No evidence for human or rodent deltavirus replication in yeast
Louison Bois Naegelin, Valérie Courgnaud, Dominique Helmlinger, Karim Majzoub

TL;DR
This paper shows that human and rodent deltaviruses cannot replicate in yeast, suggesting yeast lacks necessary factors for their replication.
Contribution
The study introduces a genetic tool to test RNA virus replication in yeast and applies it to deltaviruses.
Findings
Hepatitis D and Rodent delta viruses cannot replicate in yeast.
Yeast lacks essential factors for deltavirus RNA amplification.
Abstract
Deltaviruses were recently discovered across a broad range of metazoan species. Their genome encodes a single protein and thus requires host factors for replication, which are likely evolutionarily conserved given their broad host range. Here we describe a genetic tool to unambiguously determine whether an RNA virus replicates in model yeast species. Our system involves transcriptional induction of viral RNA from an integrated cDNA template followed by its genomic excision. Testing Hepatitis D and Rodent delta viruses revealed that neither viral RNAs can replicate in yeasts, suggesting that Ascomycetes lack factors essential for their RNA-dependent amplification
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
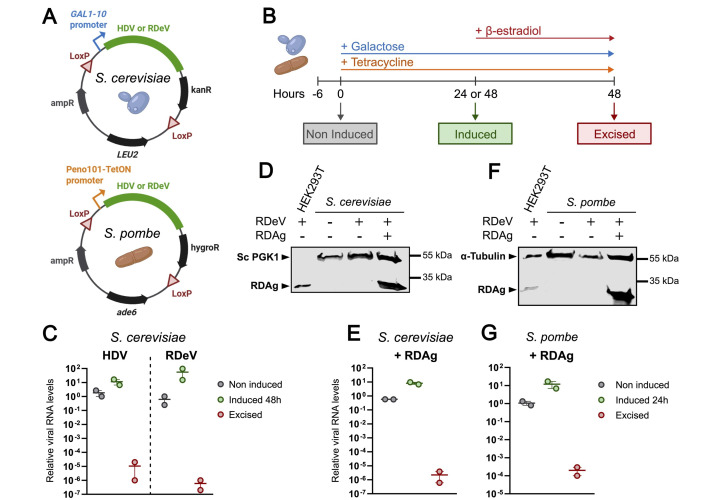 Figure 1
Figure 1|
|
|
|
|
DHP1801
|
|
Lab stock |
|
DHP1802
|
|
This study |
|
DHP1803
|
|
This study |
|
DHP1804
|
|
This study |
|
DHP1805
|
|
Lab stock |
|
DHP1806
|
|
This study |
|
DHP1807
|
|
This study |
|
|
|
|
OKM690 HDV forward |
5’-CTCGGTCAACCTCCTGAGTT-3’ |
|
OKM691 HDV reverse |
5’-AAGGCCCTCGAGAACAAGAA-3’ |
|
OKM678 RDeV forward |
5’-CCATATTTCCAACAGCCGGG-3’ |
|
OKM679 RDeV reverse |
5’-AAGGAGGGAGAGGGGAAAAC-3’ |
|
|
|
|
DHB331 |
pD371-loxP-kanMX6-GAL1-10-loxP-ScADH1term |
|
DHB402 |
pD371-loxP-kanMX6-GAL1-10-RDeV2X-loxP-ScADH1term |
|
DHB403 |
pD371-loxP-kanMX6-GAL1-10-HDV2X-loxP-ScADH1term |
|
DHB332 |
pDB5320-ade6-NotI-CMV-tetR-loxP-enotetSW2-ScADH1term-hphMX6-loxP |
|
DHB404 |
pDB5320-ade6-NotI-CMV-tetR-loxP-enotetSW2-RDAg-ScADH1term-hphMX6-loxP |
|
DHB401 |
pSP403-HIS3-CEN/ARS-TDH3prom-CYC1term |
|
OKM80 |
pSP403-HIS3-CEN/ARS-TDH3prom-RDAg-CYC1term |
|
DHB389 |
pREP1NT-ura4-ars1-nmt1prom |
|
DHB405 |
pREP1NT-ura4-ars1-nmt1prom-RDAg |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Virus Research Studies · Viral Infections and Immunology Research · Animal Disease Management and Epidemiology
Description
Recent years have witnessed the discovery of Hepatitis D virus- like sequences in a wide range of animal species, including rats, bats, snakes, frogs, birds, and termites (Bergner et al., 2021; W.-S. Chang et al., 2019; Hetzel et al., 2019; Iwamoto et al., 2021; Paraskevopoulou et al., 2020; Wille et al., 2018). This led to the establishment of the viral family Kolmioviridae , also known as deltaviruses (Hepojoki et al., 2021). Deltaviruses are the smallest satellite viruses known to infect animals. Their genome is a single-stranded negative-sense circular RNA of approximately 1.7 kb (Pérez-Vargas et al., 2021). The most studied deltavirus, Hepatitis D Virus (HDV), infects humans and causes the most severe form of viral hepatitis (Khalfi et al., 2023). Because these viruses infect a broad range of animal species, their replication depends on highly conserved mechanisms (Lehmann et al., 2007). Indeed, several studies have shown that both HDV and rodent deltavirus (RDeV) are able to replicate in heterologous animal cells (Gnouamozi et al., 2024; Khalfi et al., 2024; Paraskevopoulou et al., 2020). Whether these viruses can replicate in other eukaryotic kingdoms of life has never been reported. To address this question, we tested the ability of HDV and RDeV to replicate in the model organism Saccharomyces cerevisiae . The budding yeast is indeed one of the most well-understood eukaryotic organisms and has already proven useful in virology. Notably, the yeast two-hybrid system was used to characterize HIV-1 protein interactions with host factors (Andréola & Litvak, 2012).
Two copies of the cDNA from either the HDV or RDeV genome were cloned in head-to-tail orientation, as described before (Paraskevopoulou et al., 2020), into plasmids enabling stable genomic integration at the LEU2 gene locus. The viral cDNAs were placed under the control of the GAL1-10 promoter and before an antibiotic selection marker, generating an inducible and selectable expression module, which we flanked with LoxP sites for Cre-mediated excision ( Fig. 1A, B). The obtained plasmids were transformed into a strain stably expressing a β-estradiol-inducible Cre-ER recombinase (Verzijlbergen et al., 2010), allowing controllable excision of the viral sequence through homologous recombination ( Fig. 1B ).
We first analyzed HDV and RDeV RNA levels upon induction and excision of the viral cDNA by RT-qPCR ( Fig. 1C ). Viral RNA levels showed a 10-fold increase upon galactose induction ( Fig. 1C ). Following template excision and clone propagation, viral RNA decreased substantially, to reach the detection limit of qPCR, indicating that viral RNA is unable to replicate and persist in the absence of its cDNA template. The viral RNA detected before induction ( Fig. 1C ) is most likely due to low GAL1-10 promoter activity in raffinose media (non-induced in Fig. 1B ).
The delta-antigen (DAg) is essential for replication and forms viral ribonucleoproteins in association with the RNA genome. DAg proteins are imported into the nuclei of infected cells and recruit host RNA polymerase II to initiate viral replication (J. Chang et al., 2008a; Lehmann et al., 2007). We thus hypothesized that HDV and RDeV might be unable to replicate because the deltavirus strains we used lack DAg, possibly because induced viral RNAs are not processed into a DAg mRNA that is matured for export and translation. To test if exogenous addition of the viral DAg protein triggers viral replication, we transformed a non-integrative plasmid encoding the rodent DAg (RDAg) into the RDeV strains. Western blot analyses showed that RDeV strains lacked RDAg expression, even when RDeV genome expression was induced ( Fig. 1D ), while RDAg was detected in RDeV strains transformed with the RDAg encoding plasmid ( Fig. 1D ). We then monitored RDeV RNA genome levels by RT-qPCR and observed an increase of viral RNA levels upon induction and a decrease after viral cDNA excision, mirroring what we found in RDeV strains lacking the RDAg ( Fig. 1E ). We conclude that RDeV is unable to replicate in S. cerevisiae , even in the presence of exogenous RDAg protein.
Our results suggest that S. cerevisiae lacks one or more host factors essential for HDV and RDeV replication. Schizosaccharomyces pombe is another prime model yeast species that diverged from other Ascomycetes at the root of this clade, about 530 million years ago (Shen et al., 2020). Thus, comparing these two model species can bring complementary information on fundamental biological processes.
