Leptochilus dolichophyllus (Polypodiaceae), a New Hemiepiphytic Species From Fujian, China
Hou‐Hua Fu, Cheng‐Yuan Zhou, Xiong‐De Tu, Liang Ma, Shi‐Pin Chen, Hong‐Jin Wei

TL;DR
Scientists discovered a new fern species in Fujian, China, and confirmed it with both physical traits and genetic analysis.
Contribution
The discovery and molecular characterization of a new hemiepiphytic Leptochilus species from Fujian, China.
Findings
Leptochilus dolichophyllus is distinct from L. leveillei due to its hemiepiphytic habit and longer fronds.
Molecular analysis of three plastid regions confirms L. dolichophyllus as a distinct and well-supported clade.
The complete plastid genome of L. dolichophyllus has been assembled and annotated.
Abstract
A new species, Leptochilus dolichophyllus (Polypodiaceae), from central‐northern Fujian, southeastern China, is described and illustrated. It is morphologically similar to Leptochilus leveillei , but differs by its hemiepiphytic habit and longer frond. Molecular phylogenetic analysis based on three plastid regions (rbcL, rps4 + rps4‐trnS, and trnL + trnL‐trnF) demonstrates that the new species represents a distinct and well‐supported clade. Both morphological and molecular data provide compelling evidence for the recognition of L. dolichophyllus as a new species. Furthermore, the complete plastid genome of this new species is reported. We have identified one new Leptochilus species based on morphological observation and molecular evidence, Leptochilus dolichophyllus H.H.Fu & H.J.Wei, from central‐northern Fujian, southeastern China. Additionally, to provide valuable molecular…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
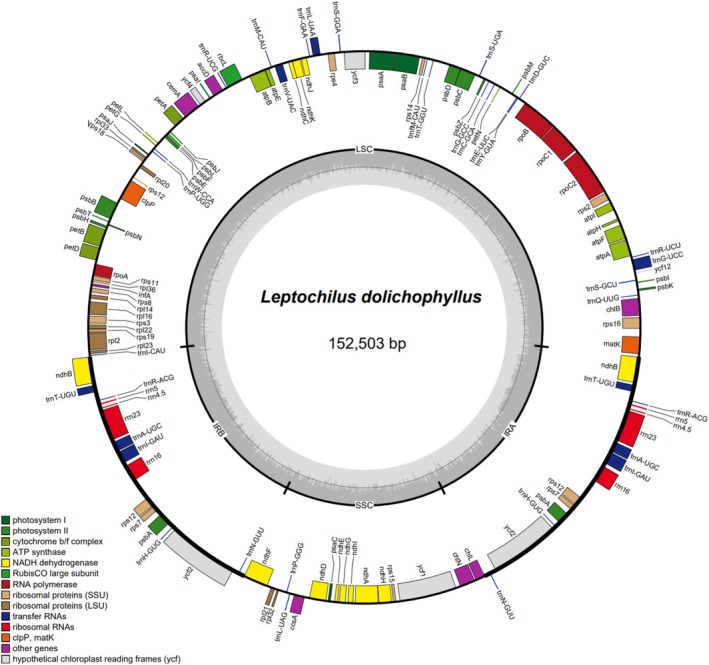 FIGURE 1
FIGURE 1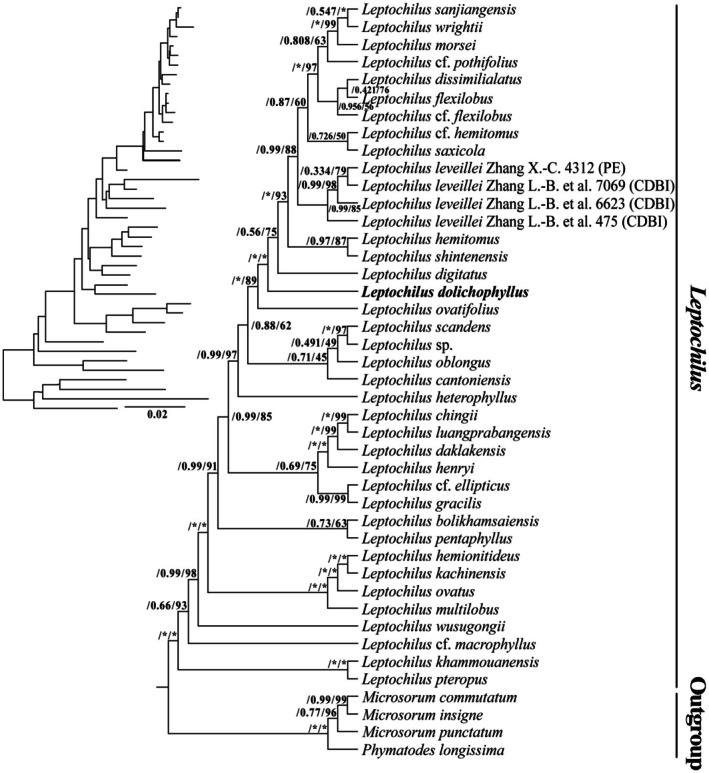 FIGURE 2
FIGURE 2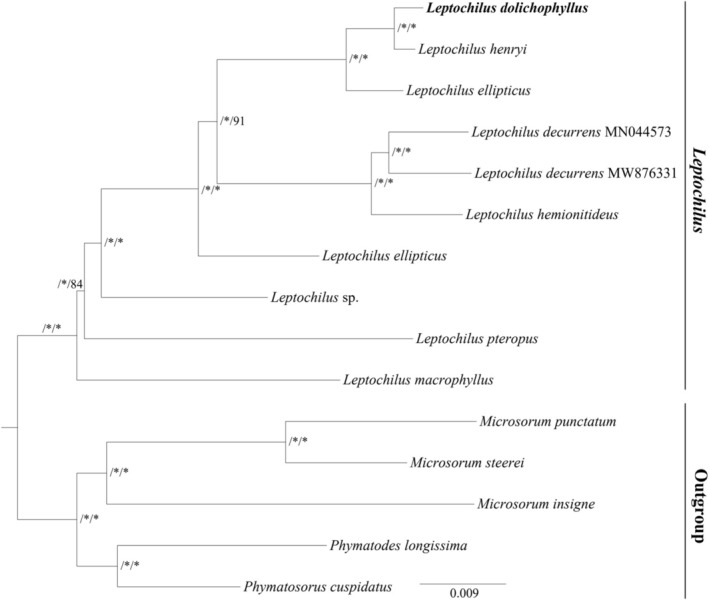 FIGURE 3
FIGURE 3 FIGURE 4
FIGURE 4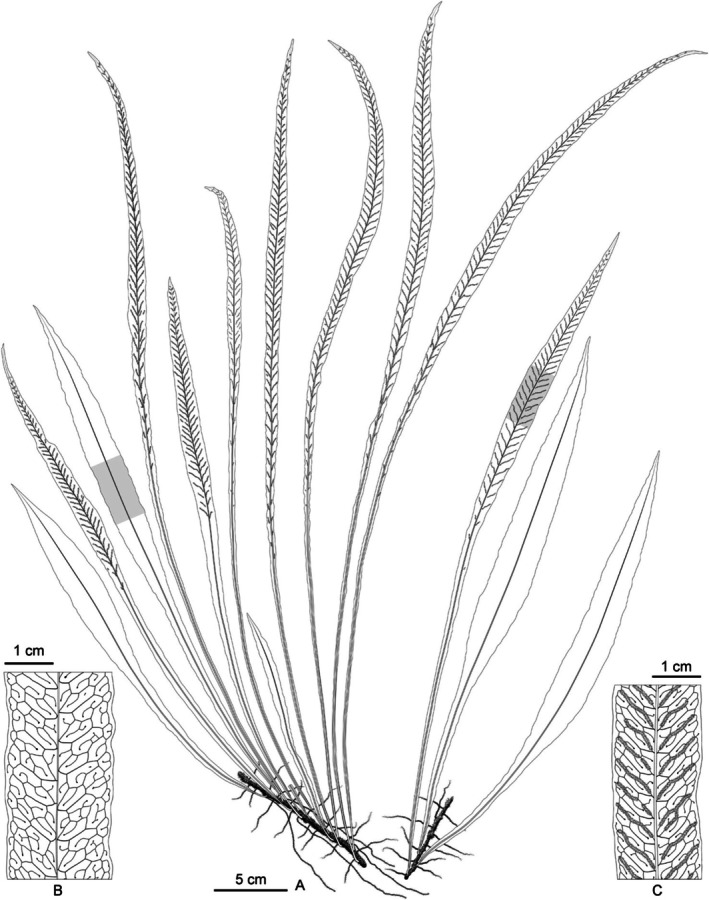 FIGURE 5
