Synergistic potential and apoptosis induction of Bunium persicum essential oil and its pure components, cuminaldehyde and γ-terpinene, in combination with fluconazole on Candida albicans isolates: in vitro and in silico evaluation
Mahsa Asgar, Mehdi Bamorovat, Seyyed Amin Ayatollahi Mosavi, Fatemeh Sharifi, Ehsan Salarkia, Ali Karamoozian, Sanaz Hadizadeh, Setareh Agha Kuchak Afshari

TL;DR
This study shows that combining Bunium persicum essential oil and its components with fluconazole can enhance antifungal effects and induce cell death in Candida albicans.
Contribution
The study demonstrates the synergistic and apoptotic potential of natural compounds with fluconazole against resistant Candida isolates.
Findings
Cuminaldehyde showed the lowest minimum inhibitory and fungicidal concentrations compared to other agents.
Fluconazole combined with cuminaldehyde had the highest synergistic effect (63.6%) against Candida isolates.
Molecular docking confirmed that cuminaldehyde and γ-terpinene interact with lanosterol 14-alpha-demethylase.
Abstract
Fluconazole resistance in Candida species is on the rise, posing a significant clinical challenge. There is a growing interest in using complementary therapies, especially those from natural sources. This study aimed to evaluate the synergistic and apoptotic effects of Bunium persicum essential oil (BPEO) and its two pure components, cuminaldehyde (CA) and γ-terpinene (γ-TPN), combined with fluconazole (FLC) on susceptible and resistant C. albicans isolates. Moreover, molecular docking was used to study the interactions between lanosterol 14-alpha-demethylase and each agent. The BPEO was prepared using the Clevenger apparatus and the hydro-distillation method. The in vitro antifungal activity was evaluated according to the Clinical and Laboratory Standards Institute guideline (M60). The checkerboard and isobologram assays assessed the interaction between BPEO, CA, γ-TPN, and FLC. The…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
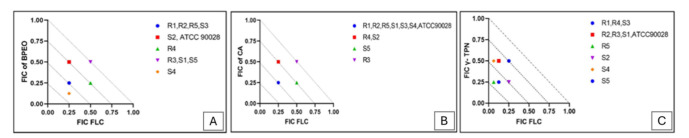 Figure 1
Figure 1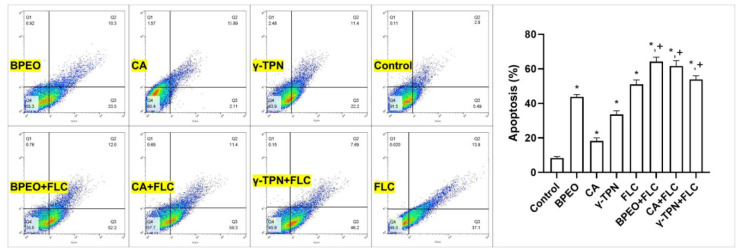 Figure 2
Figure 2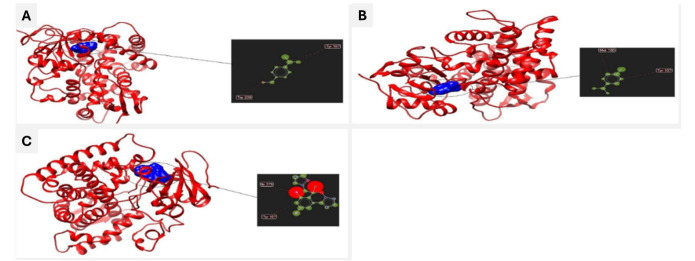 Figure 3
Figure 3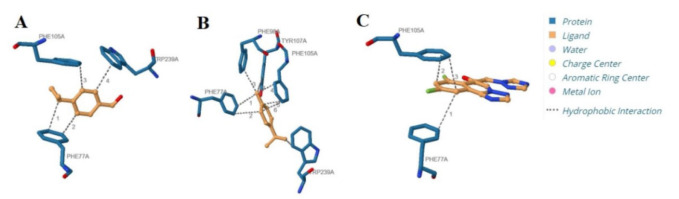 Figure 4
Figure 4| MIC (µg/ml) | MFC (µg/ml) | ||||||
|---|---|---|---|---|---|---|---|
| BPEO | CA | γ-TPN | FLC | BPEO | CA | γ-TPN | |
| R1 | 125 | 31.2 | 125 | 16 | 500 | 125 | 250 |
| R2 | 62.5 | 31.25 | 62.5 | 8 | 250 | 125 | 125 |
| R3 | 62.5 | 62.5 | 125 | 16 | 250 | 125 | 500 |
| R4 | 125 | 62.5 | 125 | 32 | 500 | 250 | 500 |
| R5 | 62.5 | 31.2 | 31.2 | 8 | 125 | 62.5 | 125 |
| S1 | 31.2 | 31.2 | 62.5 | 1 | 125 | 62.5 | 250 |
| S2 | 31.2 | 15.6 | 31.2 | 0.5 | 62.5 | 31.25 | 62.5 |
