Pre- and postnatal maternal depressive symptoms associated with local connectivity of the left amygdala in 5-year-olds
Elena Vartiainen, Anni Copeland, Elmo P. Pulli, Venla Kumpulainen, Eero Silver, Olli Rajasilta, Ashmeet Jolly, Silja Luotonen, Hilyatushalihah K. Audah, Niloofar Hashempour, Wajiha Bano, Ilkka Suuronen, Ekaterina Saukko, Suvi Häkkinen, Hasse Karlsson, Linnea Karlsson

TL;DR
Maternal depressive symptoms after birth are linked to changes in the brain's left amygdala in 5-year-old children.
Contribution
This study identifies a novel postnatal pathway linking maternal depression to offspring neurodevelopment.
Findings
Maternal depressive symptoms at 3 months postnatal correlate with increased local brain activity in the left amygdala.
The association is localized and strongest for early postnatal maternal depressive symptoms.
No significant associations were found with distal brain connectivity of the left amygdala.
Abstract
Maternal depressive symptoms can influence brain development in offspring, prenatally through intrauterine programming, and postnatally through caregiving related mother–child interaction. The participants were 5-year-old mother–child dyads from the FinnBrain Birth Cohort Study (N = 68; 28 boys, 40 girls). Maternal depressive symptoms were assessed with the Edinburgh Postnatal Depression Scale (EPDS) at gestational week 24, 3 months, 6 months, and 12 months postnatal. Children’s brain imaging data were acquired with task-free functional magnetic resonance imaging (fMRI) at the age of 5 years in 7-min scans while watching the Inscapes movie. The derived brain metrics included whole-brain regional homogeneity (ReHo) and seed-based connectivity maps of the bilateral amygdalae. We found that maternal depressive symptoms were positively associated with ReHo values of the left amygdala. The…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
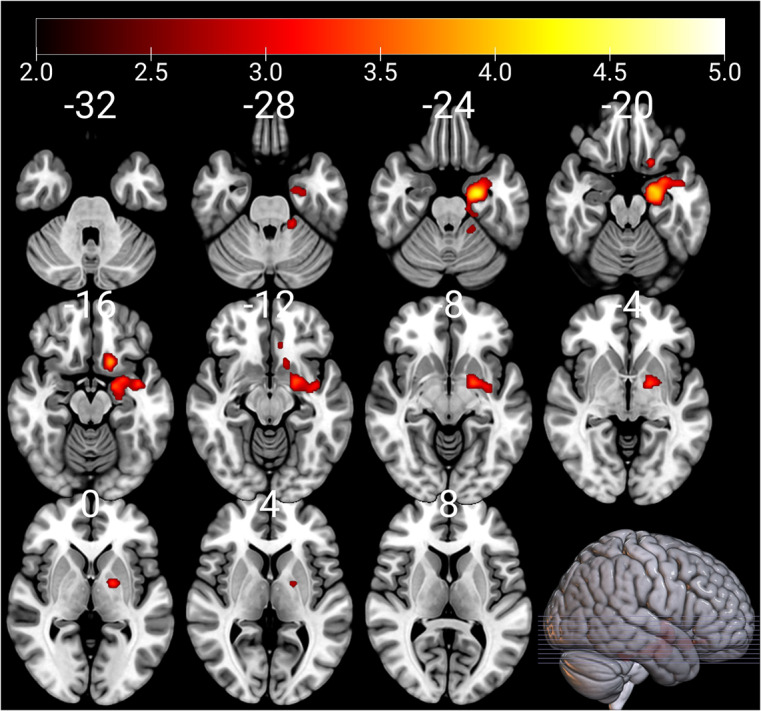 Figure 1
Figure 1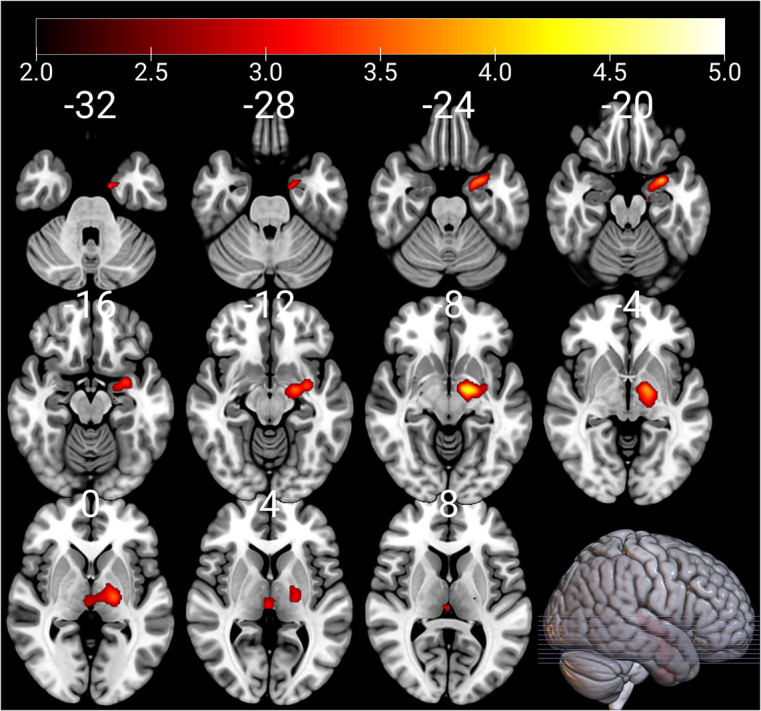 Figure 2
Figure 2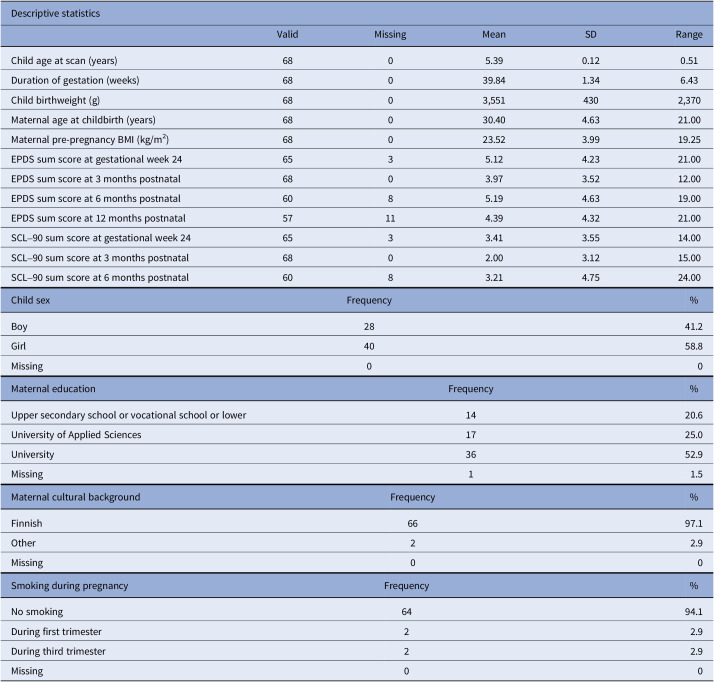 Figure 3
Figure 3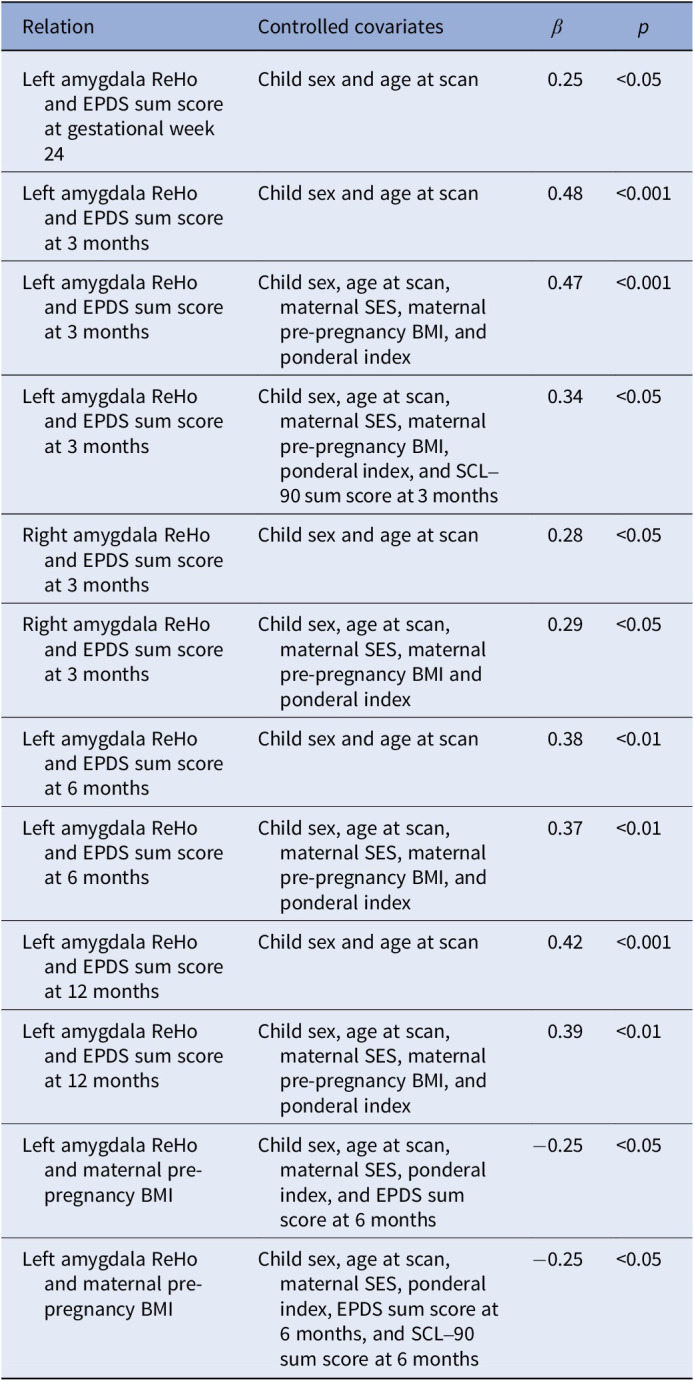 Figure 4
Figure 4- —Sigrid Juséliuksen Säätiöhttps://doi.org/10.13039/501100006306
- —Emil Aaltosen Säätiöhttps://doi.org/10.13039/501100004756
- —Suomen Lääketieteen Säätiöhttps://doi.org/10.13039/100008723
- —Alfred Kordelinin Säätiöhttps://doi.org/10.13039/100008969
- —Juho Vainion Säätiöhttps://doi.org/10.13039/501100004037
- —Turun Yliopistosäätiöhttps://doi.org/10.13039/501100022793
- —Varsinais-Suomen Sairaanhoitopiirihttps://doi.org/10.13039/501100009420
- —State Grants for Clinical Research (ERVA)
- —Orionin Tutkimussäätiöhttps://doi.org/10.13039/501100007083
- —Signe ja Ane Gyllenbergin Säätiöhttps://doi.org/10.13039/501100004325
- —Strategic Research Councilhttps://doi.org/10.13039/501100009047
- —Päivikki ja Sakari Sohlbergin Säätiöhttps://doi.org/10.13039/501100004212