FIGURE 5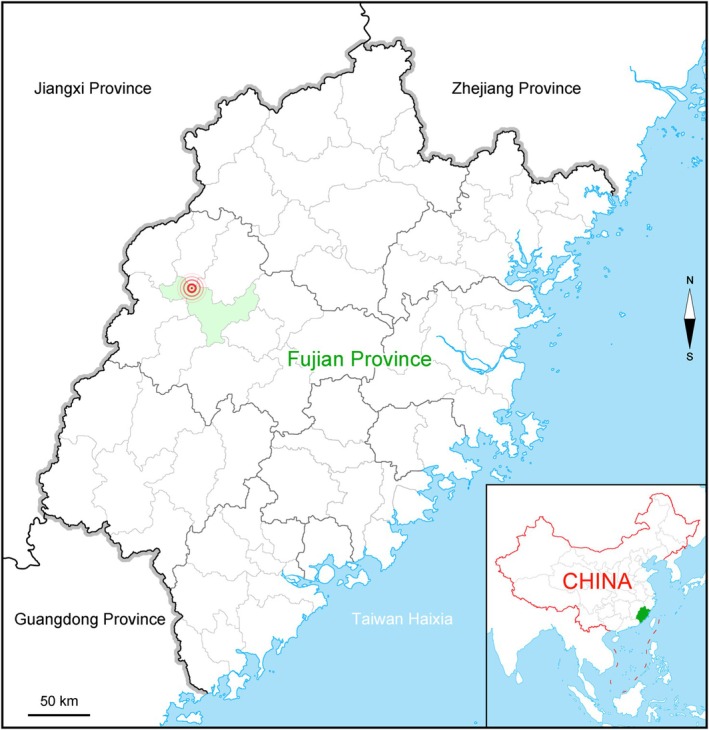 FIGURE 6
FIGURE 6| Scientific name | Voucher |
|
|
|
|---|---|---|---|---|
|
| Zhuo Z. et al. LZ156 (KUN) | |||
|
| Dong S.‐Y. 1034 (IBSC) | — | ||
|
| Chen C.‐C. 1065 (H) | |||
|
| Zhang L.‐B. et al. 6710 (CDBI, MO, VNMN) | |||
|
| Zhang L.‐B. et al. 6484 (CDBI, MO, VNMN) | |||
|
| Cicuzza D. 1998 (HITBC) | |||
|
| Zhang L.‐B. et al. 7453 (CDBI, MO, VNMN) | |||
|
| Zhang L.‐B. et al. 8944 (CCDBI) | |||
|
| A.R. Smith 00–036 (UC) | |||
|
| Zhang L.‐B. et al. 6362 (CDBI, MO, VNMN) | |||
|
| She‐Lang Jin, Hou‐Hua Fu JSL9400 | |||
|
| Zhang L. et al. 3050 (KUN) | |||
|
| Liang Z.‐L. et al. 607 (KUN, CDBI) | — | ||
|
| Wu S.‐K. et al. WS‐2437 (KUN) | |||
|
| Zhang X.‐C. 3302 (PE) | |||
|
| DJY04047 (CDBI) | |||
|
| Wu S.‐K. et al. WP‐136 (KUN) | |||
|
| Deng Y.‐F. et al.3200 (CDBI) | |||
|
| Shui Y.M. et al. LK136 (KUN) | |||
|
| Zhang L.‐B. et al. 475 (CDBI) | |||
|
| Zhang X.‐C. 4312 (PE) | |||
|
| Zhang L.‐B. et al. 6623 (CDBI) | — | ||
|
| Zhang L.‐B. et al. 7069 (CDBI) | |||
|
| Zhuo Z. et al. LZ415 (KUN) | |||
|
| Wade 1772 (TAIF) | |||
|
| Jin S.‐L. et al. JSL7322 (CSH) | — | ||
|
| YLZB2004 (CDBI) | — | ||
|
| Zhang L.‐B. et al. 6299 (CDBI, MO, VNMN) | |||
|
| Zhang L. NAS20180929_4 (KUN) | — | ||
|
| Wade 1583 (TAIF) | |||
|
| Xu C.‐D. A0357 (PE) | |||
|
| Zhang L.‐B. et al. 8014 (CDBI, MO, VNMN) | — | ||
|
| Jin S.‐L. et al. JSL7568 (CSH) | |||
|
| Zhang L.‐B. et al. 6772 (CDBI, MO, VNMN) | |||
|
| Jin S.‐L. et al. JSL8000 (CSH) | — | ||
|
| Knapp R. 3874 (P) | — | ||
|
| Knapp R. 3849 (P) | |||
|
| Li 47 (PE) | |||
|
| Wu S.‐K. et al. WS‐2591 (KUN) | |||
| Outgroup | ||||
|
| Wade 3768 (TAIF) | |||
|
| Liu 204 (PE) | |||
|
| Wade 1390 (TAIF) | |||
|
| Cheng X. et al. FB042 (KUN) | |||
| Scientific name | Accession number |
|---|---|
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
| Outgroup | |
|
| |
|
| |
|
| |
|
| |
|
| |
| Character |
|
|
|
|---|---|---|---|