| S3 | 15.6 | 7.8 | 31.2 | 0.5 | 62.5 | 31.25 | 62.5 |
| S4 | 62.5 | 31.2 | 125 | 1 | 250 | 125 | 500 |
| S5 | 15.6 | 15.6 | 31.2 | 0.25 | 62.5 | 31.25 | 62.5 |
| ATCC 90028 | 31.2 | 7.8 | 15.6 | 0.5 | 62.5 | 31.25 | 62.5 |
| Compounds | MIC | β | % 95 CI for β | |||
|---|---|---|---|---|---|---|
| Mean ± SD | Median (IQR) | |||||
|
| 56.80±38.46 | 62.50 (31.30) | 49.19 | (32.22, 66.15) | <0.001 | |
|
| 29.80±18.76 | 31.20 (15.65) | 22.19 | (15.57, 28.81) | <0.001 | |
|
| 69.58±46.03 | 62.50 (93.80) | 61.97 | (39.49, 84.44) | <0.001 | |
|
| 7.61±10.14 | 1.00 (15.50) | 0 | |||
|
| ||||||
| Mean ± SD | Median (IQR) | β | % 95 CI for β | |||
|
| 56.80±38.46 | 62.50 (31.30) | 26.99 | (10.59, 43.40) | 0.001 | |
|
| 69.58±46.03 | 62.50 (93.80) | 39.78 | (21.04, 58.51) | <0.001 | |
|
| 29.80±18.76 | 31.20 (15.65) | 0 | |||
| MIC | ||||||
|
|
|
|
|
| ||
|
|
| 60.24±42.64 | 62.50 (78.18) | 39.08 | (11.50, 66.66) | 0.005 |
|
| 27.09±29.56 | 23.40 (28.50) | 0 | |||
|
| ||||||
|
|
| |||||
|
|
|
|
| |||
|
|
| 6 (54.50) | 5 (45.50) | 0.31 | (0.09, 1.02) | 0.055 |
|
| 7 (63.60) | 4 (36.40) | 0.21 | (0.06, 0.81) | 0.023 | |
|
| 3 (27.30) | 8 (72.70) | 1 | |||
| MIC | FICI | Interpretation | MIC | FICI | Interpretation | MIC | FICI | Interpretation | |
|---|---|---|---|---|---|---|---|---|---|
| BPEO/FLC | CA/FLC | γ-TPN/FLC | |||||||
| R1 | 31.25/4 | 0.5 | Synergism | 7.8/4 | 0.5 | Synergism | 31.25/4 | 0.5 | Synergism |
| R2 | 15.6/2 | 0.49992 | Synergism | 7.8/2 | 0.5 | Synergism | 15.6/4 | 0.74992 | Additive |
| R3 | 31.25/8 | 1 | Additive | 31.25/8 | 1 | Additive | 62.5/8 | 1 | Additive |
| R4 | 62.5/8 | 0.75 | Additive | 31.25/8 | 0.75 | Additive | 62.5/8 | 0.75 | Additive |
| R5 | 15.6/2 | 0.49992 | Synergism | 7.8/2 | 0.5 | Synergism | 7.8/2 | 0.5 | Synergism |
| S1 | 15.6/0.5 | 1 | Additive | 7.8/0.25 | 0.5 | Synergism | 31.25/0.5 | 1 | Additive |
| S2 | 7.8/0.25 | 0.75 | Additive | 7.8/0.125 | 0.75 | Additive | 15.6/0.125 | 0.75 | Additive |
| S3 | 3.9/0.125 | 0.5 | Synergism | 1.95/0.125 | 0.5 | Synergism | 7.8/0.125 | 0.5 | Synergism |
| S4 | 15.6/0.25 | 0.4996 | Synergism | 7.8/0.25 | 0.5 | Synergism | 31.25/0.5 | 0.75 | Additive |
| S5 | 7.8/0.125 | 1 | Additive | 3.9/0.125 | 0.75 | Additive | 15.6/0.125 | 1 | Additive |
| ATCC 90028 | 7.8/0.125 | 0.5 | Synergism | 1.95/0.125 | 0.5 | Synergism | 7.8/0.25 | 1 | Additive |
| Residue | Aminoacids | Distance | Ligand Atom | Protein Atom | |
|---|---|---|---|---|---|
| Cuminaldehyde | 77A | PHE | 3.41 | 3592 | 157 |
| 77A | PHE | 3.34 | 3596 | 156 | |
| 105A | PHE | 3.45 | 3597 | 382 | |
| 239A | TRP | 3.35 | 3599 | 1438 | |
| Gamma Terpinene | 77A | PHE | 3.54 | 3600 | 157 |
