Morphological and phylogenetic analyses reveal Xenosporidesmiaceae fam. nov, with seven Xenosporidesmium species

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
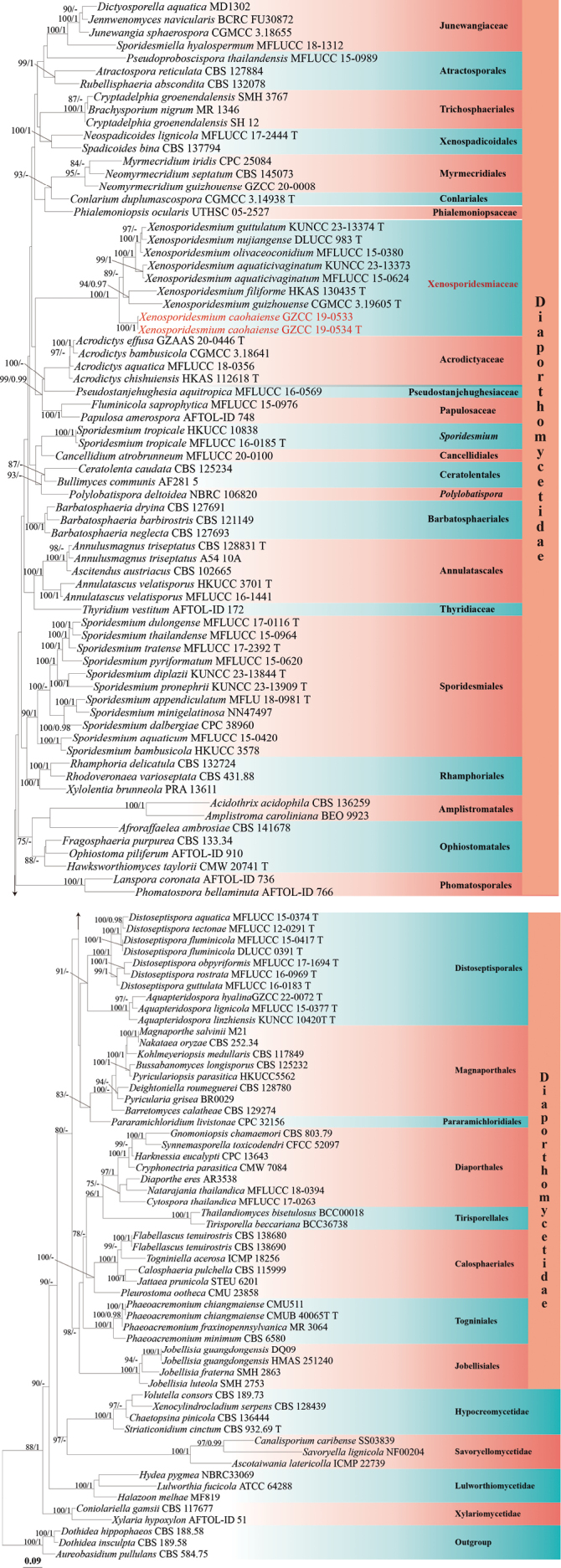 Figure 1
Figure 1 Figure 2
Figure 2| Species | Strain | Type | LSU | SSU | ITS | |
|---|---|---|---|---|---|---|
|
| CBS 136259 |
| NA | NA | NA | |
|
| MFLUCC 18-0356 |
| NA |
| NA | |
|
| CGMCC 3.18641 |
|
|
| NA | |
|
| T |
|
|
|
| |
|
| T |
| NA |
| NA | |
|
| CBS 141678 |
|
| NA | NA | |
|
| BEO 9923 |
| NA | NA | NA | |
|
| MFLUCC 16-1441 |
|
| NA | NA | |
|
| HKUCC 3701 | T |
| NA | NA | NA |
|
| A54 10A |
| NA | NA | NA | |
|
| CBS 128831 | T |
|
| NA |
|
|
| MFLUCC 15-0377 | T |
| NA | NA | NA |
|
| KUNCC 10420 | T |
| NA |
| NA |
|
| T |
| NA |
| NA | |
|
| CBS 102665 |
|
| NA | NA | |
|
| ICMP 22739 |
| NA |
|
| |
|
| CBS 127884 |
| NA | NA |
| |
|
| CBS 584.75 |
|
|
| NA | |
|
| CBS 121149 |
|
| NA |
| |
|
| CBS 127691 |
|
|
|
| |
|
| CBS 127693 |
|
| NA | NA | |
|
| CBS 129274 |
| NA |
| NA | |
|
| MR 1346 |
|
| NA |
| |
|
| AF281.5 |
|
| NA | NA | |
|
| CBS 125232 |
|
|
| NA | |
|
| CBS 115999 |
|
| NA |
| |
|
| SS03839 |
|
|
| NA | |
|
| MFLUCC 20-0100 |
|
| NA | NA | |
|
| CBS 125234 |
|
| NA |
| |
|
| CBS136444 |
| NA | NA | NA | |
|
| CBS 117677 |
| NA | NA | NA | |
|
| CGMCC 3.14938 | T |
|
|
| NA |
|
| CMW 7084 |
|
|
| NA | |
|
| SMH 3767 |
| NA | NA | NA | |
|
| SH 12 |
| NA | NA | NA | |
|
| MFLUCC 17-0263 |
| NA | NA |
| |
|
| CBS 128780 |
| NA |
| NA | |
|
| AR3538 |
| NA | NA | NA | |
|
| MD1302 |
|
| NA | NA | |
|
| MFLUCC 15-0374 | T |
| NA |
| NA |
|
| DLUCC 0391 | T |
| NA |
| NA |
|
| MFLUCC 15-0417 | T |
| NA |
| NA |
|
| MFLUCC 16-0183 | T |
|
|
| NA |
|
| MFLUCC 17-1694 | T |
| NA | NA |
|
|
| MFLUCC 16-0969 | T |
| NA |
|
|
|
| MFLUCC 12-0291 | T |
| NA |
|
|
|
| CBS 188.58 |
| NA |
| NA | |
|
| CBS 189.58 |
|
| NA | NA | |
|
| CBS 138680 |
| NA |
| NA | |
|
| CBS 138690 |
| NA |
| NA | |
|
| MFLUCC 15-0976 |
|
| NA |
| |
|
| CBS 133.34 |
|
| NA | NA | |
|
| CBS 803.79 |
| NA | NA | NA | |
|
| MF819 |
|
| NA | NA | |
|
| CPC 13643 |
| NA | NA | NA | |
|
| CMW 20741 | T |
| NA |
| NA |
|
| NBRC33069 |
|
| NA | NA | |
|
| STEU 6201 |
|
| NA | ||
|
| BCRC FU30872 |
| NA |
| NA | |
|
| SMH 2863 |
| NA | NA | NA | |
|
| HMAS 251240 |
| NA |
| NA | |
|
| DQ09 |
| NA |
| NA | |
|
| SMH 2753 |
| NA | NA | NA | |
|
| CGMCC 3.18655 |
|
|
| NA | |
|
| CBS 117849 |
| NA |
| NA | |
|
| AFTOL-ID 736 |
| NA | NA |
| |
|
| ATCC 64288 |
|
| NA | NA | |
|
| M 21 |
|
| NA | NA | |
|
| CPC 25084 |
| NA |
| NA | |
|
| CBS 252.34 |
| NA |
| NA | |
|
| MFLUCC 18-0394 |
| NA | NA |
| |
|
|
|
| NA |
| ||
|
| CBS 145073 |
| NA |
|
| |
|
| MFLUCC 17-2444 | T |
|
|
|
|
|
| AFTOL-ID 910 |
|
| NA |
| |
|
| AFTOL-ID 748 |
|
| NA |
| |
|
| CPC 32156 |
| NA | NA | NA | |