| Rhizome scale shape | Narrowly lanceolate to narrowly ovate‐lanceolate | Ovate to ovate‐lanceolate | Lanceolate to ovate‐lanceolate |
| Rhizome scale color | Dark brown or nearly dark | Brown | Dark brown |
| Rhizome scale margin | Nearly entire or sparsely denticulate | Nearly entire or sparsely denticulate | Nearly entire, occasionally with 1 or 2 min teeth |
| Rhizome scale apex | Long acuminate | Acuminate | Acuminate |
| Frond dimorphism | Slightly dimorphic | Monomorphic or slightly dimorphic | Monomorphic |
| Fertile frond shape | Linear to linear lanceolate | Oblanceolate to linear lanceolate | Ovate to broadly ovate, deeply pinnatifid |
| Fertile lamina size | 38.0–63.0 × 1.0–2.0 cm | 20.0–40.0 × 1.0–4.0 cm | 9.0–21.0 × 5.0–18.0 cm |
| Stipe length | 0.4–3.0 cm | 4.0–8.0 cm | 5.0–28.0 cm |
| Life form | Hemiepiphytic | Terrestrial or epilithic | Hemiepiphytic |
- —Forestry Peak Discipline Construction Project of Fujian Agriculture and Forestry University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFern and Epiphyte Biology · Plant Diversity and Evolution · Plant and animal studies
Introduction
1
The genus Leptochilus Kaulfuss (1824, 147), a member of the family Polypodiaceae, is primarily distributed across subtropical and tropical regions of Asia, Australia, as well as adjacent Pacific islands. This genus is morphologically characterized by long‐creeping rhizomes bearing clathrate to subclathrate scales that typically do not fully cover the rhizome; laminae ranging from simple to pinnatifid; anastomosing venation; and sori that are orbicular, elongate to linear, or occasionally acrostichoid. The genus Leptochilus displays extensive morphological variation, making it one of the most taxonomically challenging groups within Polypodiaceae. The taxonomic chaos has led to ongoing debate regarding the number of species within this genus. Based on a sampling of 40 Leptochilus species, Zhang et al. (2019), Zhang, Lian, et al. (2024), and Zhang, Lu, et al. (2024) conducted a comprehensive phylogenetic study and proposed that the genus comprises approximately 50 species. However, only 33 Leptochilus species were accepted by Govaerts et al. (2021).