- —Juho Vainion Säätiöhttps://doi.org/10.13039/501100004037
- —Emil Aaltosen Säätiöhttps://doi.org/10.13039/501100004756
- —Suomen Aivosäätiöhttps://doi.org/10.13039/501100008320
- —Turun Yliopistosäätiöhttps://doi.org/10.13039/501100022793
- —Suomen Kulttuurirahastohttps://doi.org/10.13039/501100003125
- —Suomen Kulttuurirahastohttps://doi.org/10.13039/501100003125
- —Emil Aaltosen Säätiöhttps://doi.org/10.13039/501100004756
- —Signe ja Ane Gyllenbergin Säätiöhttps://doi.org/10.13039/501100004325
- —Turun yliopiston tutkijakouluhttps://doi.org/10.13039/501100019391
- —Turun yliopiston tutkijakouluhttps://doi.org/10.13039/501100019391
- —Emil Aaltosen Säätiöhttps://doi.org/10.13039/501100004756
- —Turun Yliopistosäätiöhttps://doi.org/10.13039/501100022793
- —Research Council of Finland
- —Strategic Research Council (SRC) established within the Research Council of Finland
- —Signe ja Ane Gyllenbergin Säätiöhttps://doi.org/10.13039/501100004325
- —Finnish State Grants for Clinical Research
- —Jane ja Aatos Erkon Säätiöhttps://doi.org/10.13039/501100004012
- —Echnerska Frilasarettet Foundation
- —Signe ja Ane Gyllenbergin Säätiöhttps://doi.org/10.13039/501100004325
- —State Research Grant
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMaternal Mental Health During Pregnancy and Postpartum · Cognitive Abilities and Testing · Cardiovascular Issues in Pregnancy
Introduction
Maternal psychological distress in pregnancy can affect fetal brain development in fundamental ways [1, 2]. The prevalence of clinically relevant level of depressive symptoms, including major and minor depressive disorder, in pregnancy is around 11% in the first trimester and 8.5% in the second and third trimesters [3]. Neuroimaging studies using structural and diffusion MRI studies have indicated that maternal prenatal stress, including depression, is associated with changes in child brain morphology [2, 4, 5]. These findings affirm that the prenatal period is critical for fetal neurodevelopment and poor maternal well-being during pregnancy may have long-lasting effects on the offspring. Effects of prenatal depressive symptoms are found in brain structures that are responsible for emotional regulation, cognition, memory, and decision making – with a majority of the studies implicating the amygdala [6].
Resting state functional magnetic resonance imaging (rs-fMRI) can provide novel information on maternal distress as a predictor of offspring’s neurodevelopment [7–10]. An rs-fMRI study on infant neurodevelopment showed that maternal prenatal depressive symptoms were associated with increased functional connectivity between medial prefrontal cortex (mPFC) and left amygdala in 6-month-old infants [9]. Rajasilta et al. demonstrated that in infants approximately 1 month of age maternal prenatal distress was positively related to rs-fMRI-derived fractional amplitude