Isolation and molecular characterization of subgroup J avian leukosis virus in native chicken breeds of China during 2022–2025
Qi Liu, Sifan Ji, Minghui Li, Lei He, Ke Ding, Zuhua Yu, Jian Chen

TL;DR
This study characterizes ALV-J virus strains isolated from native Chinese chickens between 2022 and 2025, revealing mutations that may enhance their replication and pathogenicity.
Contribution
The study identifies novel mutations in ALV-J strains from native Chinese chickens, offering insights into their genetic evolution and potential pathogenicity.
Findings
Nine ALV-J isolates clustered in Clades 1.2 and 1.3, indicating concurrent prevalence in multiple native chicken breeds.
Key mutations in Gp85 and 3′ UTR suggest enhanced replication and pathogenicity of the isolates.
Variations in α-helices in Gp85 proteins may affect virus-host interactions.
Abstract
Avian leukosis virus subgroup J (ALV-J) primarily infects poultry, especially chickens, where it induces immunosuppression and tumorigenesis. ALV-J has caused substantial economic losses worldwide and is prevalent among indigenous chicken breeds in China. In this study, we analyzed the genomic characteristics of ALV-J strains isolated from diseased liver tissue or anticoagulant blood samples collected from Lushi chickens, Central Plains cockfighting, and Hetian chickens between 2022 and 2025. The results showed that the nine isolates clustered within Clades 1.2 and 1.3, indicating that ALV-J is concurrently prevalent in multiple native chicken lineages. Compared with the ALV-J prototype strain HPRS-103, multiple specific functionally significant point mutations or deletion mutations occurred in the Gp85 protein and the 3′ untranslated region (3′ UTR) of all the isolates. These included…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
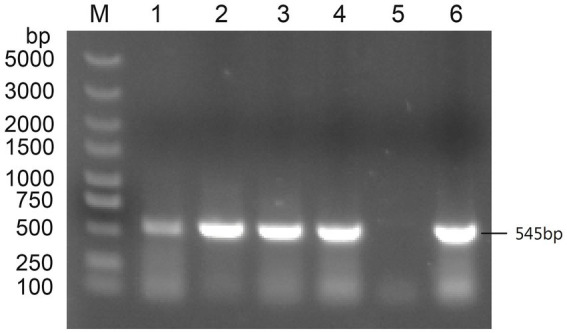 Figure 1
Figure 1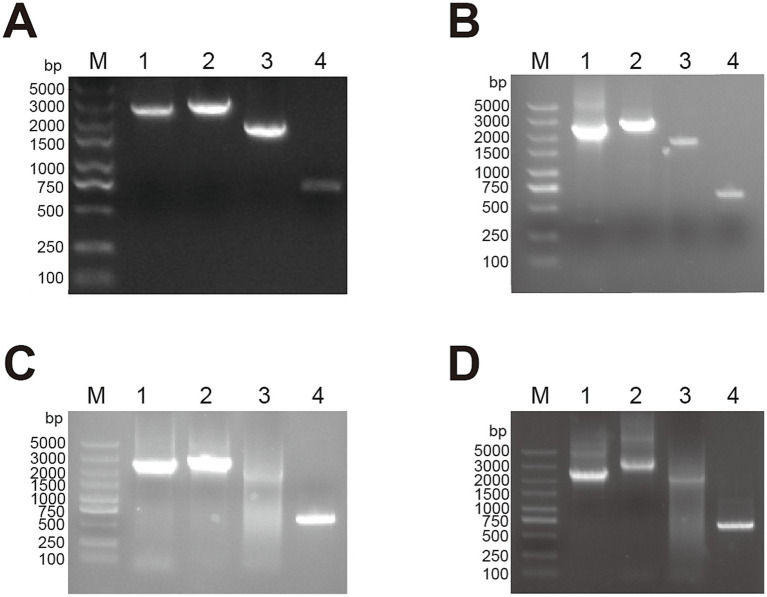 Figure 2
Figure 2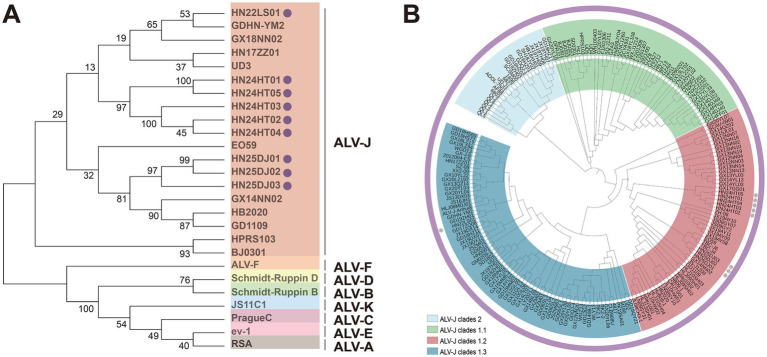 Figure 3
Figure 3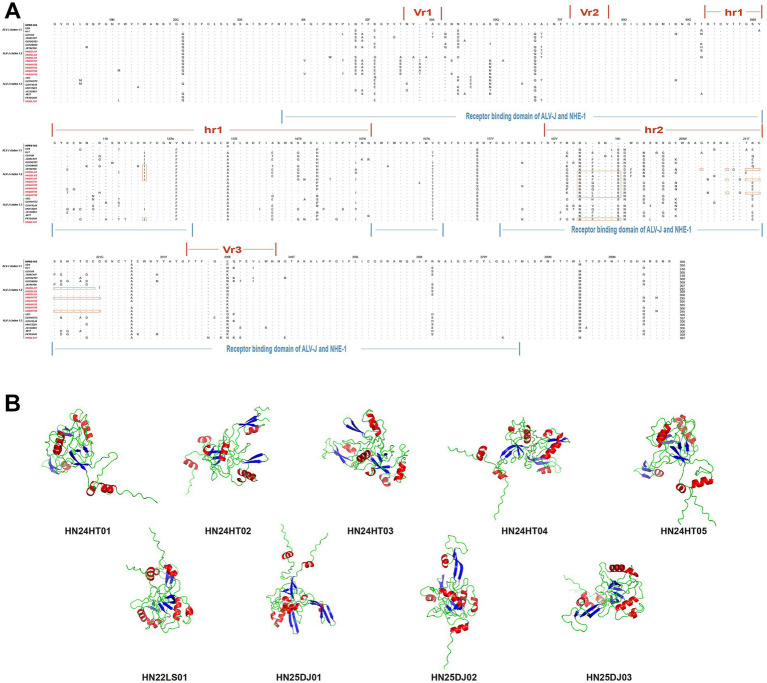 Figure 4
Figure 4 Figure 5
Figure 5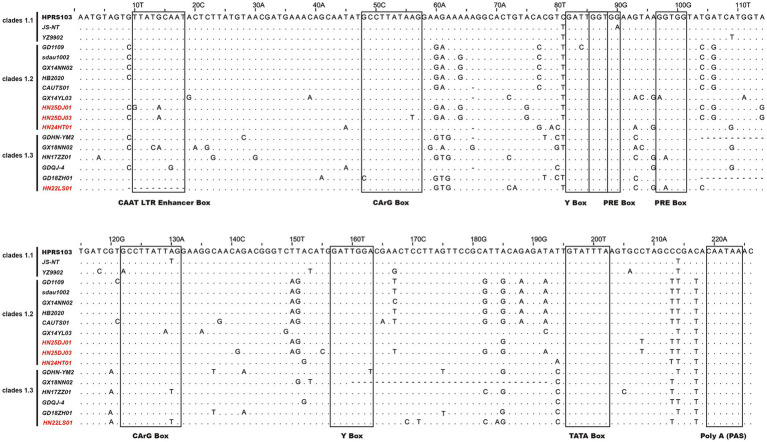 Figure 6
Figure 6| Primer | Sequences(5′−3′) | PCR product size |
|---|---|---|
| J545-F | GGATGAGGTGACTAAGAAAG | 545 bp |
| J545-R | CGAACCAAAGGTAACACACG | |
| S1-F | TGTAGTGTTATGCAATACTCTTATG | 2,457 bp |
| S1-R | CCTCCTCTGAAATAATAGTGATG | |
| S2-F | CATCACTATTATTTCAGAGGAGG | 2,806 bp |
| S2-R | CCTCATCTTTCTTAGTCACC | |
| S3-F | GATGAGGCGAGCCCTCTCTTTG | 2,200 bp |