China stands out as the epicenter of Leptochilus species globally, hosting 31 species (Zhang, Lian, et al. 2024), which include a newly published species designated as L. tridigitatus H.J.Wei (Wei et al. 2024). Interestingly, among these, only one species, L. scandens H.J.Wei & Yi Huang, is noted for its hemiepiphytic growth habit (Zotz et al. 2021; Wei et al. 2023). Herein, we introduce an additional hemiepiphytic Leptochilus species.
In 2022, the first author had photographed a plant belonging to the genus Leptochilus and named Leptochilus leveillei (Christ) X. C. Zhang & Noot. during a vascular plant survey in the Junzifeng National Nature Reserve in Fujian Province, southeastern China. The species was first observed as an epiphyte growing on Adina pilulifera (which belongs to the family Rubiaceae). However, one of the authors, Hong‐Jin Wei, learned that the plant was found climbing on some higher tree trunks, suggesting that the newly discovered plant might be a hemiepiphyte. This hypothesis was corroborated by the subsequent field investigations, which indicate that the plant should represent a different taxon. Morphologically, this plant is superficially similar to L. leveillei , an epilithic or terrestrial species that occasionally can extend its rhizomes onto lower tree trunks. We confirmed that this novel species does not match any previously known Leptochilus species after morphological and phylogenetic studies. Here, we describe it as a new species and used molecular markers to explore its phylogenetic position. Additionally, we reported the plastid genome of this new species and used it to reconstruct phylogenetic relationships of the genus Leptochilus.