of low-frequency fluctuations (fALFF) in mPFC, which indicates that prenatal stress may affect functional features of the maturing brain during gestation [11]. Children of mothers who experienced psychological distress late in their pregnancy had increased amygdala functional connectivity with the ventromedial PFC and anterior insula [12]. In comparison to prenatal maternal depressive symptoms, early postnatal maternal well-being and later child development have received significantly less attention in neuroimaging studies, even though the first years of life are crucial time for parent–child bonding and child development.
To the best of our knowledge, there is only one prior functional neuroimaging study linking both pre- and postnatal maternal depressive symptoms to adverse child brain connectivity outcomes in early childhood. This study by Soe et al. [13] investigated the relationship between perinatal maternal depressive symptoms and bilateral amygdala in 4.5-year-old children. They found significant positive associations between maternal prenatal depressive symptoms and the left amygdala functional connectivity with the right insula and putamen, bilateral subgenual anterior cingulate cortex (ACC), and caudate. Similar associations were found between the right amygdala and left orbitofrontal cortex and temporal pole. Greater pre- than postnatal depressive symptoms had associations with lower functional connectivity of the left amygdala with the bilateral subgenual ACC and left caudate. Correspondingly, greater prenatal maternal depressive symptoms were associated with lower functional connectivity of the right amygdala with the left orbitofrontal cortex, insula, and caudate. These findings were gender specific to girls. However, the study did not find significant interactions of gender with pre- or postnatal maternal depressive symptoms on amygdala functional connectivity [13].
In this study, we investigate whether maternal pre- and postnatal depressive symptoms associate with 5-year-olds’ local connectivity across the whole brain as assessed by regional homogeneity (ReHo), a measure derived from task-free fMRI while the participants were watching the Inscapes movie [14]. Second, replicating the analyses in prior work [13], we mapped the association of maternal depressive symptoms to bilateral amygdala seed-based connectivity. Third, multiple regression analyses were performed to assess the associations between maternal depressive symptoms and bilateral amygdala ReHo values. Our prior study has found that early emotional availability of the mother associated with ReHo values in the subset of the same neuroimaging data [15], which encouraged us to use ReHo as the main derived brain measure for this study as well. This was an exploratory study and thus no explicit hypotheses were placed regarding the direction or strength of the associations.