| S3-R | TGTGGTGGGAGGTAAAATGGCGT | |
| S4-F | GAATTAGGGAGCAGCTGTAG | 648 bp |
| S4-R | TGAAGCCTTCTGCTTCATTCAGG | |
| A-F | GGATGAGGTGACTAAGAAAG | 694 bp |
| A-R | AGAGAAAGAGGGGYGTCTAAGGAGA | |
| K-F | TCCAGGCCGCAACTCAC | 1,214 bp |
| K-R | CATACCACCACCCACGTACT | |
| B-F | CGAGAGTGGCTCGCGAGATGG | 1,100 bp |
| B-R | AGCCGGACTATCGTATGGGGTAA |
| Strains | Origin | Year | Host | Accession no. |
|---|---|---|---|---|
| HN22LS01 | Henan | 2022 | Lushi chicken | |
| HN24HT01 | Fujian | 2023 | Hetian chicken | |
| HN24HT02 | Fujian | 2023 | Hetian chicken | |
| HN24HT03 | Fujian | 2023 | Hetian chicken | |
| HN24HT04 | Fujian | 2023 | Hetian chicken | |
| HN24HT05 | Fujian | 2023 | Hetian chicken | |
| HN25DJ01 | Henan | 2025 | Cockfighting | |
| HN25DJ02 | Henan | 2025 | Cockfighting | |
| HN25DJ03 | Henan | 2025 | Cockfighting |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHerpesvirus Infections and Treatments · RNA and protein synthesis mechanisms · interferon and immune responses
Introduction
1
Avian leukosis virus (ALV) is an oncogenic and immunosuppressive retrovirus that causes reduced fertility, growth retardation, and increased susceptibility to secondary infections in poultry (Payne and Nair, 2012; Feng et al., 2017). Since its discovery, ALV has caused substantial economic losses globally. Although ALVs have been nearly eradicated from commercial chicken breeds in Western countries, they continue to circulate in many indigenous Chinese chicken breeds and pose a latent threat to other regions as potentially pathogenic agents (Malhotra et al., 2015; Zheng et al., 2022; Deng et al., 2022). ALVs are classified into 11 subgroups (A–K) according to host range, Gp85 (SU) envelope glycoprotein sequence, and overall genomic characteristics. Endogenous ALV is mainly represented by ALV-E, which is nonpathogenic. Among the exogenous ALVs, ALV-J, ALV-K, and ALV-A are the main prevalent subgroups (Wang et al., 2025; Zhang et al., 2024; Zhang et al., 2025).
ALV is an Alpha-Retrovirus, with a genome of approximately 7.2 ~ 7.8 Kb in size and a structure configured as 5′-LTR-gag-pol-env-LTR-3′ (Fandiño et al., 2023). The gag gene encodes internal proteins, including matrix protein (MA), p10, nucleocapsid protein (NC), protease (PRO), as well as an antigenic capsid protein p27 (CA) that is common to all subgroups of ALV (Freick et al., 2022). The pol gene encodes the proteins required for the replication of ALV (Su et al., 2018). The env gene encodes the envelope