| 77A | PHE | 3.98 | 3594 | 156 | |
| 98A | PHE | 3.44 | 3600 | 329 | |
| 105A | PHE | 3.54 | 3600 | 37 | |
| 105A | PHE | 3.62 | 3596 | 381 | |
| 105A | PHE | 3.36 | 3594 | 382 | |
| 107A | TYR | 3.37 | 3600 | 398 | |
| 239A | TRP | 3.20 | 3598 | 1436 | |
| Fluconazole | 77A | PHE | 3.99 | 3607 | 156 |
| 105A | PHE | 3.86 | 3606 | 381 | |
| 105A | PHE | 3.78 | 3607 | 382 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsComputational Drug Discovery Methods
Introduction
Opportunistic fungal infections, such as candidiasis, have become a growing health problem worldwide, especially in high-risk individuals [ 1 ]. Candida albicans is the most important commensal species that can become an opportunistic pathogen and cause many infections [ 2 , 3 ]. Azole derivatives, such as fluconazole (FLC), are among the most common antifungal drugs used to treat Candida-related infections. The FLC inhibits the 14α-lanosterol demethylase, the key enzyme in the ergosterol biosynthesis pathway [ 4 ]. Antifungal resistance to the azole derivatives has recently become a serious clinical challenge since the increase in multidrug-resistant patterns could affect mortality rates [ 5 ]. Besides, some currently available antifungal agents have limitations regarding high cost, toxicity, and low drug bioavailability [ 6 , 7 ]. Therefore, exploring and finding novel therapeutic strategies for treating fungal infections that are highly efficacious and have lower side effects is crucial.
Numerous essential oils (EOs) have gained more attention as pivotal medicinal agents [ 8
- 10 ]. Reports have indicated that EOs might be a promising potential drug candidate against various diseases, regarding their pharmacological actions and low toxicity. Among traditional natural products, Bunium persicum, which belongs to the Apiaceae family, has received much attention for its beneficial medical effects. It is a significant grassy aromatic plant that naturally grows in some South Asian countries [ 11 ]. Numerous studies have reported that B. persicum EO (BPEO) has antioxidant, anti-inflammatory, antimicrobial, antifungal, and antileishmanial properties [ 12 , 13 ]. These promising therapeutic effects of the BPEO have been attributed to the bioactivity of its major components [ 11 ]. Cuminaldehyde (CA; 4-isopropylbenzaldehyde) is one of the major components, which has anticancer, antidiabetic, anti-inflammatory, and antimicrobial activity, and is a food preservative due to its powerful fumigant against phytopathogenic fungi [ 14
- 16 ]. The other substance in the BPEO is gamma-terpinene, which has effective pharmacological properties, such as antimicrobial and antifungal activities [ 17
- 19 ].
