Three new species and two new records of the genus Laccaria (Agaricales, Basidiomycota) from subtropical China based on morphological and multi-locus phylogenetic evidence

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2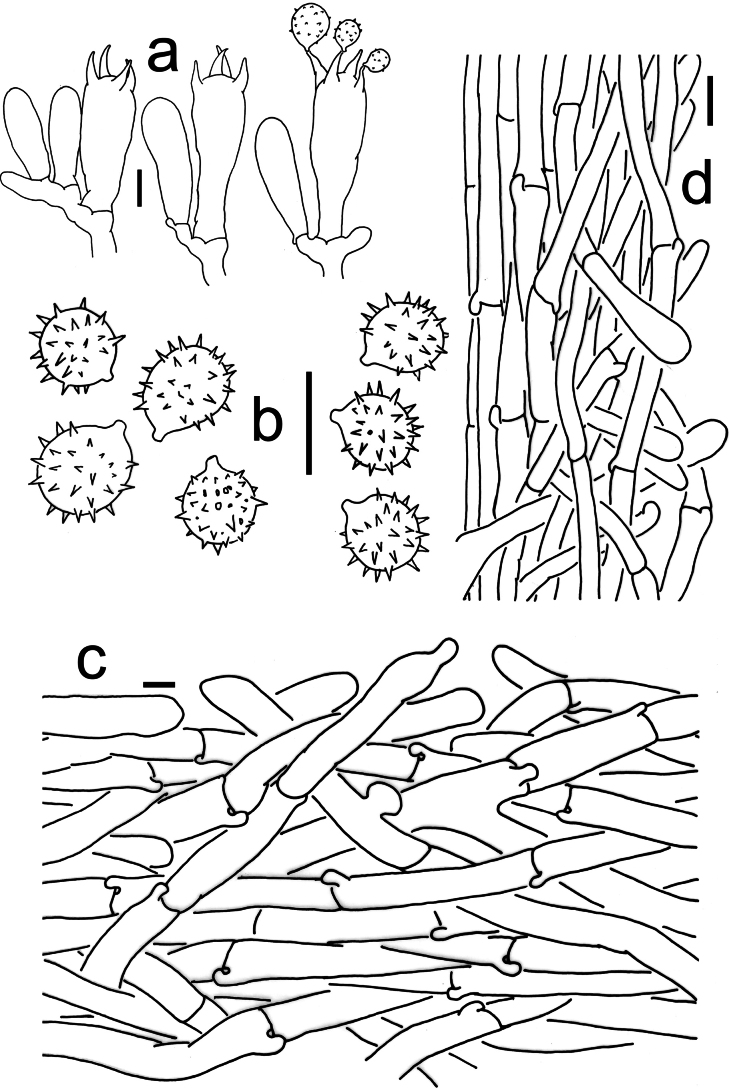 Figure 3
Figure 3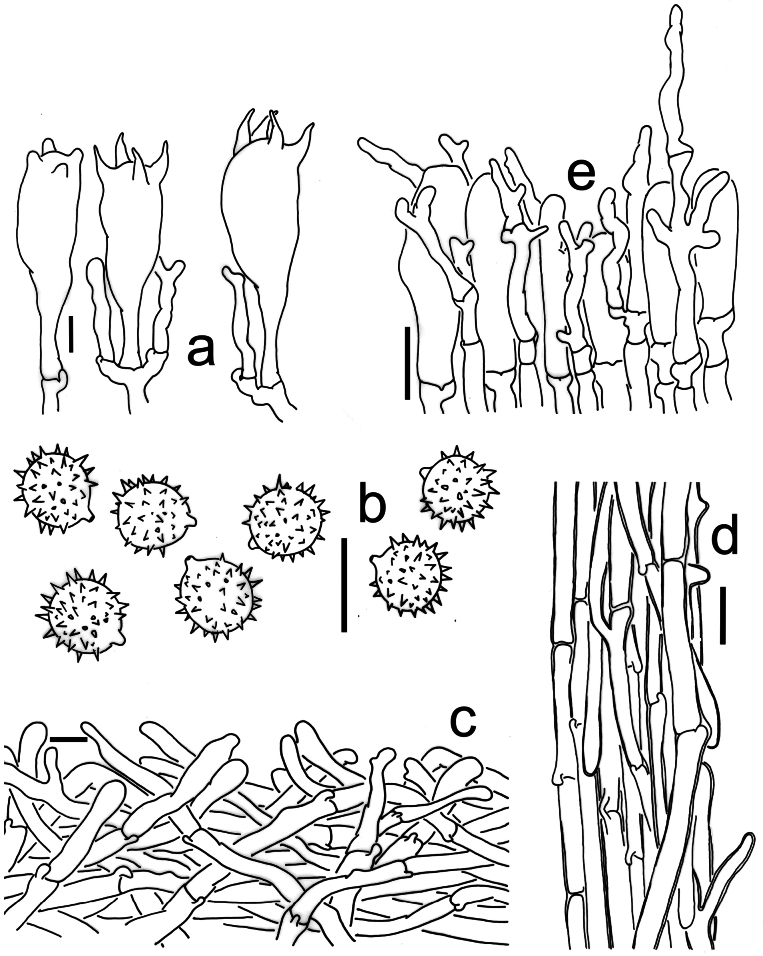 Figure 4
Figure 4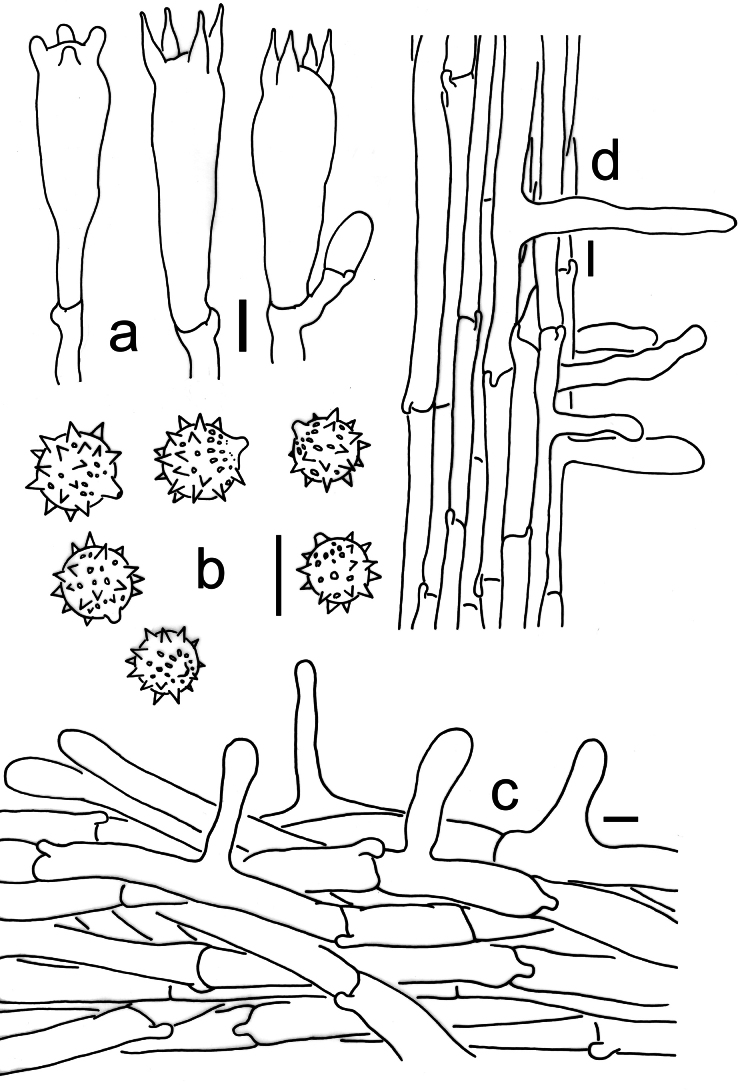 Figure 5
Figure 5 Figure 6
Figure 6| Species | voucher | locality |
| 28S |
|
| References |
|---|---|---|---|---|---|---|---|
| AWW485 | Tibet, China |
|
|
|
|
| |
|
| Yunnan, China |
|
|
|
| This study | |
|
| HKAS45998 | Tibet, China |
|
|
|
| |
|
| AWW438 | Yunnan, China |
|
|
|
|
|
|
| F1121461 | China |
|
|
| ||
|
| TPML20120807-69 | Korea |
|
|
|
|
|
|
| Hunan, China |
|
| This study | |||
|
| Guizhou, China |
|
|
| This study | ||
| New Zealand |
|
|
|
|
| ||
|
| AWW556 | USA |
|
|
|
| |
|
| AWW590 | USA |
|
|
|
| |
|
| GMM7633 | France |
|
|
| ||
|
| GMM7041 | Russia |
|
|
|
| |
|
| KHLA06002 | USA |
|
|
|
|
|
|
| GMM7621 | France |
|
|
|
|
|
|
| HKAS58714 | Yunnan, China |
|
|
| ||
|
| KNU20120912-25 | Korea |
|
|
|
|
|
|
| Hunan, China |
|
|
|
| This study | |
|
| Hunan, China |
|
|
|
| This study | |
| KNU20120912-40 | Korea |
|
|
|
|
| |
|
| MB-FB-001109 | Yunnan, China |
|
| |||
|
| GMM6172 | Yunnan, China |
|
|
|
| |
|
| Hubei, China |
|
|
|
| This study | |
|
| Guizhou, China |
| This study | ||||
| KUN-F 78557 | Yunnan, China |
|
| ||||
|
| KUN-HKAS123246 | Yunnan, China |
|
|
|
|
|
| KUN-HKAS123244 | Yunnan, China |
|
|
|
|
| |
|
| Korea |
|
|
|
|
| |
|
| AWW537 | USA |
|
|
| ||
|
| GMM7620 | France |
|
|
| ||
|
| HKAS44062 | Yunnan, China |
|
|
| ||
|
| F1121424 | China |
|
|
|
| |
|
| GMM6131 | China |
|
|
|
|
|
|
| AWW596 | USA |
|
|
| ||
|
| GMM6094 | China |
|
|
|
| |
|
| GMM2692 | Chile |
|
|
| ||
|
| A0103 | Japan |
|
| Direct Submission | ||
|
| Jiangxi, China |
|
|
|
| This study | |
|
| AWW539 | USA |
|
|
| ||
|
| KUN-HKAS 123243 | Yunnan, China |
|
|
|
|
|
|
| Guizhou, China |
|
|
|
| This study | |
| KUN-HKAS 123286 | Yunnan, China |
|
|
|
|
| |
| AWW465 | Tibet, China |
|
|
|
| ||
|
| GMM7209 | Australia |
|
|
|
| |
|
| GMM7227 | Australia |
|
|
|
|
|
|
| Hunan, China |
|
| This study | |||
|
| Hunan, China |
|
|
| This study | ||
|
| Yunnan, China |
|
|
|
| This study | |
| Hunan, China |
|
|
| This study | |||
|
| A3394 | Japan |
|
|
| Direct Submission | |
|
| A2987 | Japan |
|
|
| Direct Submission | |
|
| AWW555 | USA |
|
|
|
| |
|
| KUN-HKAS83381 | Yunnan, China |
|
|
|
|
|
| KUN-HKAS80885 | Yunnan, China |
|
|
| |||
| CUHAM788 | INDIA |
|
| ||||
|
| KUN-HKAS107731 | Yunnan, China |
|
|
|
|
|
|
| Yunnan, China |
|
|
|
| This study | |
| KUN-HKAS90435 | Yunnan, China |
|
|
|
| ||
|
| KUN-HKAS106741 | Guangdong, China |
|
| |||
| KUN-HKAS106739 | Guangdong, China |
|
|
| |||
|
| GMM7508 | New Zealand |
|
|
|
| |
| KUN-F78556 | Yunnan, China |
|
| ||||
|
| F1081213 | Chile |
|
|
|
|
|
|
| F1080983 | Argentina |
|
|
|
|
|
|
| GMM7521 | New Zealand |
|
|
|
|
|
|
| GMM7534 | New Zealand |
|
|
| ||
|
| F1104722 | Costa Rica |
|
|
| ||
|
| HMAS352266 | Guizhou, China |
|
| |||
| HMAS352265 | Guizhou, China |
|
| ||||
|
| AWW463 | Tibet, China |
|
|
|
| |
| AWW484 | Tibet, China |
|
|
|
| ||
| CUHAM786 | INDIA |
|
| ||||
|
| SFC20110921-34 | Korea |
|
|
|
| |
|
| HMHHNU 9589 | Guizhou, China |
|
| This study | ||
|
| Hubei, China |
|
| This study | |||
|
| Hunan, China |