|
| CMUB 40065 | T |
| NA |
| NA |
|
| CMU511 |
| NA |
| NA | |
|
| MR 3064 |
|
| NA |
| |
|
| CBS 6580 |
|
| NA |
| |
|
| UTHSC 05-2527 |
| NA |
| NA | |
|
| AFTOL-ID 766 |
|
| NA |
| |
|
| CMU 23858 |
|
| NA |
| |
|
| NBRC 106820 |
| NA |
| NA | |
|
| MFLUCC 15-0989 |
|
| NA | ||
|
| MFLUCC 16-0569 |
|
|
| NA | |
|
| BR0029 |
| NA |
| NA | |
|
| HKUCC5562 |
| NA | NA | NA | |
|
| CBS 132724 |
| NA | NA | NA | |
|
| CBS 431.88 |
| NA |
| NA | |
|
| CBS 132078 |
|
| NA |
| |
|
| NF00204 |
|
| NA | NA | |
|
| CBS 137794 |
|
|
|
| |
|
| MFLUCC 18-1312 |
| NA |
|
| |
|
| MFLU 18-0981 | T |
| NA |
| NA |
|
| MFLUCC 15-0420 |
| NA | NA | NA | |
|
| HKUCC 3578 |
| NA | NA | NA | |
|
| CPC 38960 |
| NA |
| NA | |
|
| KUNCC 23-13844 | T |
| NA |
|
|
|
| MFLUCC 17-0116 | T |
| NA |
| NA |
|
| NN47497 |
| NA | NA |
| |
|
| KUNCC 23-13909 | T |
| NA |
|
|
|
| MFLUCC 15-0620 |
| NA |
|
| |
|
| MFLUCC 15-0964 |
| NA |
|
| |
|
| MFLUCC 17-2392 | T |
|
|
|
|
|
| MFLUCC 16-0185 | T |
|
|
|
|
|
| HKUCC 10838 |
| NA | NA | NA | |
|
| CBS 932.69 | T |
| NA |
|
|
|
| CFCC 52097 |
| NA |
|
| |
|
|
|
| NA | NA | ||
|
| AFTOL-ID 172 |
|
| NA |
| |
|
|
|
| NA | NA | ||
|
| ICMP 18256 |
|
| NA |
| |
|
| CBS 189.73 |
| NA | NA | NA | |
|
| CBS 128439 |
| NA | NA | NA | |
|
| MFLUCC 15-0624 |
|
|
|
| |
|
| KUNCC 23-13373 |
|
|
|
| |
|
|
|
|
|
|
|
|
|
|
|
| NA |
|
| |
|
| T |
|
|
|
| |
|
| CGMCC 3.19605 | T |
|
|
|
|
|
| KUNCC 23-13374 | T |
|
|
|
|
|
| DLUCC 983 | T |
|
|
|
|
|
| MFLUCC 15-0380 |
|
|
|
| |
|
| AFTOL-ID 51 |
| NA | NA |
| |
|
| PRA 13611 |
| NA |
|
|
| Taxa | Strain | Conidiophores | Conidiogenous cells | Conidia |
|---|---|---|---|---|
|
| MFLUCC 15-0624 | 60–125 × 4–6 | NA | 49.5–80.5 × 10.5–14,6–10-distoseptate, with a sheath |
|
| 25.5–50 × 3.5–6 | (6–)9–14.5 × 2.5–5.5 | 35–51 × 7.5–11, 8–13-distoseptate, with a sheath | |
|
| 22–54 × 3.5–4.9 µm | 12–16 × 3.6–4.9 µm | 53–100 × 5.8–9.5 µm, mostly 10-distoseptate, with an apical appendage | |
|
| CGMCC3.19605 | 23–59 × 2–4.6 | 13–25 × 3.1–4 | 46–86 × 7–11.4, 6– 10-septate, with a sheath |
|
| KUNCC 23-13374 | 26–55 × 3.4–5 µm | 12– 20 × 3–4.8 µm | 44–84 × 8.7–12 µm,6–14-distoseptate, with a sheath |
|
| DLUCC 983 | 31–51 × 4–5.5 µm | 11.5–16.5 × 4–5 µm | 54–69 × 10–12.5 µm, 10–14-septate, with a sheath |
|
| MFLUCC 15-0380 | 25–55 × 3–4 | 11– 15 × 3–5 | 25–50 × 6–10, 9–16-septate, with a sheath |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Plant Diversity and Evolution · Mycorrhizal Fungi and Plant Interactions
Introduction