In addition, in recent years, combination therapy and synergistic interaction have been well-known as feasible strategies for combating drug resistance [ 20
- 22 ]. The synergy between azoles and EOs could permit the usage of lower doses of these synthetic antifungal agents effectively and safely, presenting a promising pharmaceutical strategy for future management of mycoses. Furthermore, the synergistic properties of combining medicinal plants with available antifungal drugs might enhance antifungal activities. Hence, this study aimed to evaluate the inhibitory activity of the BPEO, compared to its two major pure compounds, including CA and γ-terpinene (γ-TPN), as well as their synergistic effects and apoptosis-inducing potential in combination with FLC against both susceptible and resistant C. albicans isolates. Molecular docking was also used to study how Lanosterol 14 alpha-demethylase interacts with two pure components, CA and γ-TPN, in search of antifungal properties.
Materials and Methods
Essential oils and compounds
The B. persicum seeds were prepared from the market and confirmed by a pharmacognosist in the Department of Pharmacognosy at Kerman University of Medical Sciences, Kerman, Iran (herbarium number KF1141). The BPEO was prepared by the Clevenger apparatus and the hydrodistillation method as described previously, with some modifications [ 13 ]. The obtained BPEO was kept at 20 °C until the experiment. The FLC (Pfizer, Groton, CT, USA) and pure compounds, CA and γ-TPN (Sigma-Aldrich, St. Louis, USA), were bought at 97% purity.
Fungal isolates
This study was conducted on a standard C. albicans strain (ATCC 90028) and 10 clinical C. albicans isolates obtained from patients with vulvovaginal candidiasis, which had been previously identified using conventional and molecular methods [ 23 ].
In vitro antifungal susceptibility testing
The minimum inhibitory concentration (MIC) of agents was evaluated by the microdilution broth method according to the Clinical and Laboratory Standards Institute (CLSI M60) guideline [ 24 ]. According to the CLSI clinical breakpoint values, C. albicans isolates were considered susceptible with MIC ≤ 2 μg/mL and resistant with MIC ≥ 8 μg/mL for FLC [ 24 ]. The FLC and BPEO were prepared at a final concentration of 128-0.125 µg/ml and 500-1.95 µg/ml, respectively. A serial dilution of CA and γ-TPN was prepared from 250-0.24 µg /ml [ 25 , 26 ]. The suspension containing 0.5–2.5 × 10^3^ cells/mL of each C. albicans isolate was added to the 96-well microtiter plates containing RPMI-1640 medium (Sigma-Aldrich, USA). The plates were incubated at 35 °C for 24 h and afterward read visually. All experiments were conducted in triplicate using C. parapsilosis ATCC 22019 as the quality control strain. To assess fungicidal activity, 10 μL of cell suspensions from turbidity-free wells were cultured on Sabouraud dextrose agar plates, incubated at 35 ºC for 48 h, and the number of colony-forming units was determined. The minimum fungicidal concentration (MFC) was defined as the lowest concentration at which three or fewer C. albicans colonies were observed [ 27 ].
Assessment of the drug interaction
The microdilution checkerboard assay was performed to assess the fractional inhibitory concentration index (FICI) [ 28 ]. For this purpose, combinatorial effects between FLC and BPEO/CA/γ-TPN were evaluated in the 96-well microtiter plates. Serial two-fold dilutions of each compound, ranging from several dilutions below the MIC to 2 × MIC, were set up. All experiments were tested in duplicate. The FICI was measured as follows: FICI = FIC A + FIC B. The FIC of each agent was determined as follows: FIC A = MIC of drug A in combination / MIC of drug A alone, and FIC B = MIC of drug B in combination / MIC of drug B alone.
The corresponding FIC values of the checkerboard assay were represented graphically by isobologram analysis using Prism software (version 9.0; GraphPad Inc., San Diego, CA). The straight line connecting the intercept points on the x-axis and y-axis signifies an additive interaction between the drugs (FICI=1). Values below this line indicate synergistic (FICI≤0.5) or additive effects (0.5<FICI≤1), while values above suggest indifferent (1<FICI<4) or antagonistic effects (FICI>4) [ 29 ].