Methods
This study was conducted in accordance with the Declaration of Helsinki, and it was approved by the Joint Ethics Committee of the University of Turku and the Hospital District of Southwest Finland (March 15, 2011) §95, ETMK: 31/180/2011. Written informed consent was obtained from the participants, and parents gave consent on behalf of their children. We followed the Strengthening the Reporting of Observational studies in Epidemiology (STROBE) reporting guidelines for cohort studies (https://www.strobe-statement.org). The participant criteria, MRI scanning visits, image acquisition, and preprocessing are identical to those used in our prior work [15].
Participants
The participants are part of the FinnBrain Birth Cohort Study, which prospectively examines the influence of genetic and environmental factors on child development and later mental and physical health outcomes [16]. Pregnant women attending their first trimester ultrasound were recruited in maternal welfare clinics in the Turku region of the Southwest Finland Hospital District and the Åland Islands between December 2011 and April 2015. Ultrasound-verified pregnancy and a sufficient knowledge of Finnish or Swedish were required for participation.
The exclusion criteria for the neuroimaging study were: (1) born before gestational week 35 (week 32 for those with exposure to maternal prenatal synthetic glucocorticoid treatment), (2) developmental or major organ abnormalities in senses or communication (e.g., blindness, deafness, congenital heart disease), (3) known long-term medical diagnosis (e.g., epilepsy, autism), (4) ongoing medical examinations or clinical follow-up in a hospital, (5) the child using continuous, daily medication (including oral medications, topical creams, and inhalants; desmopressin was allowed), (6) history of head trauma (defined as concussion necessitating clinical follow-up in a healthcare setting), (7) metallic ear tubes, and (8) routine MRI contraindications.
In total, 203 children participated in a neuroimaging visit at 5 years of age, and 77 of them had successful functional scans due to limited subject compliance (the fMRI data were acquired last), of which 68 had the maternal Edinburgh Postnatal Depression Scale (EPDS) questionnaires collected, and were included in the study after quality control steps outlined later. Participant characteristics are reported in Table 1.Table 1.Demographics of the study participantsDescriptive statisticsValidMissingMeanSDRangeChild age at scan (years)6805.390.120.51Duration of gestation (weeks)68039.841.346.43Child birthweight (g)6803,5514302,370Maternal age at childbirth (years)68030.404.6321.00Maternal pre-pregnancy BMI (kg/m^2^)68023.523.9919.25EPDS sum score at gestational week 246535.124.2321.00EPDS sum score at 3 months postnatal6803.973.5212.00EPDS sum score at 6 months postnatal6085.194.6319.00EPDS sum score at 12 months postnatal57114.394.3221.00SCL–90 sum score at gestational week 246533.413.5514.00SCL–90 sum score at 3 months postnatal6802.003.1215.00SCL–90 sum score at 6 months postnatal6083.214.7524.00Child sexFrequency%Boy2841.2Girl4058.8Missing00Maternal educationFrequency%Upper secondary school or vocational school or lower1420.6University of Applied Sciences1725.0University3652.9Missing11.5Maternal cultural backgroundFrequency%Finnish6697.1Other22.9Missing00Smoking during pregnancyFrequency%No smoking6494.1During first trimester22.9During third trimester22.9Missing00Abbreviations: BMI, body mass index; EPDS, Edinburgh Postnatal Depression Scale; SCL-90, Symptom Checklist 90; SD, standard deviation.
Maternal depressive and anxiety symptoms and perinatal data
Questionnaires assessing maternal psychological health were filled in by the mothers in gestational week 24 and 3 months, 6 months, and 12 months postnatal. We assessed prenatal depressive symptoms at gestational week 24 because at this time point neurogenesis takes place, cortex begins to fold, and myelin starts to develop [17–19]. Maternal depressive symptoms were assessed with the EPDS [20], which has been validated for use during pregnancy. This 10-item questionnaire is scaled from 0 to 30 points with a higher score denoting increased symptom severity. A score of 10 has been implicated as a clinically meaningful threshold for symptoms of depression in pregnancy [21]. Pre- and postnatal maternal anxiety symptoms were evaluated using the anxiety scale of Symptom Checklist 90 (SCL-90) [22] at gestational week 24, 3 months and 6 months postnatal. The anxiety subscale of SCL-90 is a reliable and valid measure of anxiety symptoms in both clinical and research settings and the questionnaire consists of 10 items scaled from 0 to 4. Obstetric data were retrieved from the Finnish National Birth Register (National Institute for Health and Welfare, www.thl.fi). Other information was gathered using questionnaires on behalf FinnBrain Birth Cohort Study.
Magnetic resonance imaging visits
All MRI scans were performed for research purposes, and participants were scanned awake and without sedation. The imaging was performed at the Department of Radiology, Turku University Hospital between October 2017 and March 2021. The practical arrangements to perform child-friendly visits, more details of the visit have been described previously [15]. Anatomical images were screened by an experienced neuroradiologist for incidental findings. None of the participants included in this study had clinically relevant incidental findings.