|
| This study | |||
| TNS- | Japan |
|
| ||||
|
| GMM7615 | France |
|
|
| ||
|
| SB2067 | Portugal |
|
|
|
| |
|
| SB2133 | Portugal |
|
|
|
|
|
|
| SB2210 | Portugal |
|
|
| ||
|
| GMM7586 | Russia |
|
|
|
| |
|
| Cripps1603 | USA |
|
| |||
|
| GMM7605 | France |
|
|
|
|
|
|
| GMM7220 | Australia |
|
|
|
| |
|
| GMM7531 | New Zealand |
|
|
|
|
|
|
| F1092175 | USA |
|
|
| ||
|
| KUN-HKAS123799 | Yunnan, China |
|
|
|
|
|
| KUN-HKAS123801 | Yunnan, China |
|
|
| |||
|
| GMM7612 | France |
|
|
|
|
|
|
| GMM7626 | France |
|
|
|
|
|
|
| GMM6019 | Costa Rica |
|
|
|
|
|
|
| Hunan, China |
|
|
|
| This study | |
| Hunan, China |
|
|
|
| This study | ||
|
| GMM7473 | New Zealand |
|
|
|
|
|
| GDGM76043 | China |
|
|
| |||
|
| TWO591(MONT) | - |
|
| |||
|
| TWO319(MONT) | North America |
|
| |||
|
| AWW446 | Tibet, China |
|
|
|
|
|
|
| GMM7630tibet | Tibet, China |
|
|
|
|
|
|
| Yunnan, China |
|
|
|
| This study | |
| KUN-HKAS93732 | Yunnan, China |
|
| ||||
|
| Hunan, China |
|
|
| This study | ||
|
| ASIS2021 | Korea |
|
| |||
|
| ASIS24249 | Korea |
|
|
|
|
|
|
| GDGM84949 | China |
|
|
|
|
|
| GDGM84954 | China |
|
|
|
|
| |
|
| GMM7631tibet | Tibet, China |
|
|
|
|
|
| BAP360 | Tibet, China |
|
| ||||
|
| GDGM53063 | Jiangxi, China |
|
|
| ||
| GDGM52852 | Jiangxi, China |
|
|
| |||
|
| F1120629 | China |
|
|
| ||
|
| AWW584 | USA |
|
|
|
| |
|
| ObiFr | France |
|
| |||
|
| PRL3777 | USA |
|
|
|
| |
|
| PRL4777 | USA |
|
|
|
| |
|
| AFTOL447 | - |
|
|
| ||
|
| AWW545 | USA |
|
|
|
| |
|
| GMM7539 | New Zealand |
|
|
|
|
|
|
| KH_07192006_1 | USA |
|
|
|
| |
|
| HKAS53170 | Yunnan, China |
|
|
|
|
|
| HKAS107730 | Yunnan, China |
|
| ||||
| CUHAM787 | INDIA |
|
| ||||
| SFC20120919-40 | Korea |
|
| ||||
|
| KUN-HKAS106745 | Guangdong, China |
|
| |||
| KUN-HKAS106742 | Guangdong, China |
|
| ||||
| GMM7628 | France |
|
|
|
|
| |
|
| GMM7584 | Russia |
|
|
|
|
|
|
| F1133825 | USA |
|
|
|
| |
|
| F1081079 | Argentina |
|
|
|
| |
|
| HKAS-110664 | Thailand |
|
|
|
|
|
| MFLU-22-0106 | Thailand |
|
|
|
| ||
| pse1625 | USA |
|
| ||||
|
| GMM7637 | France |
|
|
|
| |
|
| pum1252 | North America |
|
| |||
| LM5099 | Mexico |
|
| Montoya et al. (2015) | |||
|
| KUN-HKAS123292 | Yunnan, China |
|
|
|
|
|
| KUN-HKAS123291 | Yunnan, China |
|
|
|
|
| |
|
| HKAS90758 | Yunnan, China |
|
| |||
|
| Yunnan, China |
|
|
|
| This study | |
| HKAS90753 | Yunnan, China |
|
| ||||
|
| GDGM89627 | Yunnan, China |
|
|
| ||
| GDGM82878 | Yunnan, China |
|
|
|
|
| |
|
| GMM7602tibet | Tibet, China |
|
|
| ||
| GMM7596tibet | Tibet, China |
|
|
|
|
| |
|
| Yunnan, China |
|
|
|
| This study | |
| Yunnan, China |
|
|
|
| This study | ||
| AWW591 | USA |
|
|
| |||
| GMM6012 | Costa Rica |
|
|
|
| ||
| GMM6800 | Guatemala |
|
|
|
|
| |
| ALB183 | China: Tibet |
|
|
|
|
| |
| F1123822 | USA |
|
|
|
| ||
| AWW569 | USA |
|
|
|
| ||
| GMM7627 | France |
|
|
|
| ||
| GMM7020 | Russia |
|
|
|
| ||
| GMM6585 | Costa Rica |
|
|
| |||
| T173 | China |
|
| Direct Submission | |||
| H160 | China |
|
| Direct Submission | |||
| A1800 | Taiwan, China |
|
| ||||
| T44 | China |
| Direct Submission | ||||
| T110 | China |
|
| Direct Submission | |||
| GMM6679 | Yunnan, China |
|
|
|
|
| |
| TWO1166 | Thailand |
|
|
|
| ||
| DED7426 | Thailand |
|
|
|
|
| |
| TWO1168 | Thailand |
|
|
|
|
| |
| TWO1150 | Thailand |
|
|
|
| ||
| TWO1178 | Thailand |
|
|
|
|
| |
| ZT9196 | Indonesia |
|
|
|
| ||