Material and Methods
2
Habitat Investigation and Sampling
2.1
The species was discovered in a stream habitat at Shuikou Village, Gaiyang Town, within the Junzifeng National Nature Reserve, Mingxi County, Fujian Province, China. An extensive field research was conducted in 2023. As many as possible similar plants were searched for in the area where the original observation had been made. Their growth habits, especially their roots, were observed carefully and photographed. Several samples of the species were collected from different individuals. Some individuals were taken to the greenhouse in Shanghai Chenshan Botanical Garden for further study.
Ploidy Assessment
2.2
Fresh leaves of the individuals in the greenhouse introduced from the field were used to estimate the levels of ploidy by flow cytometry.
Morphological Description
2.3
Morphological data of the plants was obtained from the specimens collected during the fieldwork. Vouchers were deposited in the Dendrological Herbarium of Forestry College, Fujian Agriculture and Forestry University (FJFC) and Shanghai Chenshan Herbarium (CSH). The specimens as well as rhizome scales were photographed with a digital camera (OM SYSTEM OM‐1) and a stereo microscope (Nikon SMZ18), respectively. Comparative morphological analysis of the collected material and specimens of Leptochilus henryi (CSH0038826, CSH0038838, PE02051386) and L. scandens (CSH0193333) support its recognition as a distinct species within the genus Leptochilus. Fresh rhizomes were sectioned using a Flying Eagle brand single‐edged safety razor blade and subsequently examined under a microscope.
Taxon Sampling and Sequencing
2.4
Fresh leaves were dried in silica gel. Total DNA was extracted using the cetyltrimethylammonium bromide (CTAB) method (Allen et al. 2006) and subsequently sequenced on an Illumina HiSeq 4000 platform (Illumina, San Diego, CA, USA), yielding 150‐bp paired‐end reads. Over 30 Gb raw data were generated. Quality control was conducted by FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) with default parameters.
Plastome Assembly and Annotation
2.5
The complete plastid genome of this new species was assembled by the GetOrganelle pipeline (Jin et al. 2020). The Plastid Genome Annotator (PGA) software (Qu et al. 2019) was used to annotate the complete plastid genomes, and the published sequence of Leptochilus decurrens (MN044573) was used as a reference. The annotated plastomes were manually adjusted in Geneious R11.1.5 (Kearse et al. 2012). The annotation circle maps were drawn by OGDRAW (Greiner et al. 2019).
Phylogenetic Analyses
2.6
In this study, species circumscriptions follow the framework established by Zhang et al. (2019), Zhang, Lian, et al. (2024), and Zhang, Lu, et al. (2024). A total of 39 rbcL, 40 rps4 and rps4‐trnS, and 38 trnL + trnL‐trnF sequences from 42 taxa were downloaded from GenBank to reconstruct the phylogenetic tree (Table 1). A total of 14 plastome sequences were downloaded from GenBank to reconstruct the phylogenomic relationship (Table 2). The molecular sequences were aligned using MAFFT (Katoh et al. 2002) with default parameters. The aligned sequences were then concatenated into a single matrix using the “Concatenate Sequence” script in PhyloSuite (Zhang et al. 2020). The phylogenetic trees were inferred using IQ‐TREE (Nguyen et al. 2014), incorporating the ultrafast bootstrap (UFBoot) and the approximate Bayes Test.
Results
3
Morphological Characters
3.1
Morphologically, a detailed comparison between the new species and its morphologically most similar species, L. leveillei, shows that the new species can be distinguished by narrower rhizome scales, shorter stipes, and longer lamina (Table 3).
TABLE 3: Morphological comparison between Leptochilus dolichophyllus and Leptochilus leveillei .
Characteristics of the Plastid Genome
3.2
We assembled the complete plastid genome of this new species, which possesses a total genome size of 152,503 bp with 44.0% GC content (Figure 1). The plastome displayed a typical quadripartite structure with a pair of IRs (24,631 bp), an LSC region (81,437 bp), and an SSC region (21,804 bp). The plastomes of this new species encoded 132 genes, comprising 89 protein‐coding genes, 35 transfer RNA (tRNA) genes, and 8 ribosomal RNA (rRNA) genes.
The plastome annotation map of Leptochilus dolichophyllus. The darker gray in the inner circle corresponds to GC content. The IRA and IRB (two inverted repeating regions); LSC (large single‐copy region); and SSC (small single‐copy region) are indicated outside of GC content.