Sporidesmium is a common dematiaceous hyphomycetous genus, which is generally recognized as having holoblastic phragmoconidia borne on simple, determinate, or percurrent conidiophores (Ellis 1958; 1971). Since the genus was introduced by Link (1809), numerous taxa have been accommodated to Sporidesmium. In a total of 504 epithets of Sporidesmium listed in Index Fungorum (August 2025), about 83% of them were proposed before 1992. Based on the diagnostic features such as euseptation/distoseptation, percurrent proliferation, and the absence or presence of conidiophores, Subramanian (1992) reassessed Sporidesmium into several genera. Species that have present conidiophores with none or percurrent and irregular proliferations, and euseptate conidia were disposed in Sporidesmium sensu stricto. Other genera, such as Repetophragma with annellate conidiogenous cells bearing euseptate conidia on present conidiophores, and Stanjehughesia with absent conidiophores and euseptate conidia produced on simple conidiogenous cells, were proposed (Subramanian 1992). Following Subramanian’s delimitation, McKenzie (1995) treated thirteen species of sporidesmium-like taxa, including three new species in Ellisembia and one in Sporidesmium. Hernández-Gutiérrez and Sutton (1997) introduced two genera, Imimyces (= Imicles) (Shoemaker and Hambleton 2001) based on distoseptate conidia on lageniform, doliiform, or ovoid proliferating conidiogenous cells, and Linkosia based on absent conidiophores and solitary and distoseptate conidia on simple conidiogenous cells. Wu and Zhuang (2005) merged Imicles into Ellisembia and Penzigomyces into Sporidesmium. The authors also established the genus Novozymia based on its annellidically proliferating conidiogenous cells, which differ from those of Neosporidesmium.
However, Subramanian’s delimitation criteria for Sporidesmium**sensu lato were rejected by Réblová (1999), who discussed the known connections between sexual morphs and their sporidesmium-like asexual morphs, as well as the morphological differences observed when fungi grow in nature and in culture. For example, Stanjehughesia larvata (= Sporidesmium larvatum) (Subramanian 1992) produces macronematous conidiophores when growing in nature but has semi-macronematous or micronematous conidiophores when growing in culture (Réblová 1999). Shenoy et al. (2006) revealed that Sporidesmium and morphologically similar genera are not monophyletic, being distributed across two distinct fungal classes: Dothideomycetes (e.g., Sporidesmiella fusiformisand Sporidesmium obclavatulum) and Sordariomycetes (e.g., Stanjehughesia vermiculataand Linkosia fusiformis). Since then, the polyphyletic nature of sporidesmium-like taxa was demonstrated in many studies (Su et al. 2016; Yang et al. 2018; Liu et al. 2019; Luo et al. 2019).The order Sporidesmiales containing Sporidesmium**sensu stricto is now placed in Diaporthomycetidae, Sordariomycetes (Crous et al. 2018; Hyde et al. 2020; Dong et al. 2021). Currently, 192 Sporidesmium names are listed in Species Fungorum (as of August 2025), with others transferred to more than 50 genera.
