A survey of the spider genus Lipocrea Thorell, 1878 (Araneae, Araneidae) from Guiyang City, Southwest China: An integrated morphological and molecular approach

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3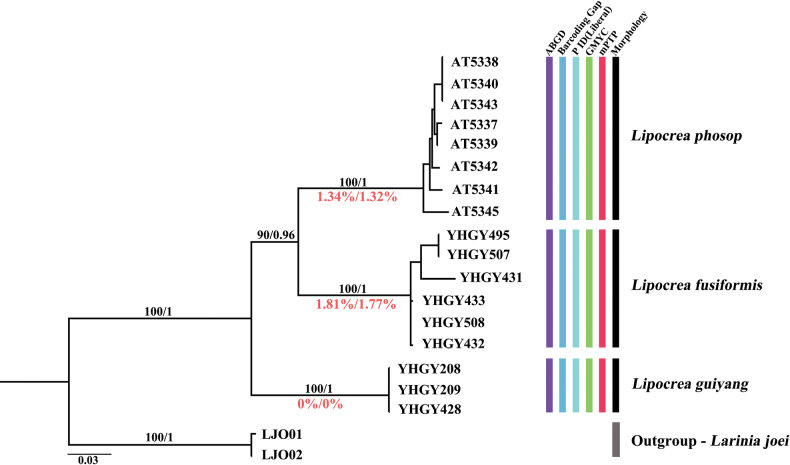 Figure 4
Figure 4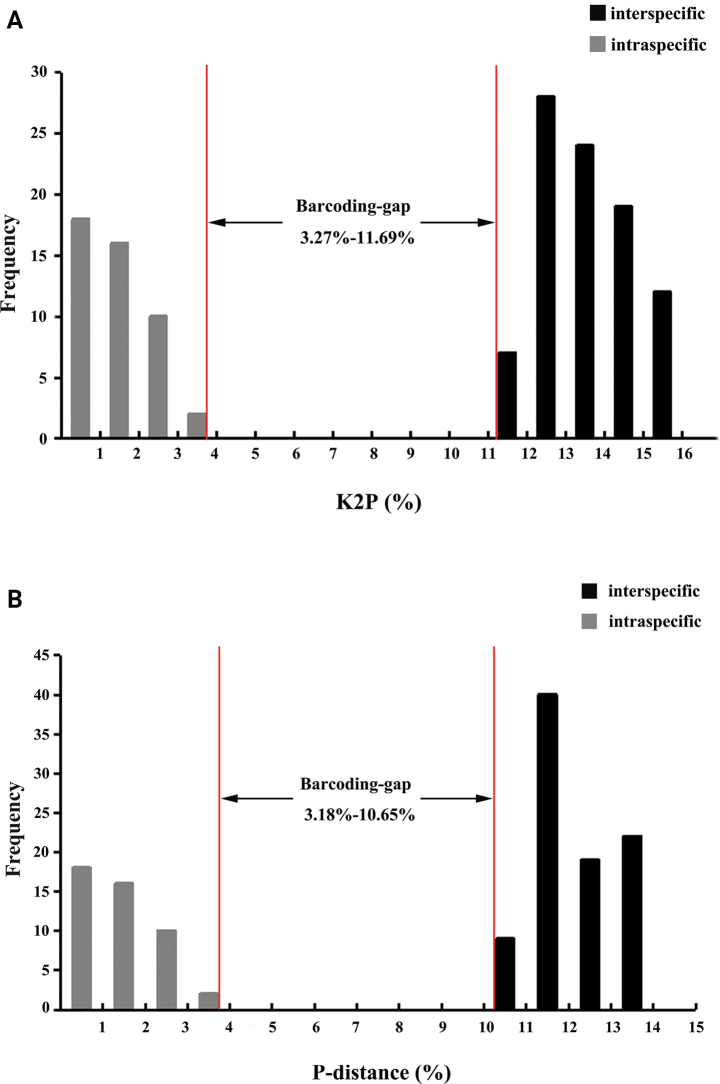 Figure 5