Characteristics of Life Form
3.3
Ecologically, all individuals of the plant at various growth stages are observed growing on tree trunks near a stream within a broad‐leaved forest. Most individuals are hemiepiphytic on A. pilulifera, and occasionally on Alniphyllum fortunei (which belongs to the family Styracaceae). They typically occur at heights above 1.5 m, with some reaching up to 2.0 m. The juvenile plants are initially epiphytic. The roots emerged from the rhizomes and extended toward the ground. Mature individuals consistently had established root contact with the soil, indicating a transition from an epiphytic to a hemiepiphytic stage. This demonstrates that the plant is unequivocally a hemiepiphyte and represents a distinct taxon from L. leveillei .
Characteristics of Ploidy and Genome Size
3.4
Cytologically, the plant showed a mean DNA content of 11.81 Gb, similar to that of L. scandens (11.82 Gb), and was inferred to be diploid.
Phylogenetic Relationships
3.5
The aligned matrix lengths for plastid regions rbcL, rps4 + rps4‐trnS, and trnL + trnL‐trnF sequences were 1269, 1108, and 926 bp, respectively. The concatenated matrix length was 3303 bp, with 818 (24.77%) variable sites and 390 (11.81%) parsimony informative sites.
The phylogenetic relationships of the genus Leptochilus were reconstructed based on three traditional molecular markers, indicating that the new Leptochilus species formed a distinct clade with strong support (Figure 2). Additionally, we conducted a phylogenetic analysis based on complete plastome sequences, indicating that the Leptochilus species formed a well‐supported monophyletic group (aBayes = 1.00, BS = 100) (Figure 3). The new species was sister to L. henryi with strong support (aBayes = 1.00, BS = 100).
*ML phylogenetic tree based on combined plastid genome regions (rbcL, rps4 + rps4‐trnS, and trnL + trnL‐trnF). The numbers near the nodes are approximate Bayesian posterior probabilities and bootstrap percentages (aBayes supports and BP). Node is the 1.00 posterior probability or 100 bootstrap percentage.
*Phylogenetic tree obtained by ML analysis of the plastomes matrix. The numbers near the nodes are approximate Bayesian posterior probabilities and bootstrap percentages (aBayes supports and BP). Node is the 1.00 posterior probability or 100 bootstrap percentage.
Discussion
4
This new species is the ninth species of the genus Leptochilus discovered in Fujian Province (An et al. 2023). Additionally, this species represents the second known case of a hemiepiphytic growth habit within the genus in China, with L. scandens previously being the only confirmed species exhibiting such a habit (Wei et al. 2023). Within the genus Leptochilus, many species are epilithic, epiphytic, or terrestrial, with a few capable of climbing onto lower trunks of trees. Although certain terrestrially germinating species like L. mengsongensis M.X.Zhao and L. axillaris (Cav.) Kaulf. demonstrate climbing capabilities reaching up to 2 m on tree trunks, they are not classified as hemiepiphytes under the criteria established by Zotz et al. (2021). Zhang, Lu, et al. (2024) recently reported another simple‐fronded hemiepiphyte, L. luangprabangensis Liang Zhang, Khamphanh Thepkaysone & Zhuo Zhou from Vietnam; however, the study lacks in situ photographic documentation to substantiate this claim, such as images of epiphytic juvenile plants and subadult or adult plants with adventitious roots anchoring to the ground, as those shown by Fujiwara et al. (2023) and Wei et al. (2023).
Previous phylogenetics studies of Leptochilus were mainly inferred by traditional molecular markers (Schneider 2004; Dong et al. 2008; Kreier et al. 2008; Kim et al. 2012; Testo and Sundue 2014). However, the interspecific relationships are still unclear, with moderate to weak support at numerous nodes, indicating that the phylogenetic relationships within the genus remain unresolved. Our phylogenetic analyses were consistent with previous results, and the support of clades was low to moderate (Figure 2). Further studies with broader sampling and molecular markers containing more loci are needed.
Due to the relatively simple and conserved structure, abundant informative sites, and well‐understood genetic background, plastomes have become an effective molecular resource for resolving complex phylogenetic relationships (Gitzendanner et al. 2018). However, plastome‐based studies of the genus Leptochilus have mostly focused on subfamily‐level phylogenies (Liu et al. 2021; Wei et al. 2021) or have been limited to reports of individual species' plastome sequences (Min et al. 2018; Su et al. 2019). To date, only nine plastid genomes of Leptochilus have been deposited in GenBank, most of which lack annotation information, hindering our understanding of the evolutionary history and phylogenetic relationships within the genus. To provide reliable molecular data for future phylogenetic studies of the genus Leptochilus, we assembled and annotated the plastome of the new species. The results revealed that its plastome has a genome size of 152,503 bp and contains 132 genes, which is entirely within the previously reported size range of Leptochilus plastomes, from 152,084 bp ( L. macrophyllus ) to 157,508 bp (L. pteropus) (Min et al. 2018; Su et al. 2019). The phylogenetic analysis based on whole plastome sequences showed that species of this genus form a highly supported monophyletic group, and the interspecific phylogenetic resolution has greatly improved. The newly described species is sister to L. henryi (aBayes = 1.00, BS = 100). This result indicates that plastome sequences are well suited for resolving intrageneric relationships within Leptochilus.