During a survey on freshwater fungi in China, two sporidesmium-like taxa were collected and identified on the basis of morpho-phylogenetic evidence. One new family, Xenosporidesmiaceae, is established to accommodate Xenosporidesmium gen. nov. and seven species of Xenosporidesmium.
Materials and methods
Sampling collection, examination, and isolation
Submerged dead wood samples collected from freshwater in Guizhou Province, China, were placed in zip-lock plastic bags, tagged with the collection information (Rathnayaka et al. 2025), and transported to the laboratory. The samples were then incubated in sterile plastic containers lined with moistened filter paper at room temperature for seven days to promote fungal growth. Motic SMZ 168 Series (Motic, Xiamen, China) and Nikon SMZ-171 (Nikon, Tokyo, Japan) dissecting microscopes were used to observe the fungal colonies and fruiting bodies. Fungal structures were examined and photographed using a Nikon ECLIPSE 80i (Nikon, Tokyo, Japan) compound microscope fitted with a Canon 600D/70D (Canon, Tokyo, Japan) digital camera.
Single-spore isolations were performed according to the method described by Senanayake et al. (2020). Germinated conidia were then aseptically transferred to fresh potato dextrose agar (PDA) plates for further cultivation. Dried specimens were deposited in the Herbarium of Cryptogams, Kunming Institute of Botany, Academia Sinica (HKAS), Kunming, China, and the Herbarium of Guizhou Academy of Agriculture Sciences (GZAAS), Guiyang, China. Pure cultures were deposited at the Guizhou Culture Collection (GZCC), Guiyang, China. The names of the new taxa were registered in Index Fungorum (2025) and Fungal Names (https://nmdc.cn/fungalnames/registe) (Wang et al. 2022).
DNA extraction, PCR amplification, and sequencing
Fresh fungal mycelia grown on potato dextrose agar (PDA) were scraped with a sterilized toothpick and transferred to a 1.5 ml microcentrifuge tube for genomic DNA extraction. Genomic DNA was extracted using the Biospin Fungus Genomic DNA Extraction Kit (BioFlux, China), following the manufacturer’s protocol. Four gene regions, LSU, SSU, ITS and rpb2 were amplified using the primer pairs LR0R/LR5, NS1/NS4, ITS5/ITS4and rpb2-5F/ rpb2-7cR, respectively (Liu et al. 1999; Vilgalys and Hester 1990; White et al. 1990). The PCR amplification reactions were performed in a 25 µL reaction volume, comprising 1 µL of DNA, 1 µL of each forward and reverse primer, and 22 µL of 1.1 × T3 Super PCR Mix (Qingke Biotech, Chongqing, China). The conditions for the polymerase chain reaction (PCR) correspond to the reaction conditions reported by Liu et al. (2019). The PCR products were detected by 1% agarose gel electrophoresis, and the sequencing results were provided by Beijing Tsingke Biotechnology Co., Ltd.
Phylogenetic analyses
The sequences incorporated in this study were downloaded from GenBank (Table 1; https://www.ncbi.nlm.nih.gov/). The single gene datasets (LSU, SSU, ITS, and rpb2) were aligned using MAFFT v.7.473 (https://mafft.cbrc.jp/alignment/server/, (Katoh and Standley 2013)) and trimmed using trimAl.v1.2rev59 software (Capella-Gutierrez et al. 2009) with the ‘gappyout’ option. Multiple gene alignments were merged using SequenceMatrix 1.7.8 (Vaidya et al. 2011).
The maximum likelihood (ML) tree was conducted in the IQ Tree webserver (http://iqtree.cibiv.univie.ac.at/) (Nguyen et al. 2014) with 1,000 ultrafast bootstrap (BS) replicates. The substitution model option was set to auto. Maximum likelihood bootstrap supports (ML-BS) equal to or greater than 75% are marked above or below each branch.
Bayesian inferences (BI) were performed using a Markov chain Monte Carlo (MCMC) algorithm in MrBayes 3.2.6 (Ronquist et al. 2012). The best-fit substitution models for each single gene alignment were decided by MrModeltest 2.3 (Nylander 2008) under the Akaike Information Criterion (AIC). Two parallel runs of four simultaneous Markov chains were performed for 10,000,000 generations, or the searches were stopped when the average standard deviation of split frequencies was below 0.01 (stopval = 0.01). Trees were sampled every 1,000^th^ generations. The first 25% of trees were set as burn-in and discarded. The remaining trees were used to calculate posterior probabilities (BYPP). BYPP supports equal to or greater than 0.95 and is marked above or below each branch.