Figure 5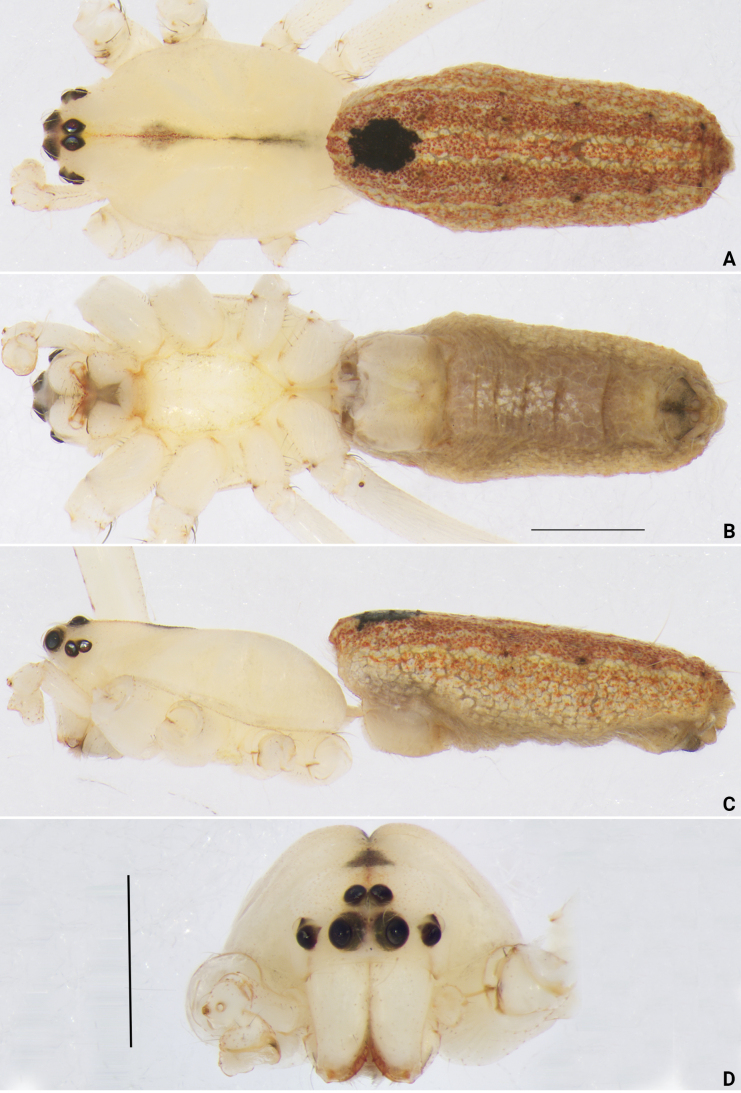 Figure 6
Figure 6 Figure 7
Figure 7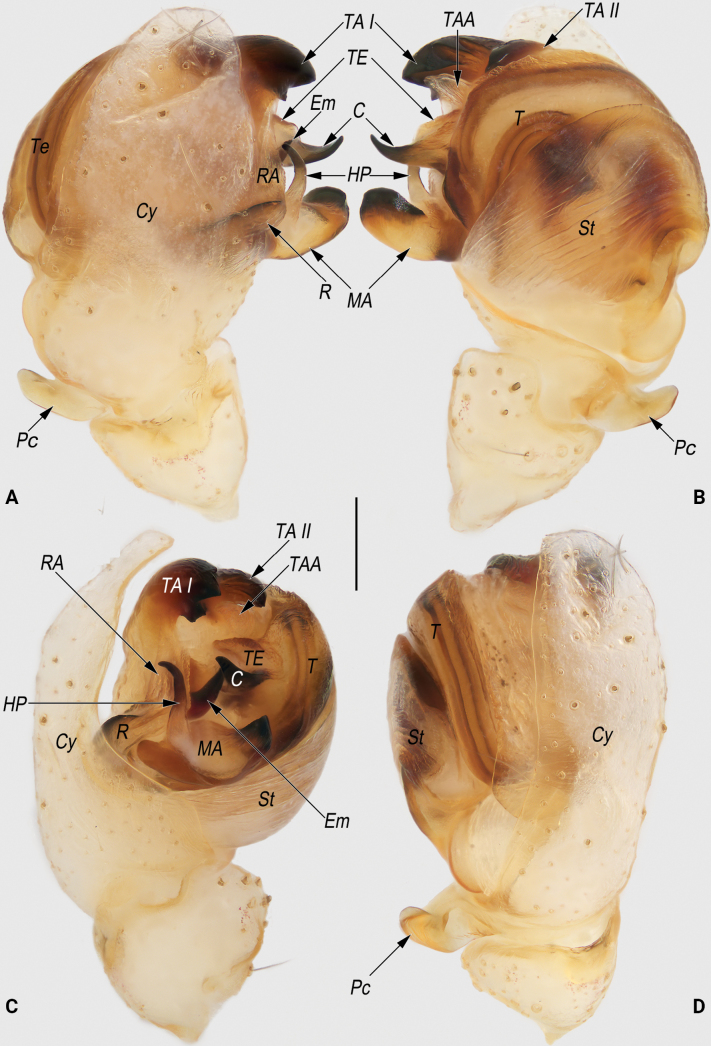 Figure 8