protein, which can be divided into two parts: the surface protein Gp85 and the transmembrane protein Gp37 (TM) (Fandiño et al., 2023). The Gp37 anchors the Gp85 to the viral envelope, enabling the proteins to be correctly positioned and facilitating their roles in viral infection and membrane fusion (Mothes et al., 2000). The host range regions 1 and 2 (hr1 and hr2) within Gp85 primarily determine the receptor specificity of ALVs, while the variable regions 1, 2, and 3 (vr1, vr2, and vr3) represent secondary sites of variation among ALV strains, notably, vr3 contributes to the specificity of receptor interactions (Chen et al., 2022; Chen et al., 2020). The long terminal repeat (LTR) plays a critical role in viral oncogenesis by activating host proto-oncogenes such as c-myc (Fung et al., 1983). It consists of three functionally distinct regions: U3, which contains enhancer elements and a promoter, R, which marks the transcription start site, and U5, which possesses post-translational regulatory elements and contribute to polyadenylation (Böhnlein et al., 1989; Ruddell, 1995). In contrast to the conserved U5 region, the U3 region exhibits considerable variability.
Avian leukosis virus subgroup J (ALV-J) is the causative agent of myeloid leukosis and was first identified in broiler chickens in England in 1988 (Payne et al., 1991). Subsequently, ALV-J was first isolated from broiler flocks in China in 1999, after which it rapidly spread to layer chickens and various indigenous breeds (Du and Ai, 1999; Xu et al., 2004; Sun and Cui, 2007). ALV-J infection leads to persistent high-level viremia in broiler chickens throughout their lifespan (Fadly and Smith, 1999). Due to the extremely high rates of vertical and horizontal transmission, ALV-J exhibits substantial genetic and antigenic variations among isolates, which complicates efforts at eradication (Meng et al., 2022; Li et al., 2017; Liao et al., 2022; Liu et al., 2021). The C terminus of Gp37 plays a crucial role in the pathogenesis of ALV-J (Li et al., 2020). The Gp85 of ALV-J exhibits high variability and undergoes rapid adaptive evolution under the selective pressure of host resistance induced by biotechnological approaches, which poses a significant challenge to develop ALV-J resistance through gene editing (Meng et al., 2016; Matoušková et al., 2024). Based on the genetic sequence variations in gp85, ALV-J can be further classified into Clades 1.1, 1.2, 1.3, 2, and 3 (Deng et al., 2022).