Phylogenetic trees were visualized using FigTree v1.4.4 and subsequently edited in Adobe Illustrator CC 2019 (v23.1.0; Adobe Systems, USA). Photo-plate and scale bars were prepared using Adobe Photoshop CC 2019 (Adobe Systems, USA) and the Tarosoft (R) Image Framework program, respectively.
Phylogenetic results
The phylogenetic placement of the two newly obtained strains was determined through multi-locus phylogenetic analysis. The concatenated sequence matrix comprised 3,292 characters (LSU: 1–822, SSU: 823–1,800, ITS: 1,801–2,273, and rpb2: 2,274–3,292) across 127 taxa. Both ML and BYPP analyses produced congruent topologies. The best ML tree with a final likelihood value of -60836.732 is presented in Fig. 1.
Phylogenetic tree generated from ML analysis based on the combined LSU, SSU, ITS, and rpb2 sequence data. Bootstrap support values for ML (≥ 75%) and BYPP (≥ 0.95) are indicated near their respective nodes. The tree is rooted with Aureobasidium pullulans CBS 584.75, Dothidea hippophaeos CBS 188.58, and Dothidea insculpta CBS 189.58. Newly obtained strains are in red.
Based on the multi-gene phylogenetic analysis (Fig. 1), our isolates (GZCC 19-0533 and GZCC 19-0534) represent a distinct species of Xenosporidesmium, forming a well-supported basal clade within the genus. Notably, this new species clusters with six other taxa—X. aquaticivaginatum, X. filiforme, X. guizhouense, X. guttulatum, X. nujiangense, and X. olivaceoconidium—to comprise a monophyletic lineage that is phylogenetically distinct from Sporidesmiales. Furthermore, this clade is phylogenetically sister to three incertae sedis families in Diaporthomycetidae (Acrodictyaceae, Papulosaceae and Pseudostanjehughesiaceae). Given the consistent molecular divergence and morphological uniqueness, we propose recognition of this clade as a new family, Xenosporidiaceae (Diaporthomycetidae, family incertae sedis).
Taxonomy
Xenosporidesmiaceae
Taxon classificationFungiDistoseptisporalesXenosporidesmiaceae
L. L. Liu & Z. Y. Liu fam. nov.
D226EE3E-473C-5938-8A9D-6604C9B929FB
Fungal Names: FN 572989
Etymology.
Referring to the name of the type genus.
Description.
Colonies on nature substrate effuse, hairy, scattered or in small groups. Conidiophores macronematous, mononematous, solitary, cylindrical, straight or slightly flexuous, dark brown, septate. Conidiogenous cells holoblastic, monoblastic, integrated, terminal, cylindrical, smooth, pale to dark brown. Conidia acrogenous, solitary, obclavate, pale brown to olivaceous, paler at the apex, tapering gradually towards the apex, smooth, straight or curved, distoseptate, with a mucilaginous sheath or apical appendage.
Type genus.
Xenosporidesmium L. L. Liu & Z. Y. Liu, gen. nov.
Notes.
The new family Xenosporidesmiaceae is established to accommodate a distinct lineage comprising six Sporidesmium species and one new species described herein. This lineage forms a highly supported clade (ML/BS = 100%, BYPP = 1) in the multi-gene phylogeny (LSU, SSU, ITS, and rpb2), diverging from both Sporidesmiales and Distoseptisporales. All members share pale brown, sheathed or appendaged conidia and are exclusively associated with freshwater habitats (Hyde et al. 2016; Liu et al. 2019; Wang et al. 2024). The family is proposed to resolve the phylogenetic placement of these morphologically similar yet evolutionarily distinct taxa, and its establishment aims to avoid taxonomic confusion with the currently recognized Sporidesmium**sensu stricto.
Xenosporidesmium
Taxon classificationFungiDistoseptisporalesXenosporidesmiaceae
L. L. Liu & Z. Y. Liu gen. nov.
2AA56D50-D3C9-56D3-AA13-AF0D12398DCD
Index Fungorum: IF904245
Etymology.
“Xeno-” means different, “Xenosporidesmium” reflects that, although this fungus resembles Sporidesmium in morphology, it is genetically distinct and phylogenetically separate.
Type species.
Xenosporidesmium aquaticivaginatum (Jiang Yang & K.D. Hyde) L. L. Liu & Z. Y. Liu, comb. nov.
Description.
Saprobic on dead wood. Sexual morph: Undetermined. Asexual morph: Colonies on nature substrate effuse, hairy, scattered or in small groups. Conidiophores macronematous, mononematous, solitary, cylindrical, straight or slightly flexuous, dark brown, septate. Conidiogenous cells holoblastic, monoblastic, integrated, terminal, determinate or with one percurrent proliferation, cylindrical, smooth, pale to dark brown. Conidia acrogenous, solitary, obclavate, pale brown to olivaceous, paler at the apex, tapering gradually towards the apex, smooth, straight or curved, distoseptate, with a mucilaginous sheath or apical appendage.