| GMM6776 | Yunnan, China |
|
|
| |||
| HKAS-83382 | China |
|
| Direct Submission | |||
| T107 | China |
|
| Direct Submission | |||
| T168 | China |
|
| Direct Submission | |||
| TWO1184 | Thailand |
|
|
|
| ||
| TWO1194 | Thailand |
|
|
|
| ||
| GMM6583 | China |
|
|
|
|
| |
|
| KUN-HKAS122272 | Yunnan, China |
|
|
|
| |
| KUN-HKAS129615 | Yunnan, China |
|
|
|
| ||
|
| DM93 | Mexico |
|
|
| ||
| DM63 | Mexico |
|
|
| |||
|
| MB-002397 | Panama |
|
| |||
| MB-002396 | Panama |
|
| ||||
|
| Yunnan, China |
|
|
|
| This study | |
|
| Yunnan, China |
|
|
|
| This study | |
|
| Yunnan, China |
|
|
|
| This study | |
|
| KUN-HKAS123285 | Yunnan, China |
|
|
|
|
|
| KUN-HKAS123300 | Yunnan, China |
|
|
|
|
| |
|
| MFLU23-0340 | Thailand |
|
|
| ||
| MFLU23-0339 | Thailand |
|
|
| |||
|
| F1080957 | Germany |
|
|
| ||
| SFC2015090217 | Korea |
|
|
|
|
| |
|
| GMM7635 | France |
|
|
|
|
|
|
| F1116205 | USA |
|
|
| ||
| ASIS22273 | Korea |
|
|
|
|
| |
|
| GMM7733 | USA |
|
|
|
| |
|
| F1111951 | Costa Rica |
|
|
|
| |
| TENN42523 | USA |
|
| ||||
|
| PRL7587 | China |
|
|
|
|
|
|
| WCG2072 | - |
|
|
|
| |
|
| GDGM82883 | China |
|
|
|
|
|
| GDGM82911 | China |
|
|
|
|
| |
|
| SFC20121010-51 | Korea |
|
|
| ||
|
| TPML20121008-03 | Korea |
|
|
|
|
|
|
| ASIA20939 | Korea |
|
|
|
|
|
|
| Hunan, China |
|
|
| This study | ||
| SFC20120926-01 | Korea |
|
|
|
|
| |
|
| A0559 | Japan |
|
|
| Direct Submission | |
|
| SFC20150810-10 | Korea |
|
|
|
|
|
|
| KH_LA06_018 | USA |
|
|
| ||
|
| GMM7580 | New Zealand |
|
|
|
| |
|
| GMM7520 | New Zealand |
|
|
|
| |
| CAL1389 | India |
|
| ||||
|
| HKAS-110636 | Thailand |
|
|
|
|
|
| KUN-F78558 | Yunnan, China |
|
| ||||
|
| AFTOL972 | Germany |
|
|
|
| DirectSubmission |
|
| ES11.10.2. A | Germany |
| DirectSubmission |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Plant and Fungal Species Descriptions · Lichen and fungal ecology
Introduction
The genus Laccaria Berk. & Broome is a group of ecologically significant ectomycorrhizal fungi> that inhabit soil (Wilson et al. 2016, 2017). The latest DNA-based phylogenetic evidence supports its placement within the family Hydnangiaceae Gäum. & C.W. Dodge, suborder Agaricineae Fries, order Agaricales Underw (Vizzini et al. 2024). The diagnostic characters for Laccaria include collybioid to omphaloid basidiomata, vividly orange to brown or purple pileus with thick, sparsely arranged lamellae, and globose to subglobose echinulate basidiospores (Berkeley and Broome 1883; Mueller 1984, 1991a; Mueller and Vellinga 1986). Ecologically, species of Laccaria always form symbiotic relationships with trees, such as Pinaceae Lindl., Myrtaceae Juss., Salicaceae Juss., Fagaceae Candolle., Dipterocarpaceae Blume., Nothofagaceae Ørsted. and a part of Fabaceae Lindl. (Mueller1991; Wilson et al. 2017). The importance of this genus lies in its ability to provide nutrients to its plant partners and to engage in nutrient cycling, which is crucial for forest function and stability (Simard 2009; van der Heijden et al. 2015; Wilson et al. 2017). Furthermore, the basidiomata of some Laccaria are known to be edible, e.g., L. alba Zhu L. Yang & L. Wang, L. laccata Cooke, L. amethystina Cooke, L. angustilamella Zhu L. Yang & L. Wang, L. aurantiaF. Popa et al, L. bullipellis A.W. Wilson & G.M. Muell., and L. himalayensis A.W. Wilson & G.M. Muell (Guzmán 2016; Li et al. 2015; Wu et al. 2019; Mao 2020; Wang et al. 2022).