The 3′ untranslated region (3′ UTR) of ALV-J is one of the principal regions for viral mutation and evolution, and it also serves as a crucial component in regulating the viral life cycle and pathogenicity. The 3′ UTR of the ALV-J prototype strain HPRS103 is composed of four regions: r-TM, DR-1, E element and U3. Compared with HPRS103, ALV-J strains isolated after 1988 showed varying degrees of deletions in the 3′ UTR. However, the functions of the ALV-J 3′ UTR are not yet fully understood. Current studies have shown that r-TM is associated with the pathogenicity of ALV-J in laying hens, and its deletion significantly enhances the replication ability of ALV-J strains (Wang et al., 2012; Xu et al., 2023). DR-1 is essential for the export of viral mRNA from the nucleus (Ogert et al., 1996). The E element is closely associated with the virus’s oncogenicity and replication in poultry (Chesters et al., 2006).
In this study, nine strains of ALV-J were isolated from Lushi chicken and Central Plains Cockfighting in Henan province and Hetian chicken in Fujian province, China. The strains were designated HN22LS01, HN24HT01, HN24HT02, HN24HT03, HN24HT04, HN24HT05, HN25DJ01, HN25DJ02, and HN25DJ03, and sequence homology analysis, phylogenetic tree analysis, characteristic mutation analysis and protei structure prediction were carried out on the gp85 gene, and characteristic mutation analysis was performed on the 3′UTR and the LTRU3 region.
Methods and materials
2
Clinical samples
2.1
During the period from 2022 to 2025, samples were collected from three farms in China. Specifically, the deceased Lushi chicken from a breeding company with a scale of 20,000 chickens in Lushi County, Henan Province, was found to have an enlarged liver with grayish-white necrotic spots after post-morexamination; anticoagulant blood samples were collected from Hetian chickens at a breeding company with a stock of 30,000 breeding chickens in Fujian Province. Virus isolation and detection using DF-1 cells showed that some samples were positive for ELISA p27; 2 batches of liver samples of deceased fighting chickens were sourced from an individual breeder with a breeding scale of about 100 chickens in Shangqiu, Henan Province. And post-morexamination revealed that the chickens’ livers were abnormally enlarged and filled the abdominal cavity.
Virus isolation and proviral DNA extraction
2.2
DF-1 cells were cultured in DMEM (Servicebio, Wuhan, China) supplemented with 10% FBS (Clark Bioscience, Shanghai, China), and 2% penicillin–streptomycin at 37 °C under 5% CO₂. Liver tissue (0.1 g) was homogenized and centrifuged at 3,000 rpm for 10 min at 4 °C. The supernatant was filtered through a 0.22 μm filter. Serum samples were processed in the same manner. Filtered supernatants and sera were inoculated onto DF-1 monolayers. After 2 h incubation at 37 °C, the medium was replaced with DMEM containing 1% FBS. Infected cells were cultured for 7 days. After infection, cells underwent one freeze–thaw cycle (−80 °C to 37 °C), were centrifuged at 2,000 rpm for 10 min at 4 °C, and genomic DNA was extracted from the pellet using a commercial kit according to the manufacturer’s instructions.
PCR and sequencing
2.3
PCR amplification was performed using ALV-A-specific primers, yielding a 694 bp product for ALV-A detection. PCR amplification was performed using ALV-B-specific primers, yielding a 1,100 bp product for ALV-B detection. PCR amplification was performed using ALV-K-specific primers, yielding a 1,214 bp product for ALV-K detection. PCR amplification was performed using ALV-J-specific primers, yielding a 545 bp product for ALV-J detection. Four primer pairs covering the entire ALV-J genome were designed for whole-genome amplification (Table 1). The 648-bp PCR product was amplified using 2 × Taq Master Mix (Vazyme, Nanjing, China), while high-fidelity DNA polymerases were used for the remaining three genomic fragments. The thermal cycling conditions for the 648-bp fragment were: initial denaturation at 95 °C for 5 min; followed by 35 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 40 s; with a final extension at 72 °C for 8 min. The cycling parameters for the other three genomic regions were: initial denaturation at 95 °C for 3 min; followed by 30 cycles of denaturation at 95 °C for 15 s, annealing at 60 °C for 15 s, and extension at 72 °C for 2 min. The amplification products were subjected to 1% agarose gel electrophoresis and visualized using a gel imaging system. The amplification products were recovered using the StarPrep DNA Gel Extraction Kit (GenStar, Beijing, China), and then ligated into the pMD-19 T vector. Individual clones were selected and subjected to gene sequencing by Beijing Tsingke Biotechnology Co., Ltd. The env genes of HN25DJ02, HN24HT02, HN24HT03, HN24HT04, and HN24HT05 were successfully sequenced. Subsequently, the whole-genome assembly of HN25DJ01, HN25DJ03, HN22LS01, and HN24HT01 was completed using the DNASTAR SeqMan software.