Notes.
The new genus Xenosporidesmium is established to accommodate seven species, including one newly discovered taxon and six species previously classified in Sporidesmium. Xenosporidesmium aquaticivaginatum (=Sporidesmium aquaticivaginatum) is designated as the type species because its epithet reflects the aquatic habitat preference characteristic of this genus, although it was published concurrently with X. olivaceoconidium in the same work (Hyde et al. 2016). In this study, we introduce one new Xenosporidesmium species and transfer Sporidesmium aquaticivaginatum, S. filiforme, S. guizhouense, S. guttulatum, S. nujiangens, and S. olivaceoconidium to Xenosporidesmium based on phylogenetic analyses.
Xenosporidesmium
caohaiense
Taxon classificationFungiDistoseptisporalesXenosporidesmiaceae
L. L. Liu & Z. Y. Liu sp. nov.
F19475B5-3072-5237-99D9-BB39FEAA61C6
Index Fungorum: IF904247
Etymology.
The epithet “caohaiense” refers to the type location, Caohai National Nature Reserve.
Xenosporidesmium caohaiense (HKAS 150269, holotype) a, b. Colony on wood; c–f. Conidiophores and conidia; g–k. Conidia; l, m. Conidiophores with conidiogenous cells; n. Germinated conidium; o, p. Culture; o. from above; p. from below. Scale bars: 50 μm (c–f, l, m); 25 μm (g–m, n).
Holotype.
HKAS 150269
Description.
Saprobic on decaying submerged wood in freshwater habitats. Sexual morph: Unknown. Asexual morph: Hyphomycetous. Colonies on natural substrate effuse, hairy, dark brown, scattered or in small groups, velvety. Mycelium mostly immersed, composed of pale to brown, branched, septate, smooth hyphae. Conidiophores macronematous, mononematous, erect, solitary, straight or slightly flexuous, unbranched, cylindrical, dark brown, 1–3-septate, 25.5–50 × 3.5–6 μm (x̄ = 37 × 4.5 μm, n = 20). Conidiogenous cells monoblastic, integrated, terminal, cylindrical or lageniform, pale brown, (6–)9–14.5 × 2.5–5.5 μm (x̄ = 11 × 4 μm, n = 20). Conidia 35–51 × 7.5–11 μm (x̄ = 42 × 9 μm, n = 20), acrogenous, solitary, obclavate, pale brown, straight or slightly flexuous, apically rostrate, hyaline to pale brown, often with a mucilaginous sheath, truncate at the base, 8–13-distoseptate, guttulate, smooth-walled, thin-walled.
Culture characteristics.
Conidia germinating on PDA medium within 12 h. Germ tubes produced from the apex. Colonies on PDA medium reaching 20 mm diam. after 3 weeks at 25 °C, circular, flat, centrally umbonate, with velvety, white to pale yellow mycelium on the surface; in reverse pale yellow with entire margin.
Material examined.
China • Guizhou Province, Bijie City, Weining District, Caohai National Nature Reserve, on decaying wood submerged in a freshwater lake, 4 October 2018, L.L. Liu, 18C-69 (HKAS 150269, holotype; GZAAS 20-0429, isotype), ex-type cultures GZCC19-0534; China • Guizhou Province, Zunyi City, Chishui District, on decaying wood submerged in a freshwater stream, 16 July 2019, J. Yang, CS2-6-1 (HKAS 150268), living cultures GZCC 19-0533.
Notes.
Xenosporidesmium caohaiense forms a distinct lineage and clusters within the Xenosporidesmium clade by high support (ML-BS = 100%, BYPP = 1) in the phylogenetic tree, indicating a close relationship with other Xenosporidesmium. The morphological differences with other Xenosporidesmium species are shown in Table 2. Xenosporidesmium filiforme is distinguished from other Xenosporidesmium species by possessing an apical appendage. Xenosporidesmium caohaiense can be differentiated from all other species (except X. olivaceoconidium) based on the dimensions of its conidiophores, conidiogenous cells,and conidia. Although X. caohaiense and X. olivaceoconidium exhibit highly similar morphological characteristics, they are phylogenetically distant in the tree. Additionally, there are 43 bp differences (313 bp, without gaps) in the ITS region between these two species. Therefore, we introduce Xenosporidesmium caohaiense as a novel species here.
New combinations
Xenosporidesmium
aquaticivaginatum
Taxon classificationFungiDistoseptisporalesXenosporidesmiaceae
(Jing Yang & K.D. Hyde) L. L. Liu & Z. Y. Liu comb. nov.