Image acquisition
The MRI scans were conducted on a Siemens Magnetom Skyra fit 3T scanner (Siemens Medical Solutions, Erlangen, Germany). A 20-element Head/Neck Matrix coil allowed the use of the generalized autocalibrating partially parallel acquisition (GRAPPA) technique to accelerate acquisitions (parallel acquisition technique [PAT] factor of 2). The scans included a high-resolution T1 magnetization prepared rapid gradient echo (MPRAGE), a T2 turbo-spin echo (TSE), diffusion tensor imaging (DTI), and a 7-min fMRI. The fMRI consisted of 170 volumes with voxel size 3.0 × 3.0 × 3.0 mm^3^, TR 2500 ms, TE 30.0 ms, flip angle of 80°, and 42 axial slices without gaps. Full cerebellar coverage was not possible in all participants. Prior to fMRI acquisition, all children had rested by watching a movie or a TV show of their choice during the 30–45 min required for structural scanning. If the child had fallen asleep, they were gently awakened. During the fMRI sequence, participants were instructed to stay as still as possible with their eyes open. To minimize motion and reduce cognitive load, the Inscapes movie was played during fMRI data collection [14]. Visual stimuli were presented on an MRI-compatible 32″ LCD monitor with full-HD resolution (Nordic Neuro Lab) located at the foot of the bed of the scanner, which participants could watch via mirrors mounted on the head coil. The total scanning time was limited to 1 h, and the imaging was discontinued if the child expressed unwillingness to continue at any point.
Image preprocessing and estimating ReHo
Functional MRI data were slice-timing corrected and motion corrected in the FMRIB Software Library (v6.00, FSL; [23]) v6.00 relative to a manually chosen reference volume, free of major artifacts. Motion outliers were estimated using artifact detection tools (ART) (https://www.nitrc.org/projects/artifact_detect). We tagged images as outliers if they had composite motion threshold >2 mm or DVARS >9, which are default parameters in the ART toolbox. All children included in the final statistical analyses had a full fMRI sequence of 170 volumes, and a maximum of 50 volumes were tagged as outliers by ART. The descriptive statistics for motion were as follows (of full sample N = 68): motion outliers (mean 15, range 0–49), mean absolute displacement (mean 0.73, range 0.06–3.71, mm), and mean relative displacement (mean 0.25, range 0.02–1.36, mm). Anatomical masks for white matter and cerebrospinal fluid were defined in the Montreal Neurological Institute (MNI) standard space and spatially normalized using FSL, then registered to functional data with an affine transformation. Average signal in white matter and cerebrospinal fluid as well as 24 motion covariates (the six realignment parameters and their temporal derivatives and quadratic terms) were included as nuisance covariates. Taken together, denoising consisted of outlier rejection, nuisance regression, detrending, and high-pass filtering (0.008 Hz).
Our main imaging derivative was ReHo, a data-driven measure of local voxel-wise connectivity. ReHo describes synchronization between time series of a given voxel in its neighbors based on Kendall’s coefficient of concordance and is interpreted as a sign of synchronized activation or deactivation in blood–oxygen-level-dependent time series [24]. ReHo was computed as implemented in DPABI (number of voxels in a cluster; N = 27). For group analysis, ReHo maps were normalized nonlinearly to 1.0 × 1.0 × 1.0 mm^3^ MNI space using FSL FNIRT. Finally, the data were smoothed with a Gaussian filter of 6 mm full width at half maximum (FWHM).
Seed-based connectivity analysis (SCA)
The primary analyses focused on whole-brain ReHo maps. Our second goal was to replicate prior work [13] by studying the association of maternal depressive symptoms to bilateral amygdala seed-based connectivity. Additionally, in line with our prior work [15, 25], we had predefined plans to conduct complementary SCAs guided by the ReHo results. The SCAs were performed with FSL tools using the same preprocessing and nuisance regression as for the ReHo analyses except that the interquartile range (obtained via fsl_motion_outliers) of DVARS was used for the removal of motion corrupted volumes after confirming that it matched the ReHo pipeline described earlier.
Statistical analysis
Whole-brain voxel-wise statistical analyses for the ReHo maps and the SCA connectivity maps were performed with SPM12 software (https://www.fil.ion.ucl.ac.uk/spm/software/spm12/) with general linear models (GLM) and “multiple regression” design for ReHo. Association between maternal perinatal depressive symptoms and ReHo of the bilateral amygdala was assessed in two stages. GLM models were performed for EPDS scores separately for each of the time points (prenatally at gestational week 24 as well as 3 months, 6 months, and 12 months postnatal). First, child age at scan and sex were set as the independent variables (IVs) of no interest and EPDS score was set as the main explanatory variable (EV). Second, we added IVs of no interest including maternal socioeconomic status (SES) measured by maternal educational level, maternal pre-pregnancy BMI, child’s ponderal index (measured at MRI scan, relationship between body mass and height; mass/height^3^) [1], and maternal anxiety symptom score (SCL-90 score). Additionally, in analyses using postnatal EPDS scores, EPDS at gestational week 24 was added as an IV of no interest, to account for the possible continuum of depressive symptom levels from prenatal to postnatal period. Prior to statistical testing, we used an inclusive binary gray matter mask. The a priori statistical threshold for clusters was tested at p < 0.005 and corrected with family-wise error (FWE) at cluster level at p < 0.0125 (taking into account four separate statistical models for the EPDS time points).
Complementary region-of-interest-based linear regression models were performed for bilateral amygdala mean ReHo values with RStudio (R Core Team [2024]. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/). This analysis was carried out to describe the effect sizes of the associations between the ReHo of the amygdala, the independent variables in line with the SPM12 models, and with other additional independent variables described later. The ReHo values were obtained by creating binary masks of bilateral amygdala from the AAL atlas, using them to mask normalized ReHo maps and estimating mean with fslmaths.