Sequence alignment and phylogenetic analysis
2.4
Sequence alignment and homology analysis were conducted using the Clutal W Method in DNASTAR MegAlign. Phylogenetic analysis was performed with the neighbor-joining method in Molecular Evolutionary Genetics Analysis version 12 (MEGA 12), with 1,000 bootstrap replicates. All the reference sequences were obtained from GenBank and shown in Supplementary Table S1.
Secondary structure prediction and analysis of ALV-J Gp85 protein
2.5
The secondary structure of ALV-J Gp85 protein was predicted by AlphaFold 3,1 and the software PyMOL was used for visual analysis of the protein structure (Abramson et al., 2024).
Nucleotide sequence accession numbers
2.6
The sequences obtained in this study have been submitted to GenBank, and the accession numbers are provided in Table 2.
Results
3
Isolation and identification of ALV-J strains
3.1
Subgroup-specific detection was performed on the clinical samples submitted by various farms to detect AIV. The ALV-J specific bands of approximately 545 bp were successfully amplified (Figure 1). When amplified using the primers specific for ALV-A and ALV-B, no bands were amplified from all the samples. When amplified using the primers specific for ALV-K, no bands were amplified from the samples of cockfighting. However, the samples of Lushi chickens and Hetian chickens were amplified to yield specific bands with a length of 1,214 bp (Supplementary Figure S1). Subsequently, the tissue homogenates or sera that were detected as ALV-J positive were inoculated into DF-1 cells for virus isolation and identification. ELISA results showed that the DF-1 cells inoculated with PCR-positive samples were all ALV-positive. Four pairs of primers were used to perform PCR amplification on four ALV-J isolates. The results showed that all four ALV-J isolates successfully amplified the four target fragments (Figure 2). The amplified fragments were ligated into the pMD-19T vector, sequenced, and assembled to obtain the complete genomic sequences of the four ALV-J isolates, which were named HN22LS01, HN24HT01, HN25DJ01, and HN25DJ03, with genome lengths of 7,609, 7,582, 7,609, and 7,645 nt, respectively. Additionally, five ALV-J isolates were obtained. These five isolates were named HN24HT02, HN24HT03, HN24HT04, HN24HT05, and HN25DJ02 respectively, and their env genes were also obtained at the same time, with env sequence lengths of 1,655, 1,649, 1,652, 1,653, and 1,683 nt, respectively. All obtained sequences were consistent with the characteristic features of ALV.
The PCR result of primers specific targeting ALV-J. M: Maker, 1: DNA of liver samples from Lushi chickens, 2: Isolate serum from anticoagulated blood to infect DF-1 cells, 3: DNA of liver samples from the first batch of cockfighting, 4: DNA of liver samples from the second batch of cockfighting, 5: Negative control, 6: Positive control.
The PCR result of full-length primers specific for ALV-J. M: Maker, 1: 2,457 bp amplification product, 2: 2,806 bp amplification product, 3: 2,200 bp amplification product, 4: 648 bp amplification product. (A) HN22LS01 (B) HN24HT01 (C) HN25DJ01 (D) HN25DJ03.
Phylogenetic and homology analysis the ALV-J isolates
3.2
The full-lengths genomes of the four ALV-J isolates ranged from 7,582 to 7,645 bp. Compared with the whole genome sequences of the reference strains, the sequence homology ranged from 93.7 ~ 95.9%. Compared with the ALV-J strains that have been published in GenBank, the pol gene sequences of the four ALV-J strains with full-length genomes were highly conserved, with a homology ranging from 96.2 ~ 98.9%. The homology of the gag gene was from 93.6 ~ 96.4%, and that of the LTR was from 84.8 ~ 95.7%. Compared with the ALV-J strains published in GenBank, the env gene homology of the 9 ALV-J strains was from 91.1% ~ 96.5%, the gp85 gene homology was from 85.5 ~ 96.8%, and the gp37 gene homology was from 91.9% ~ 97.9%.
Phylogenetic analysis of ALV-J env demonstrated that the env genes of these nine isolates clustered in the same branch as the ALV-J reference strains, while other subgroups were grouped into different evolutionary branches (Figure 3A). These data clearly showed that these nine isolates belonged to ALV-J. Further phylogenetic analysis of gp85 indicated that 8 ALV-J isolates derived from cockfighting and Hetian chickens clustered into the ALV-J Clades 1.2 branch, and the 1 ALV-J isolate from Lushi chickens clustered into the ALV-J clades 1.3 branch (Figure 3B). The isolates HN25DJ01, HN25DJ02, and HN25DJ03 isolated from cockfighting had a very close genetic distance to the Jiangsu isolates TBC-J4 and TBC-J6 as well as the Nigeria isolate EO59. The isolates HN24HT01, HN24HT02, HN24HT03, HN24HT04, and HN24HT05 isolated from Hetian chickens had a very close genetic distance to the Guangxi isolate GX17GG01 isolated in 2017. The isolate HN22LS01 isolated from Lushi chickens had a very close genetic distance to the Guangdong isolates GD19ZH01 and GDHN - YM2.