Overall, integrative evidence from ecological niche differentiation, morphometric disparity, and molecular phylogenetic divergence robustly corroborates the recognition of this species as a distinct species from L. leveillei .
Taxonomy Treatment
5
** Leptochilus dolichophyllus ** H.H.Fu & H.J.Wei, sp. nov. (Figures 4 and 5)
Habitat and morphology of Leptochilus dolichophyllus. (A) Habitat. (B) Habit. (C) Fertile fronds. (D) Adventitious roots establishing terrestrial connection. (E) Epiphytic young individuals with roots extending downwards. (F) Dorsal view of rhizome. (G) Ventral view of rhizome. (H) Portion of fertile frond. (I) Portion of sterile frond showing venation. (J) Lower portion of stipes with rhizome. (K) Scales from rhizome. (L) Cross section of rhizome.
Line drawings of Leptochilus dolichophyllus. (A) Habit. (B) Portion of sterile frond showing venation. (C) Portion of fertile frond showing sori.
Type: CHINA. Fujian: Sanming, Jun Zi Feng National Nature Reserve, under evergreen broad‐leaved forest, on tree trunks near a stream, 26°32′51″ N, 117°3′37″ E, elev. 352 m, 18 Sep. 2023, She‐Lang Jin, Hou‐Hua Fu JSL9400 (holotype: FJFC!, isotypes: CSH!, IBK!, KUN!, PE!).
Diagnosis
5.1
Leptochilus dolichophyllus is similar to L. leveillei , but differs by the narrower rhizome scales (narrowly ovate‐lanceolate, 2.0–6.0 mm vs. ovate‐lanceolate, 1.1–4.4 mm), shorter stipes (0.4–3.0 cm vs. 4.0–8.0 cm), longer fertile fronds (40.0–63.0 cm vs. 20.0–40.0 cm), and life form (hemiepiphytic vs. terrestrial or epilithic). Moreover, it can be distinguished from L. scandens by stipe length (0.4–3.0 cm vs. 5.0–28.0 cm), frond dimorphism (slightly dimorphic vs. monomorphic), and fronds (simple vs. pinnatipartite).
Description
5.2
Plants perennial, evergreen, hemiepiphytic. Rhizome slender, 2.0–3.0 mm in diam., dorsiventral, long scandent, scaly at apex; vascular bundles up to 10; scales pseudo‐peltate, dark brown or near dark, narrowly lanceolate to narrowly ovate‐lanceolate, 2.0–6.0 mm, margins nearly entire, or slightly denticulate, apex long acuminate. Fronds distant, slightly dimorphic, fertile fronds longer and narrower, stipe stramineous, much shorter than lamina, glabrous; venation visible, lateral veins straight or zigzag more or less and dichotomous distally, veinlets anadromous, thinner than lateral veins, included veinlets simple or forked; lamina simple, widest at middle, gradually decurrent nearly to stipe base, margin slightly undulate, herbaceous, glabrous on both surfaces, dark green adaxially, pale green abaxially. Sterile fronds: 27.0–42.0 cm long; stipe 3.0–10.0 mm long, 0.5–1.0 mm in diam. at middle; lamina lanceolate to narrowly lanceolate, 32.0–41.0 × 2.2–3.2 cm, apex acuminate to shortly acuminate; veins forming 2 or 3 rows of areoles between midrib and lamina margin. Fertile fronds: (38.0–)45.0–63.0 cm long, stipe 4.0–27.0 mm, 0.5–1.3 mm in diam. at middle, lamina linear to linear lanceolate, (37.0–)43.0–60.0 × 0.9–2.0 cm, apex long acuminate or caudate; veins forming 3 or 4 rows of irregularly arranged areoles between midrib and lamina margins, midrib raised on both surfaces. Sori linear, 1 regular row between lateral veins, from midrib or near midrib to near margin of lamina, at an angle of 30°–45° with midrib; paraphyses absent. Spores yellow, 64 per sporangium.