All regression models were performed for left and right amygdala mean ReHo values separately. In the first regression model, IVs were set as EPDS score (gestational week 24, 3 months, 6 months, and 12 months postnatal separately), child sex, and child age at scan. In the second model, IVs were set as EPDS score (gestational week 24, 3 months, 6 months, and 12 months postnatal separately), child sex, child age at scan, maternal SES, maternal pre-pregnancy BMI, and ponderal index. Finally, we included maternal anxiety symptoms (SCL-90 scores at gestational week 24, 3 months, and 6 months) in the second model to test if the association was specific to depressive symptoms. Missing EPDS score, SCL-90 score, and maternal educational level values were mean imputed for statistical analyses. We checked that the regression models met basic assumptions: multicollinearity through variation inflation factor (all variance inflation factors, VIFs < 2.3) and that the residuals were normally distributed through visual inspection of Q–Q plots and histograms of the residuals.
We report uncorrected p values throughout the manuscript, but note that we performed eight statistical tests for linear regression models, 2 (left and right amygdala) × 4 (EPDS questionnaire time points), and the Bonferroni-corrected p-values are p < 0.00625. We created complementary scatterplots in line with the linear regression models, which are included in Supplementary Figures S1–S7.
Results
The sample characteristics are reported in Table 1. The sample included 28 boys and 40 girls. Maternal EPDS scores showed variability across different measurement points so that the mean scores were 5.12 (gestational week 24), 3.97 (3 months postnatal), 5.19 (6 months postnatal), and 4.39 (12 months postnatal). The corresponding SCL-90 scores were 3.41 (gestational week 24), 2.00 (3 months postnatal), and 3.21 (6 months postnatal).
Whole-brain voxel-wise associations
We did not reveal positive or negative associations between EDPS score at gestational week 24 and child brain ReHo map of left amygdala when child sex and age at scan were controlled.
Significant positive association between mothers’ EPDS score at 3 months postnatal and child brain ReHo map of left amygdala region was found when child sex and age at scan were controlled (thresholded at p < 0.005, p = 0.002 FWE corrected, cluster size [kE] 1522). The association remained significant when EPDS score at gestational week 24 was controlled in addition (thresholded at p < 0.005, p = 0.009 FWE corrected, kE 1196). Also significant positive association between mothers’ EPDS score at 3 months postnatal and child brain ReHo map of left amygdala region was found when maternal pre-pregnancy BMI, maternal SES, and ponderal index were set as covariates (thresholded at p < 0.005, p = 0.007 FWE corrected, kE 1281) and the association remained significant when EPDS score at gestational week 24 was controlled as well (thresholded at p < 0.005, p = 0.008 FWE corrected, kE 1213, Figure 1). The associations did not remain significant when SCL-90 score was included as a covariate in addition. This might be because of correlations between EPDS and SCL scores and indeed both depressive symptoms and anxiety symptoms associated with the left amygdala ReHo values (Supplemental Table A). There were no negative associations between mothers’ EPDS score at 3 months postnatal and ReHo map of the left amygdala region.Figure 1.ReHo values of the left amygdala associate positively with EPDS scores at 3 months postnatal; controlling for child sex, age at scan, maternal BMI, maternal socioeconomic status, ponderal index, and EPDS score at gestational week 24. In addition to the amygdala, the cluster extends to the surrounding cortex, the anterior hippocampus, the cerebellum, and the globus pallidus (all on the left hemisphere). The results have been thresholded at p < 0.005, FWE multiple comparisons corrected at the cluster level (p < 0.0125). The color bars depict t-values. Left hemisphere is on the right-hand side.
No statistically significant associations were found between EPDS score at 6 months postnatal and child brain ReHo map of left amygdala region with the whole-brain analyses. However, we observed a positive association between mothers’ EPDS score at 12 months postnatal and ReHo map of the left amygdala region when child sex and age at scan were set as covariates (thresholded at p < 0.005, p = 0.005 FWE corrected, kE 1325, Figure 2), but not with any other covariates.Figure 2.ReHo values of the left amygdala associate positively with EPDS scores at 12 months postnatal; controlling for child sex and age at scan. The cluster extends superiorly to the left globus pallidus and thalamus. The results have been thresholded at p < 0.005, FWE multiple comparisons corrected at the cluster level (p < 0.0125). The color bars depict t-values. Left hemisphere is on the right-hand side.
There were no negative associations between child amygdala ReHo values and maternal EPDS scores in any of the time points. We did not find significant associations in seed-based connectivity analysis of our seed region of interest, bilateral amygdala, to the rest of the brain, and the EPDS scores. T-maps of all the presented GLMs are shown in Supplementary Figures S8–S24 (thresholded at voxel-level p < 0.005 [uncorrected] and visualized at t > 2).