0866A885-BDDE-5781-AAB1-1F099D2B4546
Index Fungorum: IF904253
≡ Sporidesmium aquaticivaginatum Jing Yang & K.D. Hyde, Fungal Diversity 80: 217 (2016)
Material examined.
Thailand • Prachuap Khiri Khan Province, Hua Hin, stream flowing outside Kaeng Krachan National Park, on submerged wood, 25 December 2014, Jaap van Strien, Site4-44-2 (MFLU 15-1159, holotype); ex-type living culture, MFLUCC15-0624; ibid. (HKAS950 46, isotype).
Xenosporidesmium
filiforme
Taxon classificationFungiDistoseptisporalesXenosporidesmiaceae
(W.P. Wang, H.W. Shen, K.D. Hyde & Z.L. Luo) L. L. Liu & Z. Y. Liu comb. nov.
17056FF3-EB18-5D8A-9126-6D98D7F29631
Index Fungorum: IF904254
≡ Sporidesmium filiforme W.P. Wang, H.W. Shen, K.D. Hyde & Z.L. Luo, Mycosphere15(1): 6617 (2024)
Material examined.
China • Yunnan Province, Wenshan Zhuang and Miao Autonomous Prefecture, Bamei Town, on submerged decaying wood, 7 February 2022, W.P. Wang, S-3826 (HKAS 130435, holotype).
Xenosporidesmium
guizhouense
Taxon classificationFungiDistoseptisporalesXenosporidesmiaceae
(L. L. Liu & Z. Y. Liu) L. L. Liu & Z. Y. Liu comb. nov.
5AD5E2EF-4EE0-54A2-9C5E-1CFB7B56AE94
Index Fungorum: IF904255
≡ Sporidesmium guizhouense L.L. Liu & Z.Y. Liu, Phytotaxa 422(2): 149 (2019)
Material examined.
China • Guizhou Province, Guiyang City, Baihua Lake, 26°39.29'N, 106°32.22'E, on submerged decaying twig, 18 April 2018, Lingling Liu, 18B–40 (HKAS 104644 holotype, GZAAS 19–005 isotype), ex-type living cultures CGMCC 3.19605, GZCC 19–005.
Xenosporidesmium
guttulatum
Taxon classificationFungiDistoseptisporalesXenosporidesmiaceae
(W.P. Wang, H.W. Shen, K.D. Hyde & Z.L. Luo) L. L. Liu & Z. Y. Liu comb. nov.
75E67AEC-61ED-5846-BD27-6A01AC4E5E65
Index Fungorum: IF904256
≡ Sporidesmium guttulatum W.P. Wang, H.W. Shen, K.D. Hyde & Z.L. Luo, Mycosphere 15(1): 6617 (2024)
Material examined.
China • Yunnan Province, Wenshan Zhuang and Miao Autonomous Prefecture, Bamei Town, on submerged decaying wood, 7 February 2022, W.P. Wang, S-3481 (HKAS 131397, holotype), ex-type living cultures CGMCC 3.27021 = KUNCC 23–13374.
Xenosporidesmium
nujiangense
Taxon classificationFungiDistoseptisporalesXenosporidesmiaceae
(D.F. Bao, Hong Y. Su, K.D. Hyde & Z.L. Luo) L. L. Liu & Z. Y. Liu comb. nov.
A3F87369-47AB-52E0-9E1E-A09975CC05AC
Index Fungorum: IF904257
≡ Sporidesmium nujiangense D.F. Bao, Hong Y. Su, K.D. Hyde & Z.L. Luo, J. Fungi 7(no. 669): 15 (2021)
Material examined.
China • Yunnan Province, Nujiang River, on submerged decaying wood, in July 2016, Z.L. Luo, S983 (HKAS 115795, holotype), ex-type living culture, DLUCC 983.
Xenosporidesmium
olivaceoconidium
Taxon classificationFungiDistoseptisporalesXenosporidesmiaceae
(J. Yang & K.D. Hyde) L. L. Liu & Z. Y. Liucomb. nov.
AEE504DB-53CC-5299-AD59-454C2541716E
Index Fungorum: IF904258
≡ Sporidesmium olivaceoconidium Jing Yang & K.D. Hyde, Fungal Diversity 80: 220 (2016)
Material examined.
Thailand • Chiang Rai Province, stream flowing in Tham Luang Nang Non Cave, on submerged wood, 25 November 2014, Jing Yang YJ-14 (MFLU 15-1175, holotype), ex-type living culture, MFLUCC 15-0380, GZCC 16-0008.
Discussion
Fresh samples and sequence data are urgently needed to resolve other sporidesmium-like and polyphyletic genera. For example, Linkosia was phylogenetically placed in three subclasses (Diaporthomycetidae, Hypocreomycetidae, Sordariomycetidae) in Sordariomycetes (Yang et al. 2018). However, its type species, Linkosia coccothrinacis is rarely studied and lacks molecular data, resulting in the natural classification of Linkosia unsettled. Ellisembia is another sporidesmium-like genus that differs from Sporidesmium in having distoseptate conidia (Subramanian 1992). Ellisembia was synonymized with Sporidesmium by Su et al. (2016), but the same with Linkosia, the systematic placement of the type species of Ellisembia, E. coronate is unknown. Thus, the phylogenetic affinities between Ellisembia and Sporidesmium need to be further revealed. Due to the largely morphological similarity of sporidesmium-like species, we strongly recommend the inclusion of molecular evidence when introducing new taxa into this group to prevent taxonomic confusion in the future.