Since Laccaria was established by Berkeley and Broome (1883), numerous mycologists have contributed to its taxonomy. To date, ca. 120 species have been reported worldwide (Berkeley and Broome 1883; Singer 1967; Besson and Kühner 1971; Mueller and Sundberg 1981; Osmundson et al. 2005; Popa et al. 2016; Campi et al. 2017; Ramos et al. 2017; Corrales et al. 2020; Cui et al. 2021; Dovana et al. 2021), with the majority discovered in Europe, North America. Recently, the number of Laccaria species recognized in Asia is increasing, driven by growing awareness of the ecological and economic importance of this genus (Cho et al. 2018, 2020; Deepna Latha et al. 2019; Tang et al. 2024; Thapa et al. 2024). In China, 25 new species of Laccaria have been described since the year 2000 (Wang et al. 2004; Wilson et al. 2013; Popa et al. 2014; Luo et al. 2016; Vincenot et al. 2017; Li 2020; Cui et al. 2021; Wang et al. 2022; Zhang et al. 2023; Li et al. 2024). They were found in Southern China, a region whose biodiversity is reflected in its diverse forest ecosystems. Furthermore, most of them were discovered from Yunnan Province. However, studies on the diversity of Laccaria taxa in other provinces of Southern China remain limited. In this study, (i) two new species from Hunan Province and one new species from Yunnan Province are proposed based on both molecular and morphological evidence; (ii) Chinese specimens of L. japonica Popa & K. Nara and L. versiforma H.J. Cho & Y.W. Lim, collected from Guizhou, Hubei and Hunan Provinces, are reported for the first time.
Materials and methods
Specimen collection
A total of 28 specimens of Laccaria were involved in this study. These specimens were collected from multiple locations in Southern China, including Yunnan, Guizhou, Hubei, Jiangxi, and Hunan Provinces, between 2016 and 2024. After dehydration using heat or silica gel, they were deposited in the Mycological Herbarium of Hunan Normal University (MHHNU, Changsha, China). Information regarding the specimens, including species name, GenBank accession number, voucher and location, is provided in Table 1.
Morphological study
Macroscopic characters of species were described based on field notes and digital images. The size of basidiomata, as determined by pileus width, was described as tiny (<1.5 cm), small (1.5–3 cm), medium-sized (3–5 cm) or large (>5 cm). The color codes mentioned in descriptions are from Kornerup and Wanscher (1978). For microscopic studies, hand-made sections of dried basidiomata were prepared under a stereomicroscope to ensure precise dissection of specific tissues. Microscopic examinations were then conducted using a light microscope. The dried specimens mounted in either a 5% KOH solution or distilled water, with Congo red staining when necessary. Melzer’s reagent was used to test the amyloidity of basidiospores. With preheating, Cotton blue reagent was to test the cyanophily of basidiospores. Basidiospores, basidia, pileipellis, stipitipellis and cystidia were illustrated by hand drawing.
In the description of basidiospores, the abbreviation [n/m/p] represents ‘n’ basidiospores measured from ‘m’ basidiomata of ‘p’ collections. Dimensions for basidiospores are given using notation of the form (a) b–c (d), while ‘a’ and ‘d’ mean the minimum and maximum, respectively, and ‘b–c’ contains a minimum of 90% of the measured values. The Q value represents the length/width ratio of a basidiospore inside view, and Qm value indicates average Q ± standard deviation. The terminology for denoting the shape of basidiospores followed Bas (1969).
The scanning electron microscopy (SEM) was also applied to observe the basidiospores ornamentation. Fragment of dry lamellae tissue samples were securely fastened to aluminum stubs and coated gold palladium before observed under a TESCAN CLARA Xplore 30 (Brno, Czech Republic) SEM.
DNA extraction, PCR amplification and sequencing
Total genomic DNA was extracted using the Fungal DNA Mini Kit (Omega Bio-Tek, Norcross, USA) following the manufacturer’s instructions. For the PCR amplifications, the following primers were employed: (1) ITS5 and ITS4 (White et al. 1990) were used for the internal transcribed spacer (ITS); (2) LR0R and LR5 (Vilgalys and Hester 1990) for the nuclear ribosomal large subunit (LSU); (3) EF1-983F, EF1-1953R and EF1-1567R (Matheny et al. 2007), or the newly designed primers EF1-Laccaria-F1 (5’-ATGGACACCACCAAGGTAAGA-3’) and EF1-Laccaria-R1 (5’-ACGTTGCCACG.
ACGAATAT-3’), EF1-Laccaria-F2 (5’-TGCCTTTGTCCCTATTTCCG-3’) and EF1-Laccaria-R2 (5’-GGGTGGTTGAGGACGATGAC-3’) for the translation elongation factor 1-α (TEF1); (4) bRPB2-6F and bRPB2-7.1R (Matheny 2005), or the newly designed primers RPB2-Laccaria-F1 (5’-ACCATCACAAACGGTCTCA-3’) and RPB2-Laccaria-R1 (5’-CACCCTTTACCAGATGTTCC-3’), RPB2-Laccaria-F (5’-CTGAAGGTCAAGCCTGTGG-3’) and RPB2-Laccaria-R (5’-ACTTTGCTGTAGGC.
GAGAAT-3’) for polymerase II second largest subunit (RPB2). The new primers were designed using Primer 5.0.
The PCR mixtures were composed of 1 × PCR buffer, 1.5 mM MgCl_2_, 0.2 mM dNTPs, 0.4 μm forward primer, 0.4 μm reverse primer, 1.25U of Taq polymerase (CWBIO, Jiangsu, China), and 1 μL of DNA template in a total volume of 25 μL. Amplification reactions were performed with the following program: initial denaturation at 94 °C for 5 min, 35 cycles at 94 °C for 30 s, 52 °C (LSU and TEF1) or 54 °C (ITS and RPB2) for 30 s, and 72 °C for 30 s (ITS and LSU) or 45 s (RPB2 and TEF1), and a final extension at 72 °C for 8 min (He et al. 2023). The products were subjected to electrophoresis on a 2% agarose gel, and the positively identified ones were sent to the Changsha branch of Youkang Biotechnology Co., Ltd. (Zhejiang, China) for sequencing.
Sequence alignment and phylogenetic analysis
The sequences (Table 1) were aligned with the strategy FFT-NNS-I in the software MAFFT v7.511 (Katoh and Standley 2016). As shown in Table 1, a total of 94 sequences (28 ITS, 23 LSU, 19TEF1 and 24RPB2) were newly generated in the present study. The intronic regions of TEF1 and RPB2 were manually excised. The ambiguously aligned regions of ITS and LSU were removed in Gblocks v0.91b (Castresana 2000).