Geographical Distribution
5.3
China, Fujian, Sanming, Junzifeng National Nature Reserve in Fujian (Figure 6).
Geographical distribution of Leptochilus dolichophyllus (center of rings).
Ecology
5.4
Leptochilus dolichophyllus mostly climbs on tree trunks in the understory of moist montane forest, about 350 m above mean sea level.
Etymology
5.5
The epithet of L. dolichophyllus is taken from the Greek compound word formed by “dolichos” (long) and “phyllon” (leaf), together meaning “long‐leaved,” referring to the elongated leaves morphology of the new species. Most individuals of L. dolichophyllus are hemiepiphytic on A. pilulifera and occasionally occur on Al. fortunei.
Vernacular Name
5.6
Simplified Chinese: 长叶线蕨; Chinese pinyin: cháng yè xiàn jué.
Conservation Assessments
5.7
The population of L. dolichophyllus covers an area of about 50–100 square meters, with about 50 individuals (clones of the same individual excluded) encountered in the field. As the species is currently known only from a single locality, we recommend its classification as Data Deficient under the IUCN Red List criteria (IUCN 2012).
Author Contributions
Hou‐Hua Fu: conceptualization (equal), data curation (equal), formal analysis (lead), investigation (equal), visualization (equal), writing – original draft (equal). Cheng‐Yuan Zhou: data curation (equal), software (lead), visualization (supporting). Xiong‐De Tu: data curation (equal), validation (equal). Liang Ma: conceptualization (equal), investigation (equal), methodology (equal). Shi‐Pin Chen: conceptualization (equal), funding acquisition (lead), methodology (equal), project administration (lead), writing – review and editing (equal). Hong‐Jin Wei: investigation (equal), visualization (lead), writing – original draft (equal), writing – review and editing (equal).
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Allen, G. C. , M. A. Flores‐Vergara , S. Krasynanski , S. Kumar , and W. F. Thompson . 2006. “A Modified Protocol for Rapid DNA Isolation From Plant Tissues Using Cetyltrimethylammonium Bromide.” Nature Protocols 1: 2320–2325. 10.1038/nprot.2006.384.17406474 · doi ↗ · pubmed ↗
- 2An, C. , Y. X. Zhuang , P. Zheng , Y. X. Lin , C. Z. Yang , and Y. Qin . 2023. “A Checklist of Vascular Plants in Fujian Province, China.” Biodiversity Science 31, no. 6: 19–27. 10.17520/biods.2022537. · doi ↗
- 3Dong, X. D. , S. G. Lu , and C. X. Li . 2008. “Molecular Phylogeny of Colysis (Polypodiaceae) Based on Chloroplast rbc L and rps 4‐trn S Sequences.” Journal of Systematics and Evolution 46: 658–666.
- 4Fujiwara, T. , H. Q. Bùi , T. Shuichiro , M. Noriaki , and O. Emiko . 2023. “ Leptochilus ornithopus (Polypodiaceae), a New Hemiepiphytic Fern Species From Central Highlands of Vietnam.” Phytotaxa 584: 149–160. 10.11646/phytotaxa.584.3.2. · doi ↗
- 5Gitzendanner, M. A. , P. S. Soltis , G. K. S. Wong , B. R. Ruhfel , and D. E. Soltis . 2018. “Plastid Phylogenomic Analysis of Green Plants: A Billion Years of Evolutionary History.” American Journal of Botany 105: 291–301. 10.1002/ajb 2.1048.29603143 · doi ↗ · pubmed ↗
- 6Govaerts, R. , E. N. Lughadha , N. Black , R. Turner , and A. Paton . 2021. “The World Checklist of Vascular Plants, a Continuously Updated Resource for Exploring Global Plant Diversity.” Scientific Data 8: 215. 10.1038/s 41597-021-00997-6.34389730 PMC 8363670 · doi ↗ · pubmed ↗
- 7Greiner, S. , P. Lehwark , and R. Bock . 2019. “Organellar Genome DRAW (OGDRAW) Version 1.3.1: Expanded Toolkit for the Graphical Visualization of Organellar Genomes.” Nucleic Acids Research 47, no. W 1: W 59–W 64.30949694 10.1093/nar/gkz 238PMC 6602502 · doi ↗ · pubmed ↗
- 8IUCN . 2012. Red List Categories and Criteria: Version 3.1. 2nd ed. iv+32 pp. IUCN. https://www.iucnredlist.org/.