Figure 8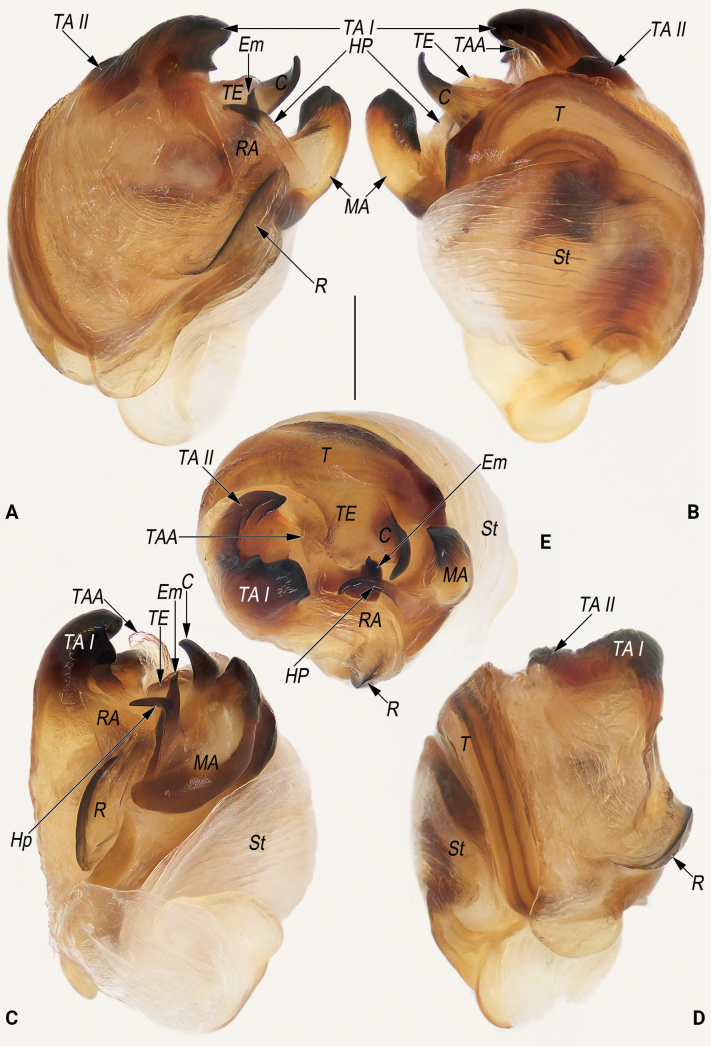 Figure 9
Figure 9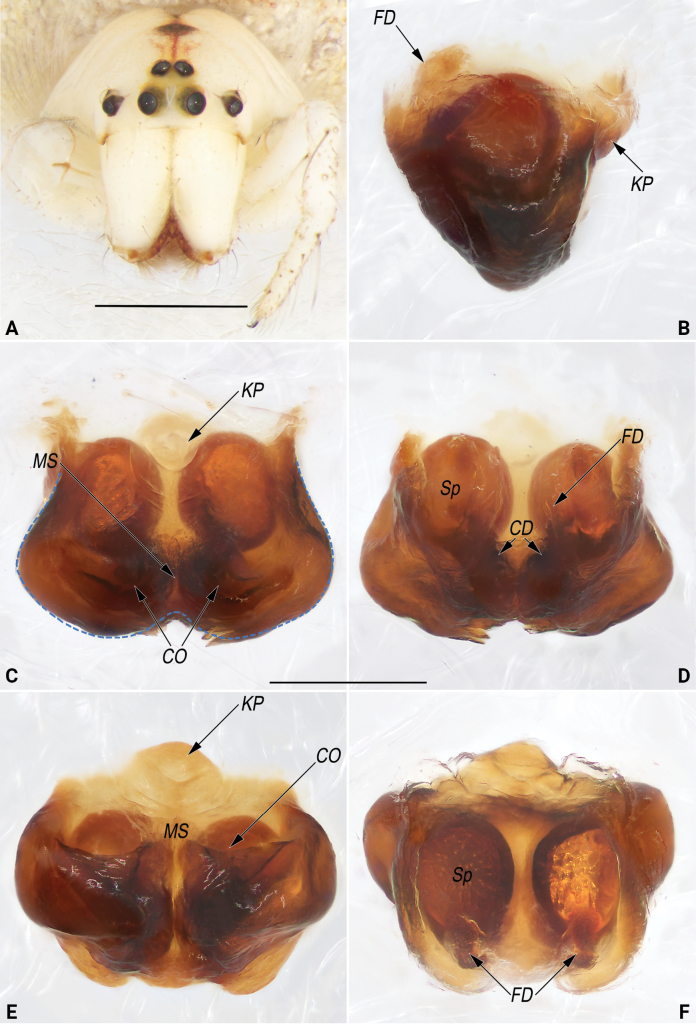 Figure 10
Figure 10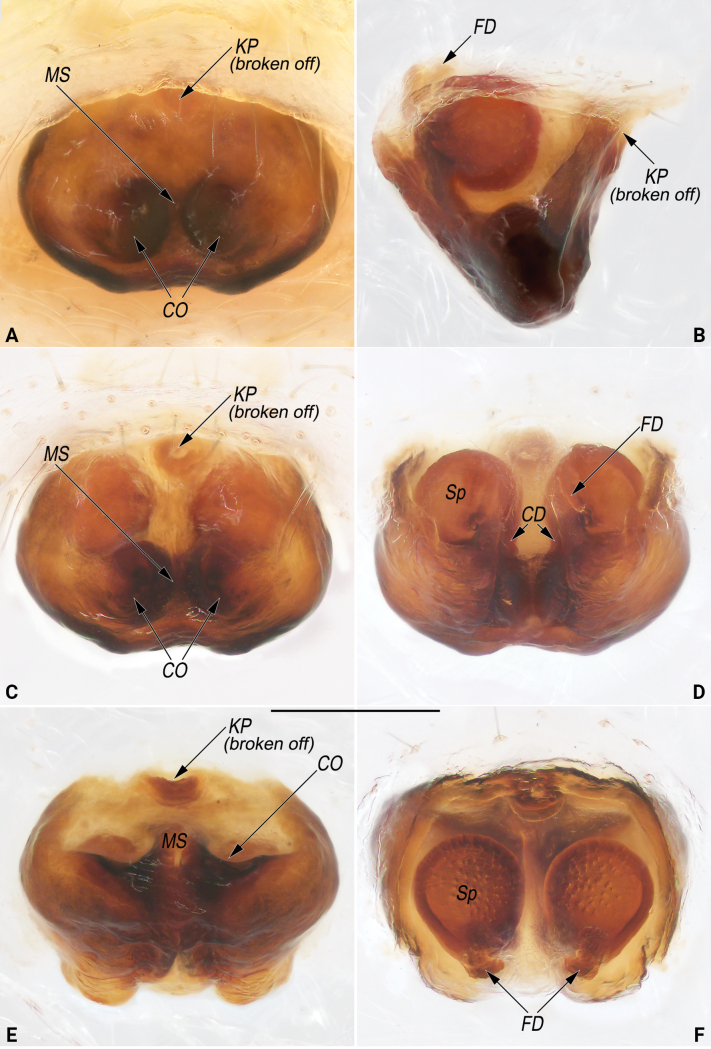 