A four-locus matrix for Laccaria (Suppl. material 1) was generated by SEQUENCEMATRIX 1.7.8 (Vaidya et al. 2011). The alignment contains 3423 positions from 215 samples, partitioned as follows: 1–631 (ITS), 632–1,510 (LSU), 1,511–2,402 (TEF1), and 2,403–3,423 (RPB2). The alignment has been deposited in TreeBASE (http://www.treebase.org/treebase/) under submission ID 32143. According to the AIC criterion in MRMODELTEST v2.4 (Nylander 2004), GTR+I+G was selected as the best-fit model for the four loci. Bayesian inference (BI) analysis was performed in MRBAYES v3.2.7 (Ronquist and Huelsenbeck 2003), with the GTR+I+G model for each partition, two simultaneous runs, four Markov Chain Monte Carlo (MCMC) chains, and sampling every 100 generations. After 20 million generations, the standard deviation of split frequencies was below 0.01. The first 25% generations were discarded as burn-in and the convergence was visually assessed by TRACER v1.7.2 (Rambaut et al. 2018). ML analysis with 1000 bootstrap replicates was computed in RAXML v8.0.20 (Stamatakis 2014), using the GTR+I+G model for each partition. Separate phylogenetic analyses based on the four loci were also conducted to assess potential conflicts, with the procedure described above.
Results
Phylogenetic analysis
The phylogenetic trees of the genus Laccaria were constructed based on a four-locus matrix (ITS-LSU-TEF1-RPB2). As the topologies resulting from ML and BI analyses are consistent, only the ML tree is displayed (Fig. 1). Phylogenetic trees based on ITS, LSU, TEF1 and RPB2, respectively, are provided in Suppl. material 2: figs S2–S5, which show no conflicts with the four-locus tree.
ML analysis of Laccaria based on ITS-LSU-TEF1-RPB1 sequence data. Bootstrap values (BP) ≥ 50% from ML analysis and Bayesian posterior probabilities (PP) ≥ 0.90 from BI analysis are shown at nodes. Newly generated sequences are highlighted in bold. GenBank accession numbers of sequences and their geographic origins are shown. Taxa marked with (T) represent type specimens.
According to the phylogenetic tree, 87 species were recognized. The samples of the three newly discovered species described below, namely L. carminostipes (Species 38, 99% BP, 1.00 PP), L. mangshanensis (Species 41, 94% BP, 0.94 PP), and L. sinolateritia (Species 67, 98% BP, 1.00 PP) formed three independent clades. Laccaria carminostipes (MHHNU 11944,31552, 31553 and 34706) was recovered as sister (60% BP, 0.93 PP) to the clade (60% BP, 0.91 PP) containing L. darjeelingensis A. Thapa & K. Acharya, L. fagacicola Y.Y. Cui et al., and L. aurantiaca S.M. Tang et al. These species further formed a clade (60% BP, 0.93 PP), as sister (86% BP, 1.00 PP) to the clade (59% BP, 0.94PP) containing L. aurantia and L. rubroalba X. Luo et al. Laccaria mangshanensis (MHHNU 8850 and 8856) and the Chinese sample A1800 formed a clade with strong support (96% BP, 1.00 PP). Laccaria sinolateritia (MHHNU 11956 and MHHNU 11958) clustered together with L. subroseoalbescens S.M. Tang & S.H. Li with very low support (40% BP, 0.99 PP). Our Chinese samples MHHNU 9589, 34710, and 34711 formed a strongly supported clade with the holotype of L. japonica with minimal genetic distance (Species 19, 90% BP, 1.00 PP), which indicated that they should belong to the same species. Similarly, the Chinese sample MHHNU 10896 clustered with four Korean samples of L. versiforma (Species 46, 99% BP, 1.00 PP), which indicated that they could be conspecific.
The other samples analyzed in this work matched seven known species: L. alba (Species 1, 100% BP, 1.00 PP), L. bicolor (Maire) P.D. Orton (Species 15, 93% BP, 1.00 PP), L. araneosa H.J. Cho & Y.W. Lim (Species 17, 93% BP, 1.00 PP), L. moshuijun F. Popa & Zhu L. Yang (Species 20, 96% BP, 1.00 PP), L. murina S. Imai (Species 33, 99% BP, 1.00 PP), L. fagacicola (Species 37, 88% BP, 1.00 PP), L. aurantia (Species 39, 84% BP, 1.00 PP), L. rubroalba (Species 40, 99% BP, 1.00 PP), L. stipalba S.M. Tang et al. (Species 48, 100% BP, 1.00 PP), and L. brunnea S.M. Tang et al. (Species 54, 100% BP, 1.00 PP).
Taxonomy
Laccaria
carminostipes
Taxon classificationFungiAgaricalesHydnangiaceae
Y.D. Xu & Z.M. He sp. nov.
C47C7829-3954-5964-ACF8-5E7AC8AC03FE
Fungal Names: FN 572506
Etymology.
‘carminostipes’ (Latin), referring to the carmine stipe surface.
Basidiomata of Laccaria carminostipes (a. MHHNU 31553; b. MHHNU 31552; c. MHHNU 34706, respectively); L. mangshanensis (d, e. MHHNU 8850; f. MHHNU 8856, respectively); L. sinolateritia (g, h. MHHNU 11956; i. MHHNU 11958, respectively); L. japonica (j. MHHNU 9589; k. MHHNU 34711, respectively); L. versiforma (l. MHHNU 10896). Scale bars: 1 cm.
Diagnosis.
L. carminostipes exhibits a red-brown to orange-brown, translucent-striate pileus, salmon-pink adnate lamellae, a carmine equalstipe, globose to subglobose echinulate basidiospores, and a loosely interwoven pileipellis.
Microscopic features of Laccaria carminostipes (MHHNU 31553, holotype). a. Basidia; b. Basidiospores; c. Pileipellis; d. Stipitipellis. Scale bars: 10 μm.
Type.
China • Hunan Province: Sangzhi County, Badagongshan National Nature Reserve, at 29.65000°N, 109.695833°E, alt. 1742 m, in broad forest with trees of Fagaceae and bamboo, 29 July 2019, Z.H. Chen 31553 (MHHNU 31553, holotype).
Description.
Basidiomata small to medium-sized. Pileus 15–45 mm in diam, applanate to plano-concave, umbilicate, surface always fibrillose, sometimes missing, red-brown (7B6–7) to orange-brown (6A5–7), becoming darker with age, subhygrophanous; margin slightly translucent-striate. Lamellae adnate, distant, salmon-pink (7A3), narrow (about 2 mm high), entire, ventricose. Stipe 70–90 × 3–7 mm, central, equal, hollow, often slightly flexible, carmine (7C6–8); surface with indistinct to distinct whitish fibrillose coating; base with white (1A1) tomentum. Context thin, whitish (2A1) to brownish (5A2).
Basidiospores [100/4/4] (6.5) 7–9 (9.5) × (6) 6.5–8 (8.5) μm, Q = (0.88) 0.96–1.18 (1.33), Qm = 1.04 ± 0.08, mostly globose to subglobose, sometimes broadly ellipsoid, inamyloid, cyanophilous, echinulate; spines up to 1 μm long, ≤ 1 μm broad at base, distant; hilar appendix 1.0–1.8 μm long, prominent, truncate. Basidia 38–46 × 10–13 μm, clavate, mostly 4-spored, rarely 2-spored; sterigmata 6–8 μm long. Pleurocystidia and cheilocystidia not seen. Lamellar trama regular to subregular, composed of filamentous hyphae 3–6 μm wide. Pileipellis a cutis; hyphae loosely interwoven, thin-walled, cylindrical, 8–10 μm wide, in place with erect ends, with a brownish (3A2) intracellular pigment. Stipitipellis a cutis, composed of parallel to interwoven, thin-walled, cylindrical hyphae 3–8 μm wide, with some exserted inflated ends. Clamp connections present in all parts of basidiomata.