Phylogenetic analysis of ALV-J isolates. (A) Phylogenetic analysis for the Env protein of ALV-J isolates. Solid cycle (●) indicates ALV isolates in this study. The Neighbor-Joining method with 1,000 bootstrap replicates was used to infer the evolutionary relationships (MEGA 12). (B) Phylogenetic analysis for the Gp85 protein of ALV-J isolates. Solid cycle (●) indicates ALV isolates in this study. The Neighbor-Joining method with 1,000 bootstrap replicates was used to infer the evolutionary relationships (MEGA 12).
Characteristics of the Gp85 subunit of the isolates
3.3
When comparing the Gp85 amino acid sequences of the isolated strains with those of the ALV-J reference strain, it was found that the mutations in all isolated strains mainly occurred in vr1, hr1, hr2, vr2, and vr3 (Figure 4A). Remarkable deletion mutations within the receptor-binding domain (RBD) were observed in HN25DJ01, HN24HT01, and HN24HT05, and these mutations partially overlapped with hr2. Specifically, compared with the ALV-J prototype strain HPRS-103, the deletion mutations of HN25DJ01 occurred at amino acid positions 206 and from 210 to 220, while those of HN24HT01 and HN24HT05 were located from position 210 to 221. Moreover, the N123I mutation in HN25DJ01, HN25DJ02, HN25DJ03, HN24HT01, and HN22LS01 enhanced the viral replication by increased Gp85 stability and an expanded interaction interface. Notably, the D191N mutation observed in all isolated strains compared with HPRS-103 suggested the emergence of a novel N-glycosylation site in the Env protein. Furthermore, using structural predictions generated by AlphaFold 3, we conducted a comparative analysis of the Gp85 proteins of nine ALV-J isolated strains to analyze the differences in their secondary structures. As depicted in Figure 4B, all isolated strains consistently contained nine β-sheet structures, whereas the number of α-helices varied: HN25DJ01 possessed 9 α-helix structures, HN25DJ03 and HN22LS01 had 8 α-helix structures, and HN25DJ02, HN24HT01, HN24HT02, HN24HT03, HN24HT04, and HN24HT05 each contained 7 α-helix structures.
Amino acid sequence analysis and secondary structure prediction of ALV-J Gp85 protein. (A) Comparison of the amino acid sequences of Gp85 from the 9 ALV-J isolates and the ALV-J reference strains. The inferred Gp85 was compared to other ALV strains. The colored underlines represent the hr1, hr2, vr1, vr2, and vr3 regions and the receptor binding domain of ALV-J Gp85. The letters indicate amino acid substitutions, dots (.) indicate identical amino acid, and dashes (−) indicate gaps produced in the alignment. The 9 ALV-J isolates were marked in red. (B) Secondary structure of Gp85 protein analyzed using AlphaFold3. Red represents the alpha helix, and blue represents the beta sheet.
Characteristics of the 3′ UTR of the isolates
3.4
Genetic variations in the rTM, DR-1 and E elements within the 3′ UTR of ALV-J can influence the pathogenicity, oncogenicity and replication capacity. As shown in the Figure 5, the genetic variations within the 3′ UTR of the four isolated strains were relatively consistent. All of them retained the intact DR-1 and E elements and showed highest similarity to the reference strains GD1109, HB2020, CAUTS01 and sadu1002. The four isolated strains exhibited varying degrees of deletions in the r-TM region. HN25DJ01 and HN25DJ03 each had a deletion of 176 base pairs in the r-TM sequence, while HN24HT01 and HN22LS01 had a deletion of 210 base pairs in the r-TM sequence.
Comparison of the important regulatory elements of the 3′UTR regions of the isolates. The 4 ALV-J isolates were marked in red.
Transcription factor analysis of 3′LTR
3.5
Analysis of the LTR U3 region sequences and their transcription factor elements revealed that, compared with the 226 bp U3 region of the HPRS103 strain, the HN24HT01 isolate exhibited a 1 bp deletion at position 66, while the HN22LS01 isolate harbored a 10 bp deletion spanning positions 9 ~ 18 (Figure 6). Except for HN22LS01, which lacked one CAAT enhancer, all isolates contained two CArG boxes, Y boxes, PRE motifs, as well as a single CAAT enhancer, C/EBP box, and TATA box.
Analysis of the nucleotide sequences in the U3 region of ALV-J isolates. Letters indicate nucleotide substitutions; dots represent identical nucleotides; dashed lines indicate gaps produced in the alignment. The putative transcriptional regulatory elements are represented by black boxes.