Region-of-interest analyses of ReHo for bilateral amygdala
Summary of all statistically significant results of the linear regression analyses for bilateral amygdala mean ReHo values are shown in Table 2. Additionally, summary of all linear regression analysis for left amygdala mean ReHo values are shown in Supplementary Table A, and for right amygdala mean ReHo values in Supplementary Table B.Table 2.Summary of the statistically significant results of the regression analyses for bilateral amygdala mean ReHo valuesRelationControlled covariates β
p Left amygdala ReHo and EPDS sum score at gestational week 24Child sex and age at scan0.25*<0.05Left amygdala ReHo and EPDS sum score at 3 monthsChild sex and age at scan0.48<0.001Left amygdala ReHo and EPDS sum score at 3 monthsChild sex, age at scan, maternal SES, maternal pre-pregnancy BMI, and ponderal index0.47<0.001Left amygdala ReHo and EPDS sum score at 3 monthsChild sex, age at scan, maternal SES, maternal pre-pregnancy BMI, ponderal index, and SCL–90 sum score at 3 months0.34<0.05Right amygdala ReHo and EPDS sum score at 3 monthsChild sex and age at scan0.28<0.05Right amygdala ReHo and EPDS sum score at 3 monthsChild sex, age at scan, maternal SES, maternal pre-pregnancy BMI and ponderal index0.29<0.05Left amygdala ReHo and EPDS sum score at 6 monthsChild sex and age at scan0.38<0.01Left amygdala ReHo and EPDS sum score at 6 monthsChild sex, age at scan, maternal SES, maternal pre-pregnancy BMI, and ponderal index0.37<0.01Left amygdala ReHo and EPDS sum score at 12 monthsChild sex and age at scan0.42<*0.001Left amygdala ReHo and EPDS sum score at 12 monthsChild sex, age at scan, maternal SES, maternal pre-pregnancy BMI, and ponderal index0.39<0.01Left amygdala ReHo and maternal pre-pregnancy BMIChild sex, age at scan, maternal SES, ponderal index, and EPDS sum score at 6 months−0.25<0.05Left amygdala ReHo and maternal pre-pregnancy BMIChild sex, age at scan, maternal SES, ponderal index, EPDS sum score at 6 months, and SCL–90 sum score at 6 months−0.25<0.05Abbreviations: BMI, body mass index; EPDS, Edinburgh Postnatal Depression Scale; ReHo, regional homogeneity; SCL-90, Symptom Checklist 90; SES, socioeconomic status.
There was a significant positive association between left amygdala mean ReHo value and EPDS score at gestational week 24 when child sex and age at scan were controlled (p < 0.05, β = 0.25). This association did not persist when maternal SES, maternal pre-pregnancy BMI, and ponderal index nor when SCL-90 score at gestational week 24 were statistically controlled for. There were no associations with the right amygdala mean ReHo value and EPDS score at gestational week 24.
We found a positive association between mean left amygdala mean ReHo value and EPDS score at 3 months postnatal when child sex and age at scan were included as covariates (p < 0.001, β = 0.48), and when maternal SES, maternal pre-pregnancy BMI, and ponderal index were controlled in addition (p < 0.001, β = 0.47). When SCL-90 score at 3 months postnatal was additionally set as covariate the association attenuated (p < 0.05, β = 0.34). Right amygdala mean ReHo value and EPDS score at 3 months postnatal had statistically significant association when child sex and age at scan were controlled (p < 0.05, β = 0.28), and the association remained significant when additionally controlling for maternal SES, maternal pre-pregnancy BMI, and ponderal index (p < 0.05, β = 0.29), but did not remain significant when SCL-90 score at 3 months postnatal was controlled in addition.
Left amygdala mean ReHo value and EPDS score at 6 months postnatal had a positive association when sex and age were controlled (p < 0.01, β = 0.38). The same association was found when maternal SES, maternal pre-pregnancy BMI, and ponderal index were controlled in addition (p < 0.01, β = 0.37). The association did not remain significant when SCL-90 score at 6 months postnatal was controlled in addition. There were no associations with right amygdala mean ReHo value and EPDS score at 6 months. Finally, when EPDS score at 6 months postnatal, child sex, age at scan, maternal SES, and ponderal index were controlled, we found association between left amygdala ReHo and maternal pre-pregnancy BMI (p < 0.05, β = −0.25) and the association remained when SCL-90 score at 6 months postnatal was controlled in addition.
There was statistically significant association between left amygdala mean ReHo value and EPDS score at 12 months postnatal when child sex and age at scan were controlled (p < 0.001, β = 0.42), but the association attenuated when maternal SES, maternal pre-pregnancy BMI, and ponderal index were controlled in addition (p < 0.01, β = 0.39). No associations between right amygdala mean ReHo values and EPDS score at 12 months postnatal were found.
Discussion
In this study, we investigated whether maternal perinatal depressive symptoms associate with child’s bilateral amygdala ReHo and seed-based functional connectivity. The voxel-wise analyses of ReHo implicated that the left amygdala had a strong positive association to EPDS scores obtained 3 months postnatally, and similar associations were detected for EPDS scores at 6 and 12 months postnatal albeit with smaller effect sizes. Association between left amygdala ReHo values and depressive symptoms did not persist when maternal anxiety symptoms were controlled in the whole-brain analysis but were the strongest predictor in the ROI analyses (Supplementary Table A). Seed-based connectivity analysis did not reveal any significant associations in the connectivity of the left amygdala and the rest of the brain. We were thus unable to replicate prior findings by Soe et al., who reported that maternal perinatal depression was associated with altered functional connectivity of the amygdala with the corticostriatal circuitry, especially with the OFC, insula, subgenual ACC, temporal pole, and striatum [13]. Lack of replication is important to note but is not rare in the field. It is also not a sign of contradictory findings. Our sample size was significantly smaller (N = 68) than Soe et al. (N = 128), from a different ethnic background, and we used task-free fMRI with video stimuli, which might explain the absence of their SCA findings in our results.
Our study showed that perinatal maternal depressive symptoms were associated with heightened ReHo, that is, heightened local functional activity, in the offspring’s left amygdala. An increase in ReHo may indicate heightened local synchronization in the amygdala or more tightly synchronized activity within the amygdala subregions, potentially exhibiting hyperactivity and hyper-responsiveness. These findings are intriguing given that the association was relatively strong and particularly localized in the left amygdala region in the whole-brain statistical models. Research of the mechanisms underlying depression have shown that left amygdala hyperactivity increases the risk for depression [26–28]. However, we note that the spatial resolution of our fMRI sequence does not allow reliable delineation of amygdala subregions, and that in lack of joint functional studies testing the activation profile of the amygdalae are unable to link the increased local connectivity to the direction of functional activation (positive, negative, or alternating patterns of both). Taken together, our findings raise the question of the possibility that amygdala regional connectivity may serve as a neural marker of intergenerational transmission of risk for depression and other affective disorders. However, this remains to be addressed in future studies. It is interesting to note that the associations were not visible in the hippocampi, though both amygdalae and hippocampi have been associated with prenatal and postnatal maternal depressive symptoms [29, 30].