Ecology.
Always gregarious, under the trees of Fagaceae, in montane coniferous and broad-leaved mixed forests; summer (Jul.–Aug.).
Distribution.
Known from the subtropical zones of China.
Additional specimen examined.
China • Hunan Province, Sangzhi County, Badagongshan National Nature Reserve, at 29.65000°N, 109.695833°E, alt. 1740 m, 29 July 2019, Z.H. Chen 31552 (MHHNU 31552); • Sangzhi County, Badagongshan National Nature Reserve, at 29.755000°N, 109.762500°E, alt. 1592 m, under trees of Fagaceae,27 July 2020,Z.H. Chen 34706 (MHHNU 34706); • Yunnan Province, Jingdong Yi Autonomous County, Ailao Mountain, at 23.490278°N, 100.273333°E, alt. 2550 m, under trees of Quercus, 8 August 2024, P. Zhang 5444 (MHHNU 11944).
Notes.
Laccaria rubroalba and L. carminnostipes are characterized by medium-sized reddish basidiomata with a transluscent-striate pileus, but the former can be distinguished from the latter by having longer spines (1.2–2.7 μm long in L. rubroalba vs. up to 1 μm long in L. carminnostipes) and the presence of pleurocystidia and cheilocystidia (Luo et al. 2016). Laccaria cinnabarina J. Li & Y.Y. Cui is similar to L. carminnostipes by the reddish-brown stipes, but differs by its larger pileus (10–90 mm in L. cinnabarinavs. 15–45 mm in L. carminnostipes) and stronger spines (2 × 2 μm in L. cinnabarinavs. 1 × 1 μm in L. carminnostipes) (Li et al. 2024). Laccaria macrobasidia H.J. Cho & Y.W. Lim may sometimes be confused with L. carminnostipes, due to the similarity in size and color of their basidiomata. However, the two species can be differentiated based on the following diagnostic characteristics: basidiospores (9–11 × 8–10 μm in L. macrobasidia vs. 7–9 × 6.5–8 μm in L. carminnostipes), basidia (52–80 × 11–15 μm in L. macrobasidia vs. 38–46 × 10–13 μm in L. carminnostipes), pleurocystidia (present in L. macrobasidia, absent in L. carminnostipes), and the occurrence (L. macrobasidiain temperate forests, L. carminnostipes in subtropical forests) (Cho et al. 2020).
According to our phylogenetical analysis (Fig. 1), L. carminostipes (Species 38) could be most closely related to L. fagacicola (Species 37), L. darjeelingensis (Species 36), and L. aurantiaca (Species 35). Laccaria fagacicola can be distinguished from L. carminostipes by the presence of abundant cheilocystidia (Cui et al. 2021). Laccaria darjeelingensis differs by its dull red pileus and possesses both pleurocystidia and cheilocystidia (Thapa et al. 2024). Laccaria aurantiaca displays longer spines (0.7–1.8 μm vs. ≤ 1.0 μm in L. carminostipes) and produces pleurocystidia and cheilocystidia (Tang et al. 2025). Our phylogenetic analysis shows the placement of the sample GMM 6585 within the clade of L. carminostipes (Species 38, 99% BP, 1.00 PP, pairwise identity values of ITS = 99.84%), suggest that this specimen might be L. carminostipes.
Laccaria
mangshanensis
Taxon classificationFungiAgaricalesHydnangiaceae
Y.D. Xu & Z.M. He sp. nov.
A583755F-1E1D-5D26-B8FB-B2779C6CF47F
Fungal Names: FN 572507
Etymology.
‘mangshanensis’ referring to the locality of the holotype.
Microscopic features of Laccaria mangshanensis (MHHNU 8850, holotype). a. Basidia; b. Basidiospores; c. Pileipellis; d. Stipitipellis; e. Cheilocystidia. Scale bars: 10 μm.
Diagnosis.
Laccaria mangshanensis has a pale rosy, hemispherical, radially translucent-striate pileus, pinkish adnate lamellae, a reddish equal stipe, globose to subglobose echinulate basidiospores, and cheilocystidia.
Type.
China • Hunan Province: Yizhang County, Mangshan National Nature Reserve, at 24.866667°N,112.721944°E, alt. 1200 m, 28 July 2016, P. Zhang 2350 (MHHNU 8850, holotype).
Description.
Basidiomata small. Pileus 10–30 mm in diam, convex to hemispherical, centrally depressed, surface tomentose, dry, not hygrophanous, reddish-brown (7A5) to rosy (7A6) when young, then fading to pink-beige (7A2); margin strongly striate, rugulose-striate or rugulose-sulcate, involute to inflexed. Lamellae adnate, distant, pinkish (7A2) to white (1A1), narrow (about 2 mm high), entire, ventricose. Stipe 20–70 × 2–5 mm, central, equal, hollow, obviously flexible, reddish (7B5) to dull red (7C6), surface with distinct whitish fibrillose coating, base with white (1A1) tomentum. Context thin, whitish (2A1).
Basidiospores [100/4/2] (6.5) 7–8.5 (10) × (6) 6.5–8.5 (9) μm, Q = (0.86) 0.93–1.18 (1.31), Qm = 1.05 ± 0.08, mostly globose to subglobose, thin-walled, inamyloid, cyanophilous, hyaline, echinulate, spines (0.5) 1–1.2 (1.5) μm long, 0.3–0.8 (1) in width, subdistant; hilar appendix 0.8–1.5 μm long, subtruncate. Basidia 38–47 × 10–13 μm, 4-spored, clavate, sharply narrowed, thin-walled, hyaline; sterigmata up to 9 μm long. Pleurocystidia lacking. Cheilocystidia 25–55 × 3–5 μm, filamentous to narrowly clavate, flexuose, thin-walled, hyphae, abundant. Lamellar trama regular to subregular; hyphae cylindrical, hyaline, thin- walled, 3–8 μm wide. Pileipellis a cutis, composed of thin-walled, interwoven, cylindrical hyphae 4–10 μm wide, with exserted ends, hyphae. Stipitipellis a cutis, composed of appressed, parallel, thin- to slightly thick-walled (ca. 0.5 μm) hyphae. Clamp connections present in all parts of basidiomata.
Ecology.
Single, in clusters or in groups, on soil, under the trees of Fagaceae, in subtropical montane coniferous and broadleaved mixed forest, summer (Jul.).
Distribution.
Known from Central China.
Additional specimen examined.
China • Hunan Province: Yizhang County, Mangshan National Nature Reserve, at 24.869722°N, 112.721944°E, alt. 1200 m, 29 July 2016, P. Zhang 2356 (MHHNU 8856).
Notes.