Discussion
4
Among the known ALV subgroups, ALV-J represents the greatest ongoing threat to China’s poultry industry due to its heightened pathogenicity and transmissibility (Liang et al., 2019; Tan et al., 2024). Since 2008, outbreaks of this virus have occurred in China’s layer chicken flocks and spread rapidly among various chicken breeds in China. To date, it has not been completely eradicated (Gao et al., 2012; Pan et al., 2012; Lin et al., 2016; Yang et al., 2025). This situation is largely because of its continuous evolution, which generates novel mutations affecting viral infection, replication, and pathogenicity (Cui et al., 2003; Zhang et al., 2025). Therefore, ongoing surveillance of the prevalent strains ALV-J and analysis of their molecular biological characteristics are of vital importance for the prevention and control of ALV in China and for the sustainable development of the poultry industry.
In this study, we successfully isolated and identified 9 strains of ALV-J (HN22LS01, HN24HT01, HN24HT02, HN24HT03, HN24HT04, HN24HT05, HN24DJ01, HN24DJ02, HN24DJ03) from native chickens in China. Compared with the ALV-J strains that have been published in GenBank, the homology of the gp85 gene of the 9 strains of ALV-J was between 85.5 and 96.8%. Phylogenetic analysis of the Gp85 protein showed that all the isolated strains belonged to ALV-J Clade 1.2 and Clade 1.3. Consistent with the research results in recent years, compared with the initial ALV-J Clade 1.1 strains that entered China, Clade 1.2 and Clade 1.3 demonstrated a higher prevalence (Deng et al., 2021; Deng et al., 2022). The ALV-J isolates from cockfighting exhibited high homology with the isolates TBC-J4 and TBC-J6 from Jiangsu, as well as EO59 from Nigeria in Africa. The ALV-J isolates from Hetian chickens showed high homology with the isolate GX17GG01 from Guangxi. The ALV-J isolate from Lushi chickens were highly homologous to the isolates GDHN-YM2 and GD19ZH01 from Guangdong. The high homology between the Chinese isolates and the African isolates implies that ALV-J might have achieved cross-regional transmission through channels like transnational live chicken trade (Deng et al., 2022). This will help trace the global diffusion trajectory of ALV-J. Based on the homology, global monitoring can focus on the shared mutation features and collaboratively adjust prevention and control strategies.
Through in-depth analysis of the sequence and structural characteristics of the Gp85 protein, we found that significant differences existed in the functional domains and secondary structures among different isolates. Previous studies have confirmed that the two discontinuous amino acid sequences, namely positions 38–131 and 159–283 of the ALV-J Gp85 protein, constitute the RBD of Gp85 (Zhang et al., 2020). Continuous deletion mutations within the RBD of HN25DJ01 occurred at the amino acid positions from 210 to 220, while the deletion mutations of HN24HT01 and HN24HT05 were located at the amino acid positions from 210 to 221, potentially changing the efficiency of the binding between the virus and the host cell receptors and further impacting their replications (Federspiel, 2019). In addition, the N123I mutation occurring in HN25DJ01, HN25DJ02, HN25DJ03, HN24HT01, and HN22LS01 can enhance the replication ability of the virus by strengthening the stability of Gp85 and expanding the interaction interface (Yu et al., 2024). Mutations occurring at the 191st amino acid position in all isolated strains might lead to the formation of a new glycosylation site “NWS,” which could potentially result in changes in the epitope pattern of the env protein as well as in the replication of the virus (Xu et al., 2022). Secondary structure prediction analysis reveals that the number of α-helices varies among different isolates, which might have an impact on the interaction between the virus and its host (Guarracino et al., 2019). It also indicates that the Gp85 protein located on the surface of the ALV-J virion may enhance its adaptability to the host through conformational remodeling (Wang et al., 2025).
Through its three important elements (DR-1, E element, and r-TM), the 3′ UTR of ALV-J can influence the tissue preference of the virus and act as an enhancer to promote viral replication (Xu et al., 2023). Intriguingly, the ALV-J strain containing △-r-TM-type 3′ UTR exhibited stronger replication capacity and pathogenicity (Xu et al., 2023; Wang et al., 2012). Meanwhile, the integrity of the E element further enhances the replicative capacity of the virus (Cao et al., 2024). This type of 3′ UTR endows the virus with the strongest competitive advantage during the evolutionary process (Xu et al., 2023). In this study, the analysis of the 3′ UTR revealed that 4 ALV-J isolates (HN22LS01, HN24HT01, HN24DJ01, HN24DJ03) were similar to the majority of ALV-J isolates in that they all had deletions at r-TM while retaining the integrity of the E element. Therefore, compared with ALV-J with other types of 3′ UTR, we hypothesize that these four ALV-J isolates may possess enhanced replication and pathogenicity, which awaits further verification.