To determine whether S. pombe is able to sustain deltavirus replication, the RDeV cDNA was cloned as described before, in a plasmid for inducible expression and controllable excision in S. pombe . For this, we used a recently available vector containing a tetracycline-inducible promoter (Lyu et al., 2024), a constitutively expressed Tet repressor cDNA, and a homologous region for stable integration upstream of the ade6 gene locus. The entire viral expression module was flanked by two LoxP sites for β-estradiol-inducible Cre-mediated excision ( Fig. 1A ). An S. pombe strain stably expressing the Cre-ER recombinase was transformed and analyzed as schematized in Fig. 1B, analogous to S. cerevisiae . Western blot analysis showed that the RDAg was not expressed upon viral genome induction with tetracycline ( Fig. 1F ). We therefore transformed this RDeV strain with a non-integrative plasmid encoding RDAg to directly assess the replication in the presence of the delta-antigen, which expression was confirmed by Western blot ( Fig. 1F ). In this strain, while the levels of RDeV viral RNA increased by 10-fold upon tetracycline induction, they decreased to barely detectable levels upon RDeV cDNA excision ( Fig. 1G ). These results indicate that RDeV cannot replicate in S. pombe , even when the RDAg is present, as in S. cerevisiae ( Fig. 1E ).
In conclusion, our study suggests that despite the broad range of species infected by deltaviruses, their replication appears restricted to animals, suggesting that deltavirus replication requires metazoan-specific co-factors, for example for recruiting or activating the RNA polymerase II machinery on an RNA template (J. Chang et al., 2008b; Lehmann et al., 2007). Although S. cerevisiae RNA polymerase II has RNA-dependent RNA polymerase (RdRP) activity on HDV RNA *in vitro * (Lehmann et al., 2007), our results demonstrate that other factors might be required for viral RNA amplification in vivo . In addition, other essential pro-viral host factors involved in the deltavirus lifecycle may be lacking in Ascomycetes, such as an RNA ligase (Reid & Lazinski, 2000). Another possibility explaining the non-permissiveness of yeasts to deltavirus replication is the expression of antiviral factors that might repress viral replication. For instance, the SKI complex is important for Killer virus attenuation in S. cerevisiae and is present in S. pombe . Similarly, a conserved RNAi machinery exists in S. pombe and might inhibit deltavirus replication in this species, as RNAi has a potent antiviral function in plants and possibly fungi (S.-S. Chang et al., 2012; Ding & Voinnet, 2007). Importantly, our work provides a novel methodological framework for the rigorous assessment of circular RNA virus replication in yeast, through sequential transcriptional induction and recombination-mediated excision of a viral cDNA template. This issue is important because previous studies may have mistakenly interpreted residual viral RNA levels as evidence of autonomous replication, which we show completely disappeared upon loss of the cDNA template for deltaviruses (Delan-Forino et al., 2011; Janda, 1993).
Methods
Cloning:
Human and rodent deltavirus genomes and antigen-coding sequences were amplified from previously described pcDNA3.1 plasmids (Khalfi et al., 2024) and cloned in expression plasmids for S. cerevisiae and S. pombe by Gibson Assembly (New England Biolabs).
S. cerevisiae * and * S. pombe * :
All S. cerevisiae and S. pombe strains are listed in Table 1. S. cerevisiae was grown at 30°C in rich YEPD medium for parental, HDV, and RDeV strains, and in synthetic minimal YNB without histidine for RDAg strains. S. pombe was grown at 32°C in rich YES medium for parental and RDeV strains, and in synthetic minimal EMM without uracil for RDAg strains. Yeasts were transformed using a LiAc-based procedure for chemical competence (Gietz et al., 1992). HDV and RDeV cDNA integration was selected on YNB without leucine or YEPD with G418 for S. cerevisiae and on EMM without adenine or YES with hygromycin for S. pombe . S. cerevisiae harboring RDAg coding plasmids were selected and kept on YNB without histidine, while S. pombe harboring RDAg coding plasmids were selected and kept on EMM without uracil.