The effect of postnatal maternal depressive symptoms on child’s developing brain mediates via parenting and mother–child interaction [31, 32]. Infancy and toddlerhood are crucial periods for the development of child’s self- and emotion regulation, cognitive, language, and motor skills [33–35]. By the age of 2–3 months, infants begin responding to social interactions, for example, smiles, recognizing familiar faces, and facial expressions and communicating through vocalization [36–38]. At this stage, socioemotional development begins to emerge, and the child is highly dependent on the caregiver’s emotional support (i.e., emotional coregulation). These early developmental steps could be especially sensitive to changes in maternal mood, and based on our findings, could affect later brain development. A key feature of depression is dysfunction of self- and emotion regulation [39]. Therefore, maternal depressive symptoms may affect the self-regulation, sensitivity of the mother, and reduce prosocial interaction toward the child. Sensitive interaction implies mother’s ability to interpret child’s physical needs and emotional cues and to response appropriately [40]. When this sensitivity is disrupted, as often seen in cases of postnatal depression, it can have lasting effects: preschool-aged children of mothers experiencing postnatal depression had more internalizing and externalizing behavioral problems, that is, altered self- and emotion regulation [41,42]. Moreover, postnatal depression has been found to be negatively associated with white matter integrity in vast areas in 5-year-old girls [43].
Limitations
Although our sample size is relatively small, our results are compatible with the few previous studies investigating similar effects using fMRI [9, 13]. Nevertheless, studies with larger cohorts are needed in the future. The study population’s cultural background was narrow and therefore it is needed to further study this association in other populations. As the children were scanned at one time point, further studies are needed to determine whether amygdala functional changes persist or evolve across older ages. Regarding the limitations of fMRI acquisition, the use of a passive viewing paradigm to reduce head motion [44, 45] and the presence of the parent in the imaging room may have affected the measured fMRI signal. The EPDS and SCL-90 scores are self-reported measures and majority of the participating mothers did not have clinically meaningful or severe symptoms of perinatal depression.
Conclusions
In conclusion, we found that maternal perinatal depressive symptoms were associated with heightened local functional connectivity of the left amygdala in 5-year-olds. We did not find associations in the patterns of the left amygdala connectivity to the rest of the brain. Our results provide preliminary indication that maternal mental health during pregnancy and especially soon after birth may influence offspring’s neurodevelopment related to emotional regulation.
Supporting information
10.1192/j.eurpsy.2025.10097.sm001Vartiainen et al. supplementary materialVartiainen et al. supplementary material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1den Bergh BRHV, Dahnke R, Mennes M. Prenatal stress and the developing brain: Risks for neurodevelopmental disorders. Dev Psychopathol. 2018;30(3):743–62.30068407 10.1017/S 0954579418000342 · doi ↗ · pubmed ↗
- 2Lautarescu A, Craig MC, Glover V. Prenatal stress: Effects on fetal and child brain development. In: International review of neurobiology [internet]. Elsevier; 2020. pp. 17–40 [cited 2023 Mar 10]. Available from: https://linkinghub.elsevier.com/retrieve/pii/S 007477421930119910.1016/bs.irn.2019.11.00232204831 · doi ↗ · pubmed ↗
- 3Gavin NI, Gaynes BN, Lohr KN, Meltzer-Brody S, Gartlehner G, Swinson T. Perinatal depression: A systematic review of prevalence and incidence. Obstet Gynecol. 2005;106(5 Part 1):1071.16260528 10.1097/01.AOG.0000183597.31630.db · doi ↗ · pubmed ↗
- 4Rifkin-Graboi A, Bai J, Chen H, Hameed WBR, Sim LW, Tint MT, et al. Prenatal maternal depression associates with microstructure of right amygdala in neonates at birth. Biol Psychiatry. 2013;74(11):837–44.23968960 10.1016/j.biopsych.2013.06.019 · doi ↗ · pubmed ↗
- 5Wen DJ, Poh JS, Ni SN, Chong YS, Chen H, Kwek K, et al. Influences of prenatal and postnatal maternal depression on amygdala volume and microstructure in young children. Transl Psychiatry. 2017;7(4):e 1103.28440816 10.1038/tp.2017.74PMC 5416711 · doi ↗ · pubmed ↗
- 6Mandl S, Alexopoulos J, Doering S, Wildner B, Seidl R, Bartha-Doering L. The effect of prenatal maternal distress on offspring brain development: A systematic review. Early Hum Dev. 2024;192:106009.38642513 10.1016/j.earlhumdev.2024.106009 · doi ↗ · pubmed ↗
- 7Bale TL, Baram TZ, Brown AS, Goldstein JM, Insel TR, Mc Carthy MM, et al. Early life programming and neurodevelopmental disorders. Biol Psychiatry. 2010;68(4):314–9.20674602 10.1016/j.biopsych.2010.05.028PMC 3168778 · doi ↗ · pubmed ↗
- 8Posner J, Cha J, Roy AK, Peterson BS, Bansal R, Gustafsson HC, et al. Alterations in amygdala-prefrontal circuits in infants exposed to prenatal maternal depression. Transl Psychiatry. 2016;6(11):e 935–5.27801896 10.1038/tp.2016.146PMC 5314110 · doi ↗ · pubmed ↗