Apoptotic effects analysis
The approach of cell death induced by BPEO, CA, and γ-TPN, alone and in combination with FLC, was investigated using the fluorescein isothiocyanate Annexin V Apoptosis Detection Kit I (BD Biosciences, CA, USA) as previously described [ 30 ]. The C. albicans standard strain was exposed to minimum inhibitory concentrations of each tested compound and incubated at 35 °C for 48 h. After incubation, the protoplast of yeast cells was prepared and resuspended in an annexin V binding buffer at a concentration of 1×10^6^ cells/mL. Subsequently, 5 µL of fluorescein isothiocyanate annexin V and Propidium iodide were added to the 100 µL of this solution and kept at room temperature for 15 min. Subsequently, the annexin V binding buffer (400 µL) was used in each tube, and the samples were investigated by a flow cytometer (BD LSRFortessa cell analyzer, Becton Dickinson, USA). Untrained cells were used as controls, and the data obtained were analyzed with FlowJo software (version 10).
In silico analysis
Protein-ligand docking
The 3D structure of FLC, CA, γ-TPN, and Lanosterol 14 alpha-demethylase was achieved by the Protein Data Bank (https://www.rcsb.org/) [ 31 ]. The HDOCK server (©Lab of Biophysics and Molecular Modeling, [email protected]) utilized peptide-protein docking to determine the receptor association (http://hdock.phys.hust.edu.cn/). The root-mean-square deviation method was applied to assess the difference in the location of the docked peptides. The default parameters were used for all docking runs. The HDOCK server applies an IT Score-PP iterative knowledge-based scoring system (Docking score).
2D interaction plots of protein-ligand complexes
The Protein-Ligand Interaction Profiler site (using the Michael Schroeder group at the Biotechnology Center TU Dresden (BIOTEC) at https://academic.oup.com/nar/article/49/W1/W530/6266421) was applied to assess the ligand interaction site (https://plip-tool.biotec.tu-dresden.de/plip-web/plip/index) after the docking procedure [ 32 ]. Afterward, the interactions between the amino acid residues of the target protein and the ligand were determined. Lastly, the 3D shapes of the final interaction were designed using the UCSF Chimera software (version 1.12; University of California, San Francisco, USA).
Statistical analyses
Statistical analyses were carried out using the generalized estimating equation method based on the Gaussian distribution using logistic regression and linear regression models. An exchangeable correlation structure was used for this method. The SPSS software (version 22.0; IBM, Armonk, NY, USA) was used for data analysis. A p value of less than 0.05 was considered statistically significant.
Results
In vitro susceptibility testing results
Table 1 shows the MIC and MFC values obtained for BPEO, CA, γ-TPN, and FLC against all C. albicans isolates. Based on the antifungal susceptibility testing, clinical C. albicans isolates are categorized as different types of FLC-susceptible (n=5) and FLC-resistant (n=5) species. The MIC values for FLC against resistant and susceptible C. albicans isolates were within 8-32 and 0.25-1 µg/ml ranges, respectively. The MIC values for BPEO and γ-TPN ranged from 125 to 15.6 µg /ml, while the MIC for CA ranged from 62.5 to 7.8 µg /ml. The MFC values of BPEO and γ-TPN varied from 500 to 62.5 µg /ml, while for CA, it ranged from 250 to 31.25 µg /ml.
Table 2 shows the mean MIC values obtained for BPEO, CA, γ-TPN, and FLC. According to the statistical analyses, there is a significant difference between the MIC values of FLZ and those of three natural compounds, including BPEO, CA, and TPN (p<0.001). Besides, CA had a lower MIC value than γ-TPN and BPEO. In addition, there were significant differences between resistant and sensitive isolates regarding MIC values of BPEO and active compounds, including CA and γ-TPN. Moreover, the MIC values for FLC-resistant isolates were more than those for sensitive isolates (p =0.005).