Viral genome induction:
Viral genome expression was induced with 2% galactose for S. cerevisiae and with 2.5 µg.mL ^-1^ tetracycline for S. pombe (Lyu et al., 2024). Cre-ER recombinase was activated with 1µM β-estradiol. Excision was validated on a rich medium plate containing G418 or hygromycin for S. cerevisiae and S. pombe , respectively. For each sample, 50 mL of exponentially growing cells were collected, pelleted, and stored at -80°C before RNA or protein extraction.
RT-qPCR:
RNA extraction was performed using hot acidic phenol as described previously (Toullec et al., 2021). and DNase I digestion was used to remove contaminating DNA. 1 µg of RNA was reverse transcribed with SuperScript III (Invitrogen). Fluorescence-based quantitative PCR was performed using SYBR Green on the CFX Opus 384 Real-Time PCR System (Bio-Rad), and relative quantities were calculated from standard curves.
Western blot:
Western blot was then performed as described before (Khalfi et al., 2024), using the same secondary antibodies. S. cerevisiae was incubated in 0.1M NaOH for 5 minutes, pelleted, and resuspended in protein loading dye. Samples were then denatured for 3 minutes at 95°C, pelleted, and the supernatant was used for Western blot. An anti-Pgk1 antibody was used to control for loading (ThermoFisher #459250). RDAg was detected using polyclonal antibodies extracted from patient sera (Khalfi et al., 2024).
For S. pombe , protein extraction was performed using lysis buffer (40 mM HEPES-NaOH pH 7.4, 350 mM NaCl, 0.1% NP40, and 10% glycerol) supplemented with protease inhibitors (cOmplete EDTA-free cocktails tablets, 1 mM PMSF, 1 mg.ml ^-1^ bestatin, and 1 mg.ml ^-1^ pepstatin A). An anti-α-Tubulin antibody was used to control for loading. RDAg was detected using rabbit serum/antiserum immunized with recombinant SDAg (Hetzel et al., 2019).
Reagents
**: **
**: **
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Andréola ML Litvak S 2012617 Yeast and the AIDS virus: the odd couple.J Biomed Biotechnol 20121110-724354902054902010.1155/2012/54902022778552 PMC 3385842 · doi ↗ · pubmed ↗
- 2Bergner LM Orton RJ Broos A Tello C Becker DJ Carrera JE Patel AH Biek R Streicker DG 2021119 Diversification of mammalian deltaviruses by host shifting.Proc Natl Acad Sci U S A 11830027-842410.1073/pnas.201990711833397804 PMC 7826387 · doi ↗ · pubmed ↗
- 3Chang J Nie X Chang HE Han Z Taylor J 20071121 Transcription of hepatitis delta virus RNA by RNA polymerase II.J Virol 8230022-538X 1118112710.1128/JVI.01758-0718032511 PMC 2224410 · doi ↗ · pubmed ↗
- 4Chang SS Zhang Z Liu Y 2012628 RNA interference pathways in fungi: mechanisms and functions.Annu Rev Microbiol 660066-422730532310.1146/annurev-micro-092611-15013822746336 PMC 4617789 · doi ↗ · pubmed ↗
- 5Chang WS Pettersson JH Le Lay C Shi M Lo N Wille M Eden JS Holmes EC 2019715 Novel hepatitis D-like agents in vertebrates and invertebrates.Virus Evol 522057-1577 vez 021vez 02110.1093/ve/vez 02131321078 PMC 6628682 · doi ↗ · pubmed ↗
- 6Delan-Forino C Maurel MC Torchet C 2011126 Replication of avocado sunblotch viroid in the yeast Saccharomyces cerevisiae.J Virol 8570022-538X 3229323810.1128/JVI.01320-1021270165 PMC 3067865 · doi ↗ · pubmed ↗
- 7Ding SW Voinnet O 2007810 Antiviral immunity directed by small RN As.Cell 13030092-867441342610.1016/j.cell.2007.07.03917693253 PMC 2703654 · doi ↗ · pubmed ↗
- 8Gietz D St Jean A Woods RA Schiestl RH 1992325 Improved method for high efficiency transformation of intact yeast cells.Nucleic Acids Res 2060305-10481425142510.1093/nar/20.6.14251561104 PMC 312198 · doi ↗ · pubmed ↗