The Sporidesmiaceae was originally established by Fries (1849), and forms a monophyletic group, currently represented by 20 phylogenetic species (Wang et al. 2024). However, neither the original generic type Sporidesmium atrum nor the lectotype S. ehrenbergii was included in phylogenetic analyses due to the lack of sequence data. Notably, several sporidesmium-like families exist, such as Distoseptisporaceae (Su et al. 2016; Yang et al. 2021; Zhang et al. 2022; Shen et al. 2024;), Neomassariaceae (Zhang et al. 2024; Xiao et al. 2025; Sun et al. 2025), Pseudosporidesmiaceae (Crous 2008; Crous et al. 2017), and Xenosporidesmiaceae (this study). The family affiliations of Sporidesmium atrum and S. ehrenbergii need to be further revealed.
Supplementary Material
XML Treatment for Xenosporidesmiaceae
XML Treatment for Xenosporidesmium
XML Treatment for Xenosporidesmium caohaiense
XML Treatment for Xenosporidesmium aquaticivaginatum
XML Treatment for Xenosporidesmium filiforme
XML Treatment for Xenosporidesmium guizhouense
XML Treatment for Xenosporidesmium guttulatum
XML Treatment for Xenosporidesmium nujiangense
XML Treatment for Xenosporidesmium olivaceoconidium
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Capella-Gutierrez S Silla-Martinez JM Gabaldon T (2009) trim Al: A tool for automated alignment trimming in large-scale phylogenetic analyses.Bioinformatics (Oxford, England)25: 1972–1973. 10.1093/bioinformatics/btp 34819505945 PMC 2712344 · doi ↗ · pubmed ↗
- 2Crous PW (2008) Sporidesmium knawiae Crous, sp. nov. Fungal Planet: 29.
- 3Crous PW Wingfield MJ Burgess TI Carnegie AJ Hardy GESJ Smith D Summerell BA Cano-Lira JF Guarro J Houbraken J Lombard L Martín MP Sandoval-Denis M Alexandrova AV Barnes CW Baseia IG Bezerra JDP Guarnaccia V May TW Hernández-Restrepo M Stchigel AM Miller AN Ordoñez ME Abreu VP Accioly T Agnello C Agustin Colmán A Albuquerque CC Alfredo DS Alvarado P Araújo-Magalhães GR Arauzo S Atkinson T Barili A Barreto RW Bezerra JL Cabral TS Camello Rodríguez F Cruz RHSF Daniëls P Pda Silva BD Bde Almeida DA Cde Carvalho Júnior AA Decock CA Delgat L Denman S Dimitrov RA Edwards J Fedosova AG Ferreira RJ Fi · doi ↗ · pubmed ↗
- 4Crous PW Wingfield MJ Burgess TI Hardy GESJ GenéJ Guarro J Baseia IG García D Gusmão LFP Souza-Motta CM Thangavel R Adamčík S Barili A Barnes CW Bezerra JDP Bordallo JJ Cano-Lira J Fde Oliveira RJV Ercole E Hubka V Iturrieta-González I KubátováA Martín MP Moreau PA Morte A Ordoñez ME Rodríguez A Stchigel AM Vizzini A Abdollahzadeh J Abreu VP AdamčíkováK Albuquerque GMR Alexandrova AVÁlvarez Duarte E Armstrong-Cho C Banniza S Barbosa RN Bellanger JM Bezerra JL Cabral TS CaboňM Caicedo E Cantillo T Carnegie AJ Carmo LT Castañeda-Ruiz RF Clement CRČmokováA Conceição LB Cruz RHSF Damm Uda Silv · doi ↗ · pubmed ↗
- 5Dong W Hyde KD Jeewon R Doilom M Yu XD Wang GN Liu NG Hu DM Nalumpang S Zhang H (2021) Towards a natural classification of annulatascaceae-like taxa II: Introducing five new genera and eighteen new species from freshwater.Mycosphere 12: 1–88. 10.5943/mycosphere/12/1/1 · doi ↗
- 6Ellis MB (1958) Clasterosporium and some allied dematiaceae-phragmosporae. I.Mycological Papers 70: 1–89.
- 7Ellis MB (1971) Dematiaceous hyphomycetes.Commonwealth Mycological Institute, Kew, Surrey, England, 594 pp. 10.1079/9780851986180.0000 · doi ↗
- 8Fries EM (1849) Summa vegetabilium Scandinaviae II. Sectio posterior, Upsaliae.