Laccaria fengkaiensis F. Liand L. mangshanensis share pale red to pastel red basidiomata, but the former can be distinguished from the latter by the larger pileus (50–90 mm pileus width vs. 10–30 mm in L. mangshanensis), smaller basidiospores (5–6 × 5–6 μm vs. 7–8.5 × 6.5–8.5 μm in L. mangshanensis), and the presence of pileocystidia (Li 2020). However, the phylogenetic analysis (Fig. 1) suggests that L. mangshanensis represents a well-supported clade with strong support (Species 41, 94% BP, 0.94 PP), and is clearly separated from L. fengkaiensis (Species 65).
Laccaria
sinolateritia
Taxon classificationFungiAgaricalesHydnangiaceae
Y.D. Xu & Z.M. He sp. nov.
7DE42960-73A2-5B48-8C29-C5CB0A941462
Fungal Names: FN 572508
Etymology.
‘sino’ referring to China, ‘lateritia’ (Latin) referring to its red to brownish orange fruiting body.
Microscopic features of Laccaria sinolateritia (MHHNU 11956, holotype). a. Basidia; b. Basidiospores; c. Pileipellis; d. Stipitipellis. Scale bars: 10 μm.
Basidiospores of described species under SEM. a, b. Laccaria carminostipes (MHHNU 31553, holotype); c, d. L. mangshanensis (MHHNU 8850, holotype); e, f. L. sinolateritia (MHHNU 11956, holotype). Scale bars: 2 μm.
Diagnosis.
Laccaria sinolateritiais characterized by brownish red basidiomata, globose to subglobose echinulate basidiospores, and the presence of pileocystidia and caulocystidia.
Type.
China • Yunnan Province: Jingdong Yi Autonomous County, Ailaoshan, at 24.904444°N,101.059444°E, alt. 2500 m, 8 August 2024, P. Zhang 5456 (MHHNU 11956, holotype).
Description.
Basidiomata small to medium-sized. Pileus 10–40 mm in diam, at first convex, then plano-concave to concave; surface tomentose, brownish red (6B4), subhygrophanous; translucent-striate or rugulose-sulcate, straight, undulate with age. Lamellae distant, entire, adnate, broad (about 5 mm high), ventricose, concolorous with pileus surface. Stipe 35–50 × 1–3 mm, brownish red (6B4), equal, broadly fistulose; surface covered with whitish (2A1) longitudinal fibrils; base with a whitish (2A1) mycelium. Context brownish (6A4).
Basidiospores [100/4/2] (7) 7.5–10 (10.5) × (7) 7.5–10 (10.5) μm, Q = (0.90) 0.91–1.07 (1.11), Qm = 1.00 ± 0.05, mostly globose, hyaline, echinulate; spines 2–2.5 (3) μm long, ca. (1) 1.5–2 μm wide at base, crowded; hilar appendix 1.3–2.5 long, prominent, subtruncate. Basidia 40–58 × 12–14 μm, with 4 sterigmata up to 12 μm long, hyaline, subcylindrical to clavate. Pleurocystidia and cheilocystidia not seen. Lamellar trama subregular to interwoven; hyphae cylindrical, hyaline, thin-walled, 2–6 (7) μm wide. Pileipellis a cutis, hyphae cylindrical, 8–10 (12) μm in diameter with some hyphal ends, subregular to interwoven. Pileocystidia abundant, 25–45 × 5–15 μm, cylindrical to clavate, vertically to subvertically arrange, scattered to aggregating into clusters on the pileus surface. Stipitipellisa cutis, composed of cylindrical hyphae 5–10 (11.5) μm wide with abundant caulocystidia, thin- to thick-walled (≤0.5 μm). Caulocystidia 37.5–55 (67.5) × 7.5–9.5 (10) μm, clavate, scattered to aggregating into clusters on the stipitipellis surface. Clamp connections present in all parts of basidiomata.
Ecology.
Gregarious, under the trees of Fagaceae, in subtropical montane forests, summer (Aug.).
Distribution.
Known from the subtropical zones of China.
Additional specimen examined.
China • Yunnan Province: Jingdong Yi Autonomous County, Ailaoshan, at 24.904444°N, 101.059444°E, alt. 2500 m, 8 August 2024, P. Zhang 5458 (MHHNU 11958).
Notes.
Macroscopically, L. lateritia Malençon and L. sinolateritia are easily confused. For instance, they share similar-sized basidiomata, reddish-brown pileus and similar-sized basidiospores (7.5–10.5 × 7.5–10.5 µm in L. lateritia vs. 7.5–10 × 7.5–10 µm in L. sinolateritia). However, L. lateritia can be distinguished by the production of 2-spored basidia, shorter spines (±1 μm high vs. 2–2.5 μm high in L. sinolateritia) and the presence of cheilocystidia (Cooper 2015). Additionally, L. darjeelingensis resembles L. sinolateritia in its red basidiomata, but differs in having smaller basidiospores (5.9–7.6 × 5.9–7.6 µm vs. 7.5–10 × 7.5–10 µm in L. sinolateritia) and abundant flexuous pleurocystidia and cheilocystidia (Thapa et al. 2024).
According to the phylogenetical analysis (Fig. 1), L. subroseoalbescens (species 68) and L. infundibuliformis (species 69) are related to L. sinolateritia (species 67), and form a clade with low support (54%BP, 0.91PP). Laccaria subroseoalbescensis characterized by a yellow pileus, the presence of cheilocystidia and pleurocystidia, and the absence of pileocystidia (Tang et al. 2024). Laccaria infundibuliformis differs in its smaller pileus (8–27 mm in diam vs. 10–40 mm in diam in L. sinolateritia), smaller basidia (29–50 × 6–9 µm vs. 40–58 × 12–14 μm in L. sinolateritia), and the absence of pileocystidia (Thapa et al. 2024).The four-locus phylogenetical analysis also supports the identification of the four Chinese samplesKUN-HKAS83382, GMM6776, T168, and T107 as L. sinolateritia (pairwise identity values of ITS = 99.39%–100%).
Discussion
Our study presents three new species of Laccaria, viz. L. carminostipes, L. mangshanensis and L. sinolateritia based on a comprehensive analysis of morphological and molecular phylogenetic data (Aime et al. 2021). The four-locus phylogenetic tree strongly supports that they are monophyletic clades separated from other Laccaria species (Fig. 1). In addition, L. japonica and L. versiforma is formally reported here for the first time in China.
Laccaria carminostipes, L. mangshanensis, and L. sinolateritia were collected from Yunnan and Hunan Provinces, regions known for their rich ecological resources and high fungal diversity. Laccaria carminostipesis can be distinguished from other Laccaria species by its distinctive carmine-colored stipe. L. mangshanensis is characterized by a rosy pileus and the presence of abundant cheilocystidia. And L. sinolateritia is characterized by its brownish-red basidiomata and the presence of abundant pileocystidia and caulocystidia.
Regarding L. japonica (Vincenot et al. 2017), the Chinese specimens, MHHNU 9589, 34710 and 34711 (Fig. 2j, k), resemble to this species, exhibiting small to medium-sized, bright purple basidiomata, adnate lamellae, and similar basidiospores size (8–10 × 8–10 µm in our specimens, 9–10 × 7–9 µm in L. japonica). Our phylogenetic analysis (Fig. 1) further indicates that these samples form a well-supported clade together with the holotype of L. japonica (90% BP, 1.00 PP, pairwise identity values of ITS = 99.68%–99.83%). Similarly, the Chinese specimen MHHNU 10896 (Fig. 2l) has small to medium-sized basidiomata, featuring pale brown to pinkish brown pileus, brown stipes, pinkish lamellae, and echinulate basidiospores measuring 7–10 × 7–9.5 µm. These features are entirely consistent with the description of L. versiforma provided by Cho et al. (2018). Furthermore, in the four-locus phylogenetic tree (Fig. 1), the specimen MHHNU 10896 clusters together with four Korean specimens of L. versiforma with strong support (99% BP, 1.00 PP). The ITS sequence of MHHNU 10896 exhibits a similarity of 98.12%–99.64%. Both L. japonica and L. versiforma occur in temperate or subtropical forest. Laccaria japonicawas previously documented in temperate forests of Salix reinii Franch. & Sav.in Japan (Vincenot et al. 2017), while our Chinese collections of L. japonica were found in subtropical forests of Fagaceae. In Korea, L. versiforma was found in temperate forests of Quercus and Pinus densiflora Siebold &Zucc. (Cho et al. 2018), while our specimen of L. versiforma was found in subtropical forests of Quercus.
