. 2017). Furthermore, O. quercus, a widespread species that primarily infects sapstain in hardwood hosts (Taerum et al. 2018), has been isolated also from conifers (Reay et al. 2005; Thwaites et al. 2005; Zhou et al. 2006; Jankowiak 2012; Jankowiak and Bilański 2013a, 2013b; Jankowiak et al. 2017). It seems that some species of the O. ulmi complex can be found in a much broader range of hosts than what is commonly considered and host specificity is not as strict for some species.
Ophiostoma babimostense has been isolated from Scots pine shoots infested by an unknown species of Tomicus, probably T. piniperda (L.) or T. minor (Hart.). This species was not observed in previous studies dealing with associations of Scots pine-infesting Tomicus spp. and fungi in Poland (Jankowiak 2006, 2008; Jankowiak and Bilański 2007), but was reported as an unknown species later on (Jankowiak and Kolařík 2011; labelled in this paper as Ophiostoma sp. 1). The ecological niche of O. babimostense remains incompletely unknown. It can be assumed that it is associated with Tomicus species in Poland although this should be confirmed.
Sporothrix europaea forms part of the S. gossypina complex, bringing the total number of species in the complex to 19. This species complex was introduced by De Beer et al. (2016). Members of the S. gossypina complex are most often associated with bark and ambrosia beetles, although a few species have been isolated from stained oak wood or tree wounds (Aghayeva et al. 2004; De Beer et al. 2022).
Sporothrix europaea was isolated from different beetle species and host trees (Jankowiak et al. 2017, 2019b; labelled in this paper as Sporothrix sp. 4), confirming that it is not host specific. In a Polish study, S. europaea was found in association with Scolytus intricatus and Xyleborus monographus on Quercus robur, Scolytus rugulosus on Prunus domestica and Anisandrus dispar and Xyleborinus saxesenii on Fagus sylvatica (Jankowiak et al. 2019b; Suppl. material 1). In addition, a single isolate of S. europaea was isolated from Ips cembrae on Larix decidua (Jankowiak et al. 2017). The Norwegian isolates of S. europaea were collected from Q. robur infested by A. dispar and Trypodendron domesticum (H. Solheim, unpublished data).
In addition, our data show that S. fusiformis is also commonly associated with diverse species of bark beetles and host trees, including A. dispar, Dryocoetes villosus, S. intricatus, S. ratzeburgi, X. monographus, X. saxesenii and Betula pendula, Fagus sylvatica, Larix decidua, Prunus domestica and Quercus robur (Jankowiak et al. 2019b; species labelled in this paper as Sporothrix sp. 9). This fungus has previously been described by Aghayeva et al. (2004), where it was found in Azerbaijan and Austria occurring on stained wood of Castanea sativa, L. decidua, Populus nigra, Quercus petraea (Aghayeva et al. 2004). In addition, S. fusiformis was previously found in association with Ips cembrae in Austria (Aghayeva et al. 2004).
Species of the S. gossypina complex exhibit a relative morphological homogeneity. Based on DNA sequence comparisons, S. europaea is closely related to S. fusiformis and S. lunata. In addition to phylogenetic differences, these species can be separated from each other, based on characteristics of their sexual and asexual morphs.
The present study provides novel information about the new species from the O. ulmi complex that includes non-pathogenic species like Ophiostoma quercus (Taerum et al. 2018) or the highly aggressive pathogenic fungus O. novo-ulmi, the causal agents of Dutch Elm Disease (DED) (Brasier 1991). The results of this study have also expanded our knowledge of Sporothrix. Broadly, the results suggest that Sporothrix species from the S. gossypina complex are common and ecologically diverse members of the Ophiostomatales in hardwood ecosystems in Europe.
Supplementary Material
XML Treatment for Ophiostoma babimostense
XML Treatment for Sporothrix europaea
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aas T Solheim H Jankowiak R Bilański P Hausner G (2018) Four new Ophiostoma species associated with hardwood-infesting bark beetles in Norway and Poland.Fungal Biology 122: 1142–1158. 10.1016/j.funbio.2018.08.00130449352 · doi ↗ · pubmed ↗
- 2Aghayeva DN Wingfield MJ De Beer ZW Kirisits T (2004) Two new Ophiostoma species with Sporothrix anamorphs from Austria and Azerbaijan.Mycologia 96: 866–878. 10.1080/15572536.2005.1183293321148906 · doi ↗ · pubmed ↗
- 3Bhunjun CS Chen YJ Phukhamsakda C Boekhout T Groenewald JZ Mc Kenzie EHC Francisco EC Frisvad JC Groenewald M Hurdeal VG Luangsa-ard J Perrone G Visagie CM Bai FYBłaszkowski J Braun Ude Souza F Ade Queiroz MB Dutta AK Gonkhom D Goto BT Guarnaccia V Hagen F Houbraken J Lachance MA Li JJ Luo KY Magurno F Mongkolsamrit S Robert V Roy N Tibpromma S Wanasinghe DN Wang DQ Wei DP Zhao CL Aiphuk W Ajayi-Oyetunde O Arantes TD Araujo JC Begerow D Bakhshi M Barbosa RN Behrens FH Bensch K Bezerra JDP Bilański P Bradley CA Bubner B Burgess TI Buyck BČadežN Cai L Calaça FJS Campbell LJ Chaverri P Chen YY Chethana KWT Co · doi ↗ · pubmed ↗
- 4Bilański P Jankowiak R Solheim H Fortuna P ChyrzyńskiŁWarzecha P Taerum SJ (2023) Soil-borne Ophiostomatales species (Sordariomycetes, Ascomycota) in beech, oak, pine, and spruce stands in Poland with descriptions of Sporothrix roztoczensis sp. nov., S. silvicola sp. nov., and S. tumida sp. nov.Myco Keys 97: 41–69. 10.3897/mycokeys.97.9741637251288 PMC 10210257 · doi ↗ · pubmed ↗
- 5Błońska E Lasota J Jankowiak R Michalcewicz J Wojas T Zbyryt A (2021) Biological and physicochemical properties of the nests of White Stork Ciconia ciconia reveal soil entirely formed, modified and maintained by birds. The Science of the Total Environment 763: 43020. 10.1016/j.scitotenv.2020.14302033143925 · doi ↗ · pubmed ↗
- 6Brasier CM (1991) Ophiostoma novo-ulmi sp. nov., causative agent of current Dutch elm disease pandemics.Mycopathologia 115: 151–161. 10.1007/BF 00462219 · doi ↗
- 7Carbone I Kohn LM (1999) A method for designing primer sets for speciation studies in filamentous ascomycetes.Mycologia 91: 553–556. 10.2307/3761358 · doi ↗
- 8Constantinescu O Ryman S (1989) A new Ophiostoma on polypores.Mycotaxon 34: 637–642. 10.5962/p.416750 · doi ↗