Checkerboard assay results
The checkerboard microtiter test evaluated the most effective combination of components with FLC. The obtained results and interpretation are reported in Table 3. Based on the FICI values, CA was the most effective agent with synergistic activity against seven isolates, while the γ-TPN exhibited the least synergistic activity with only three isolates (n=3). According to the statistical analyses, there was no significant difference between the FICI of FLC/CA and FLC/BPEO regarding synergistic and non-synergistic activity (Table 2). An important difference existed between the FICI of FLC/CA and FLC/γ-TPN. Moreover, the combination of FLC and CA demonstrated a higher odds ratio (OR) for synergistic effect, compared to the combination of FLC and γ-TPN (p=0.023). No antagonistic activity was observed, and the isobolographic analysis supported the synergistic interaction between FLC and the tested agents (Figure 1).
Isobologram analysis of synergistic interaction between fluconazole (FLC) and A) Bunium persicum essential oil (BPEO), B) cuminaldehyde (CA), and C) γ-terpinene (γ-TPN) against clinical resistance (R) and susceptible (S) Candida albicans isolates. FLC–fractional inhibitory concentration (FIC) values are drafted on the x-axis, while BPEO/CA/γ-TPN–FIC values are drafted on the y-axis.
Apoptosis and necrosis result
The flow cytometry results showed the apoptosis and necrosis effects on the C. albicans isolate exposed to each agent alone and in combination with FLC (Figure 2). The sum of early and late apoptosis percentages was measured for each sample to calculate the level of apoptotic cells. Briefly, when BPEO, CA, and γ-TPN were used alone, they induced apoptosis (primary/final) in 43.8%, 17.91%, and 33.6% of the yeast cells, respectively. Furthermore, the flow cytometry results revealed that the combination of all natural components with FLC had a significant apoptotic effect on the C. albicans isolate, compared to using each agent alone (p<0.001).
Flow cytometry analysis of Candida albicans exposed to the minimum inhibitory concentrations of Bunium persicum essential oil (BPEO), cuminaldehyde (CA), γ-terpinene (γ-TPN), and fluconazole (FLC) alone, and the combination of each agent with FLC, compared with untreated control after 48 h incubation. Q1: necrosis (annexin V-/propidium iodide+), Q2: late apoptosis (annexin V+/ propidium iodide+), Q3: early apoptosis (annexin V+/PI-), and Q4: viable cells (annexin V-/ propidium iodide-). Bars show the mean ± standard deviation of viability rates (n=3). Significant differences compared to the control group are marked with an asterisk (), while differences between combination and single usage of each agent are indicated with a plus sign (+) (p<0.001).
In silico docking results
Figure 3 illustrates the 3D interactions of CA, γ-TPN, and FLC with Lanosterol 14 alpha-demethylase, using Chimera software. As shown in Figure 3, CA can bind to Lanosterol 14 alpha-demethylase at active site residues Tyrosine107 and Tryptophan 239 (Figure 3A), γ-TPN at active site residues Tyrosine107 and Methionine100 (Figure 3B), and FLC at active site residues Tyrosine107 and Isoleucine379 (Figure 3C). In addition, the docking scores of CA, γ-TPN, and FLC with lanosterol 14 alpha-demethylase were -82.40, -77.19, and -156.82, respectively. Furthermore, the results of Ligand root-mean-square deviation (Å) were 80.59, 80.00, and 75.57, respectively. The amino acids involved in the interaction of CA, γ-TPN, and FLC core pocket with Lanosterol 14 alpha-demethylase are shown in Figure 4. It must be mentioned that Tyr107 is the common amino acid affirmed with Molegro Molecular Viewer 2.5.0 (Molegro ApS, Aarhus, Denmark). Table 4 displays ligands of hydrogen bonds and hydrophobic interactions.
The 3D interactions of Lanosterol 14 alpha-demethylase and A) cuminaldehyde, B) γ-terpinene, and C) fluconazole using Chimera software (target cavities show the interaction of the ligand with residues where the amino acids are involved).
Ligands of A) cuminaldehyde/Lanosterol 14 alpha-demethylase, B) γ-terpinene/Lanosterol 14 alpha-demethylase, and C) fluconazole/Lanosterol 14 alpha-demethylase by the Protein-Ligand Interaction Profiler server.