The phylogram recovers 87 species of Laccaria, yet the infrageneric classifications for some species within this genus remains indeterminate. Our specimen MHHNU 12061 could be L. acanthospora, based on its orange pileus, and basidiospores measuring 8–10 × 7–10 µm (7–10 × 7–10 µm in L. acanthospora, Wilson et al. 2013). However, the sample MHHNU 12061 exhibits a relatively long genetic distance from the holotype of L. acanthospora (AWW485, Fig. 1) and possesses a larger pileus measuring 20–30 mm in width (Suppl. material 2: fig. S1l, 4–15 mm in Wilson et al. 2013). Because we have only one specimen, it remains uncertain whether these morphological differences reflect intraspecific variation or are significant enough to warrant the recognition of a distant species.
Laccaria bicolor was originally described from Europe (Maire 1937). According to our phylogenetic analysis (Fig. 1), the sequences named L. bicolor were clustered into four clades (Species 3, 9, 14 and 15). Due to the lack of the sequence of the type specimen for L. bicolor, the exact phylogenetic position of this species remains uncertain. Our Chinese specimen MHHNU 11595 (Suppl. material 2: fig. S1e) clusters together with a French specimen of L. bicolor with full support in our phylogenetic analysis (Fig. 1). Since their ITS pairwise identity value is 97.91%, the Chinese specimen could be a cryptic species. Laccaria guizhouensis (Zhang et al. 2024) and L. neovinaceoavellanea (Zhang et al. 2023) are likely conspecific, as their holotypes (HMAS 352265 and GDGM52852) form a highly supported clade (98% BP, 1.00 PP) with an ITS pairwise identity value of 99.44%. Similarly, L. himalayensis and L. bullipellis may also be conspecific, given their high ITS pairwise identity values of 99.17% and stable clustering with a full support.
Based on the four-locus phylogenetic and morphological analyses (Fig. 1, Suppl. material 2: fig. S1), we also identified nine other previously described species: L. alba, L. araneosa, L. aurantia, L. brunnea, L. fagacicola, L. moshuijun, L. murina, L. rubroalba, and L. stipalba. Both L. alba and L. aurantina, were first reported in Yunnan Province. In this study, L. alba was also found in Hunan and Guizhou Provinces, and L. aurantina was discovered in Hubei and Guizhou Provinces. Laccaria araneosa, originally described in Korean temperate forests (Cho et al. 2018), and later also reported in Jilin Province, temperate China (Wang et al. 2022). We also found this species in Hunan Province, subtropical China.
Laccaria brunnea was first recorded in Yunnan Province, China. This species is characterized by its brownish basidioma with salmon lamellae, and the presence of pleurocystidia and cheilocystidia (Tang et al. 2025). Our specimen MHHNU 9692 (Suppl. material 2: fig. S1d), collected from Guizhou Province, China, also has a gray to brownish pileus, salmon lamellae, and globose to subglobose, echinulate basidiospores, showing morphological resemblance to L. brunnea. In our four-locus phylogenetic tree (Fig. 1), the samples MHHNU 9692, T44, and T110 cluster together with the holotype (KUN-HKAS 123286) of L. brunnea (100% BP, 1.00 PP, pairwise identity values of ITS = 98.37%–98.91%), confirming the identification of sample MHHNU 9692 as L. brunnea. Notably, both pleurocystidia and cheilocystidia were found to be absent in our specimen MHHNU 9692.Laccaria stipalbais characterized by its dark orange to grayish pink pileus, distinctive white stipe, and globose echinulate basidiospores, and was originally described in Yunnan Province (Tang et al. 2025). During 2022 to 2024, we also collected three specimens MHHNU 11314, 11324 and 12057 from Yunnan Province, China, which exhibit the same microfeatures (Suppl. material 2: fig. S1j, k and l). In our phylogenetic analysis (Fig. 1), these three samples cluster with the holotype of L. stipalba (KUN-HKAS 123285) with full support (pairwise identity values of ITS = 99.65%–99.83%), confirming that these samples should be identified as L. stipalba. It should be pointed out that the basidiospores size of our specimens is slightly larger (7–10 × 7–10 μm vs. 5.8–8.4 × 5.5–8.1 μm in Tang et al. 2025), and pleurocystidia and cheilocystidia were not found in our examination.
Supplementary Material
XML Treatment for Laccaria carminostipes
XML Treatment for Laccaria mangshanensis
XML Treatment for Laccaria sinolateritia
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aime MC Miller AN Aoki T Bensch K Cai L Crous PW Hawksworth DL Hyde KD Kirk PMLücking R May TW Malosso E Redhead SA Rossman AY Stadler M Thines M Yurkov AM Zhang N Schoch CL (2021) How to publish a new fungal species, or name, version 3.0. IMA Fungus 12: 11. 10.1186/s 43008-021-00063-1PMC 809150033934723 · doi ↗ · pubmed ↗
- 2Berkeley MJ Broome CE (1883) Notices of British fungi> (1989–2027).Annals & Magazine of Natural History 12: 370–374. 10.1080/00222938309459647 · doi ↗
- 3Besson MKühner R (1971) Ultrastructure de la paroisporique des Laccaria Berk. et Br. (Agaricales).Comptes Rendus Hebdomadaires des Séances de l’Académie des Sciences 272: 1078–1081.
- 4Campi M Mancuello C Maubet Y Niveiro N (2017) Laccaria fraterna (Cooke & Mass.: Sacc.) Pegler, 1965 (Agaricales, Basidiomycota) associated with exotic Eucalyptus sp. in northern Argentina and Paraguay.Check List 13: 87–90. 10.15560/13.4.87 · doi ↗
- 5Castresana J (2000) Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis.Molecular Biology and Evolution 17: 540–552. 10.1093/oxfordjournals.molbev.a 02633410742046 · doi ↗ · pubmed ↗
- 6Cho HJ Park MS Lee H Oh SY Wilson AW Mueller GM Lim YW (2018) A systematic revision of the ectomycorrhizal genus Laccaria from Korea.Mycologia 110: 948–961. 10.1080/00275514.2018.150754230240340 · doi ↗ · pubmed ↗
- 7Cho HJ Lee H Park MS Park KH Park JH Cho Y Kim C Lim YW (2020) Two new species of Laccaria (Agaricales, Basidiomycota) from Korea.Mycobiology 48: 288–295. 10.1080/12298093.2020.178696132952411 PMC 7476507 · doi ↗ · pubmed ↗
- 8Cooper J (2015) Mycological Notes – 30: A Preliminary Key to New Zealand species of Laccaria. Manaaki Whenua – Landcare Research. [Unpublished technical report]