The U3 region contains promoters and enhancers, which are closely related to viral replication, transcription and translation, and are also involved in the activation of host proto-oncogenes (Zachow and Conklin, 1992; Cullen et al., 1985). In this study, a unique 10 bp deletion was observed in the U3 region of the HN22LS01 ALV-J isolate. This mutation led to the loss of the CAAT enhancer box transcription factor, which was not detected in the other three isolates. The CAAT enhancer box can enhance the replication ability of ALV-J (Gao et al., 2015). Therefore, compared with other 3 isolates, it is hypothesized that HN22LS01 may have a weaker replication ability.
In conclusion, our study isolated and characterized nine ALV-J strains from three different breeds of native chickens in China during the period from 2022 to 2025, and analyzed the molecular biological characteristics. The observed structural and genetic changes, particularly in gp85 and the 3′UTR, indicate that these viruses may have evolved enhanced replication capacity and pathogenicity, which are closely related to the genetic evolution of ALV-J. In terms of disease prevention and control, ALV-J with enhanced replication capacity increases the difficulty of prevention and control. Its stronger pathogenicity causes more severe poultry diseases and enables rapid virus spread, so monitoring needs to be strengthened using more sensitive detection methods, and stricter biosecurity measures must be adopted (Cheng et al., 2023). In terms of vaccination, the possible contamination caused by the ALV-J with stronger replication capacity and pathogenicity during the production of commercial live vaccines may lead to immunization failure and greater losses (Wang et al., 2021). In terms of breeding programs, developing individuals with genetic resistance to ALV-J requires complex gene editing. Meanwhile, comprehensive analyses must be conducted to assess the potential impact of gene editing on ALV-J evolution (Matoušková et al., 2024). In the future, continuous monitoring of ALV-J variation, investigation of its epidemiology, and efforts to control and eradicate ALV-J strains in China will be essential.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abramson J.Adler J.Dunger J. (2024). Accurate structure prediction of biomolecular interactions with Alpha Fold 3. Nature. 630, 493–500. doi: 10.1038/s 41586-024-07487-w, PMID: 38718835 PMC 11168924 · doi ↗ · pubmed ↗
- 2Böhnlein S.Hauber J.Cullen B. R. (1989). Identification of a U 5-specific sequence required for efficient polyadenylation within the human immunodeficiency virus long terminal repeat. J. Virol. 63, 421–424. doi: 10.1128/JVI.63.1.421-424.1989, PMID: 2908926 PMC 247699 · doi ↗ · pubmed ↗
- 3Cao Y.Ren Q.Chang S.Cui W.Zhao P.Wang Y. (2024). N 6-methyladenosine RNA methylation modification regulates the transcription of viral-derived E (XSR) mi RN As to promote ALV-J replication. Vet. Microbiol. 298:110218. doi: 10.1016/j.vetmic.2024.11021839159504 · doi ↗ · pubmed ↗
- 4Chen J.Li J.Dong X.Liao M.Cao W. (2022). The key amino acid sites 199-205, 269, 319, 321 and 324 of ALV-K env contribute to the weaker replication capacity of ALV-K than ALV-A. Retrovirology 19:19. doi: 10.1186/s 12977-022-00598-0, PMID: 36002842 PMC 9400301 · doi ↗ · pubmed ↗
- 5Chen X.Wang H.Fang X.Gao K.Fang C.Gu Y.. (2020). Identification of a novel epitope specific for Gp 85 protein of avian leukosis virus subgroup K. Vet. Immunol. Immunopathol. 230:110143. doi: 10.1016/j.vetimm.2020.110143, PMID: 33129191 · doi ↗ · pubmed ↗
- 6Cheng X.Yang J.Bi X.Yang Q.Zhou D.Zhang S.. (2023). Molecular characteristics and pathogenicity of a Tibet-origin mutant avian leukosis virus subgroup j isolated from Tibetan chickens in China. Infect. Genet. Evol. 109:105415. doi: 10.1016/j.meegid.2023.10541536775048 · doi ↗ · pubmed ↗
- 7Chesters P. M.Smith L. P.Nair V. (2006). E (XSR) element contributes to the oncogenicity of avian leukosis virus (subgroup J). J. Gen. Virol. 87, 2685–2692. doi: 10.1099/vir.0.81884-0, PMID: 16894209 · doi ↗ · pubmed ↗
- 8Cui Z.Du Y.Zhang Z.Silva R. F. (2003). Comparison of Chinese field strains of avian leukosis subgroup J viruses with prototype strain HPRS-103 and United States strains. Avian Dis. 47, 1321–1330. doi: 10.1637/6085, PMID: 14708978 · doi ↗ · pubmed ↗