Discussion
Treatment of Candida infections due to the drug-resistant species is becoming a crucial concern worldwide [ 33 ]. Besides, since higher treatment doses of currently available antifungal agents can lead to various adverse effects, choosing the proper alternative therapies is crucial. Over recent years, traditional herbal medicine has been widely useful in fungal infection therapy due to its broad-spectrum activity and low toxicity [ 10 ]. Bunium persicum EO and its bioactive components have been used due to their promising medical activities all around the world [ 13 , 34 ]. In this regard, the present research examined the in vitro antifungal activity of BPEO, compared to its main components, CA and γ-TPN, against FLC-susceptible and -resistant C. albicans isolates.
The results confirmed the inhibitory effect of BPEO and its two pure components against both susceptible and resistant C. albicans isolates. According to the statistical analyses, CA showed the lowest MICs and MFCs compared to BPEO and γ-TPN in the present study. Furthermore, numerous previous studies have reported antimicrobial and antifungal activities of CA [ 35 , 36 ]. Sekine et al. evaluated the antifungal effects of different volatile compounds against phytopathogenic fungi and reported that B. persicum had the most antifungal activity [ 12 ]. In addition, their findings displayed CA as the main antifungal compound with stronger antifungal activity against F. oxysporum, compared to other compounds in black Zira [ 12 ]. Findings of the present study demonstrated the highest MICs for γ-TPN, which is consistent with those of a study performed by Mandras et al. They reported no anticandidal effect of γ-TPN among the tested EOs and their bioactive pure compounds, while α-pinene displayed effective anticandidal activity against non-albicans Candida isolates [ 37 ]. Another study investigated the potential of 50 EOs against Candida biofilms. In this work, γ-TPN exhibited a moderate or weak correlation to biofilm inhibition [ 38 ]. According to the findings, it is inferred that the antifungal activity of BPEO in the present study was mainly due to the presence of CA.
To the best of our knowledge, no studies have been performed on the synergistic and apoptotic effects of BPEO and its pure bioactive compounds in combination with FLC against clinically susceptible and resistant C. albicans isolates. In the present study, the FICI results indicated that CA combined with FLC had a synergistic effect against seven C. albicans strains, with no significant differences between the susceptible and resistant strains. The synergistic effect was obtained in three isolates when γ-TPN was combined with FLC. Moreover, CA showed inhibitory effects in combination with FLC at concentrations lower than their individual MIC values. Some previous studies have confirmed the synergistic activity of EOs and their main bioactive compounds in combination [ 36 , 39 ]. Touil et al. reported that combining CA and carvacrol caused synergistic interactions in the most tested C. albicans strains [ 36 ].
In the current study, flow cytometry results showed that BPEO, CA, and γ-TPN had apoptosis and necrosis effects. More remarkably, the combination of these natural compounds with FLC showed higher apoptotic effects than each tested agent alone. Some studies have reported the necrotic and apoptotic effects of other natural antimicrobial compounds on Candida species [ 40 ]. Results of the present study are consistent with those of another study, which reported a significantly greater apoptosis rate in the combination group, compared to the other groups [ 40 ]. Generally, the processes of apoptosis and necrosis can be induced by different pathways. Since there is no data about the probable induction mechanisms of apoptosis and necrosis by B. persicum and its derivatives, future research is necessary to consider these compounds as suitable new antimycotic agents.
In addition, a computational study was conducted in this research to determine the mechanism of synergistic interaction. As mentioned in the results section, natural composition exhibits a synergistic effect via multiple targets through binding to Lanosterol 14 alpha-demethylase with various active site residues. Lanosterol 14α-demethylase is a crucial enzyme in the ergosterol biosynthesis pathway, which is vital for maintaining the integrity of fungal cell membranes. Inhibition of this enzyme disrupts the production of ergosterol, which is the primary mechanism by which azole antifungals, such as FLC, operate. Fluconazole is commonly used to treat Candida infections. However, with the rising prevalence of FLC-resistant Candida strains, there is an urgent need to explore alternative or adjunctive compounds that can either inhibit this enzyme or enhance the effectiveness of the existing azole antifungals. Molecular modeling studies suggest that most natural compounds demonstrate antifungal activity by inhibiting the 14α-demethylase enzyme [ 41 ]. The findings indicate that the presence of a hydroxyl group significantly enhances the activity of these compounds.
The highlighted amino acids in Lanosterol 14α-demethylase—specifically Tyr107, Trp239, Met100, and Ile379—were identified as crucial based on molecular docking analysis results. These residues were selected since they formed direct interactions with the docked ligands (CA, γ-TPN, and FLC), as determined by the Protein-Ligand Interaction Profiler tool following the docking procedure. Their functional relevance is supported by their location within or near the active site of the enzyme, which is vital for substrate binding and catalysis. For example, Tyr107 and Trp239 often help stabilize ligand binding through hydrogen bonds or hydrophobic interactions, while residues, like Met100 and Ile379, contribute to the structural configuration of the active site, impacting the function of the enzyme and how it interacts with inhibitors [ 42 , 43 ].
The docking analysis performed in the present study revealed that CA and γ-TPN interact with Lanosterol 14α-demethylase at sites distinct from the FLC binding pocket, suggesting a potential allosteric or structural modulatory mechanism. Such interactions may induce conformational changes or destabilize the enzyme, thereby impairing its activity and enhancing the efficacy of FLC. This mechanistic hypothesis is consistent with the in vitro findings of synergistic effects in the present research and warrants further investigation through mutagenesis or biophysical studies. Nevertheless, since the in vivo interaction mode of the two molecules is affected by various physiological factors, further studies are essential to confirm the present findings.
Conclusion
In conclusion, the findings displayed that CA has more effective activity than the EO of B. persicum and γ-TPN against both FLC-resistant and -susceptible C. albicans isolates. Furthermore, the synergistic effects due to the combination of these natural products with available synthetic antifungal agents could contribute to the optimal inhibition strategies, particularly in resistant fungal strains. However, the possible mechanism of action of B. persicum EO and its pure compounds alone and combined with commercial antifungal drugs requires further studies to confirm their potential for clinical application.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Vázquez‐González D Perusquía‐Ortiz AM Hundeiker M Bonifaz A Opportunistic yeast infections: candidiasis, cryptococcosis, trichosporonosis and geotrichosis J Dtsch Dermatol Ges 2013115381932362133010.1111/ddg.12097 · doi ↗ · pubmed ↗
- 2Reda NM Hassan RM Salem ST Yousef RHA Prevalence and species distribution of Candida bloodstream infection in children and adults in two teaching university hospitals in Egypt: first report of Candida kefyr Infection 2023512389953601849310.1007/s 15010-022-01888-7PMC 10042939 · doi ↗ · pubmed ↗
- 3Hamzehee S Kalantar-Neyestanaki D Afshari SAK Mousavi SAA Molecular identification of Candida species, assessment of the antifungal susceptibility and the genetic relationship of Candida albicans isolated from immunocompromised patients in Kerman, Iran Gene Rep 201917100484
- 4Odds FC Brown AJ Gow NA Antifungal agents: mechanisms of action Trends Microbiol 200311627291282394410.1016/s 0966-842x(03)00117-3 · doi ↗ · pubmed ↗
- 5Hadrich I Ayadi A Epidemiology of antifungal susceptibility: review of literature J Mycol Med 2018283574842977343510.1016/j.mycmed.2018.04.011 · doi ↗ · pubmed ↗
- 6Brammer K Farrow PR Faulkner J Pharmacokinetics and tissue penetration of fluconazole in humans R Rev Infect Dis 199012 Supplement_3S 3182610.1093/clinids/12.supplement_3.s 3182184510 · doi ↗ · pubmed ↗
- 7Kneale M Bartholomew JS Davies E Denning DW Global access to antifungal therapy and its variable cost J Antimicrob Chemother 2016711235996062751647710.1093/jac/dkw 325 · doi ↗ · pubmed ↗
- 8Heard SC Wu G Winter JM Antifungal natural products Curr Opin Biotechnol 202169232413364059610.1016/j.copbio.2021.02.001 · doi ↗ · pubmed ↗