Figure 11
Figure 11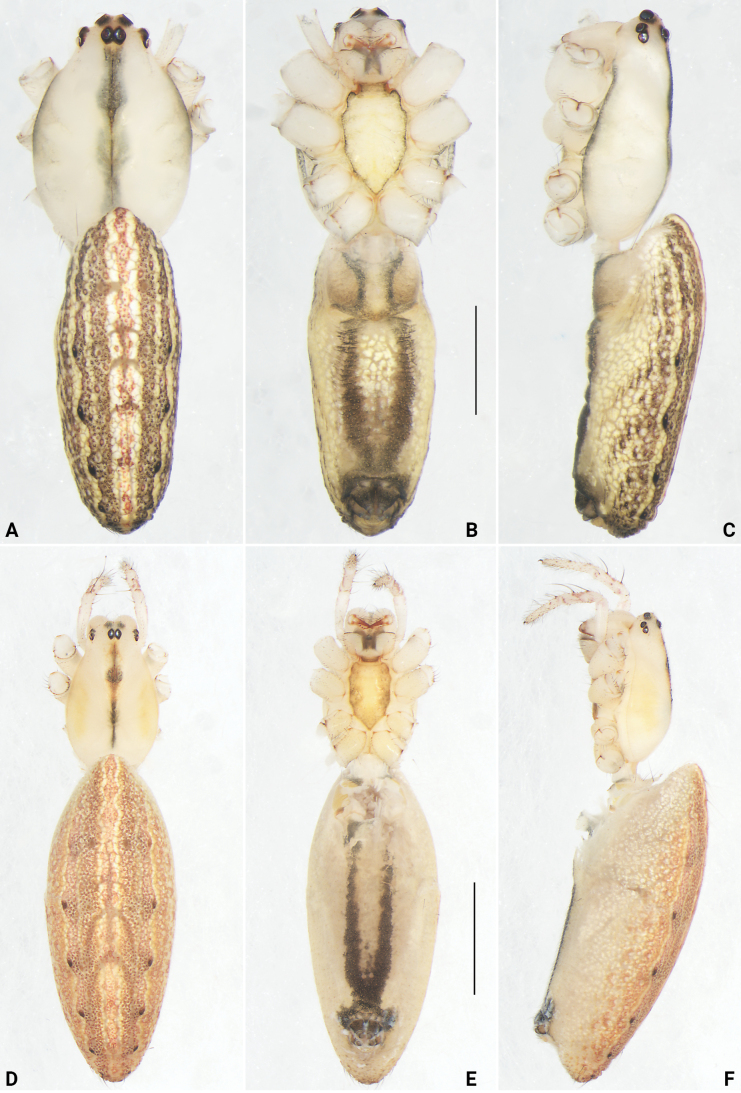 Figure 12
Figure 12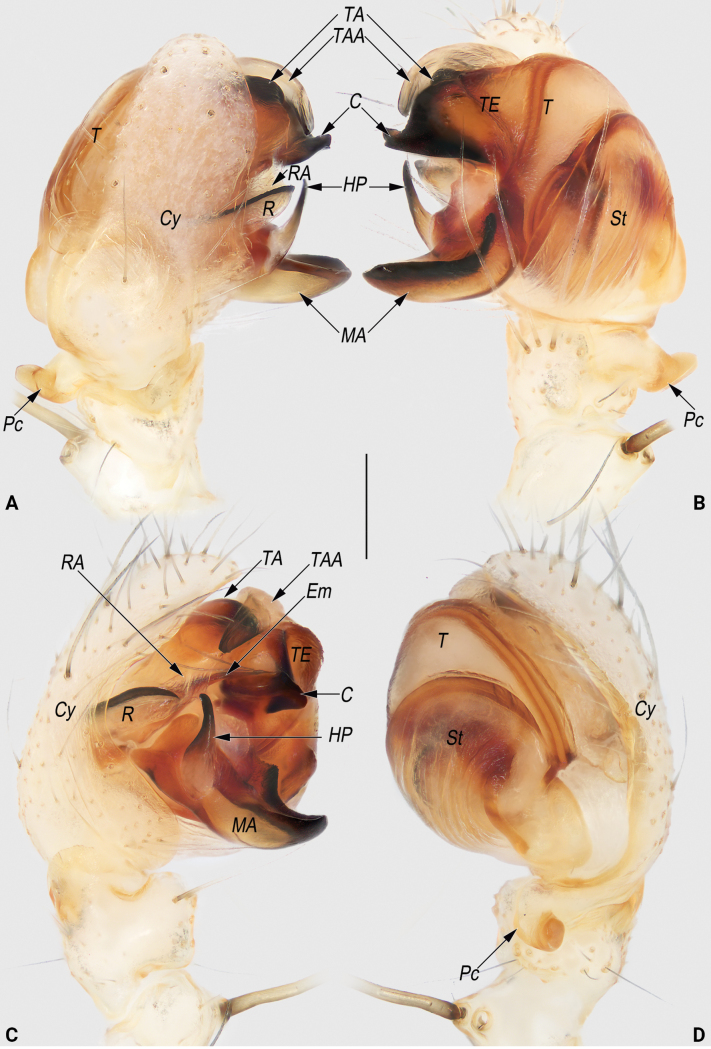 Figure 13
Figure 13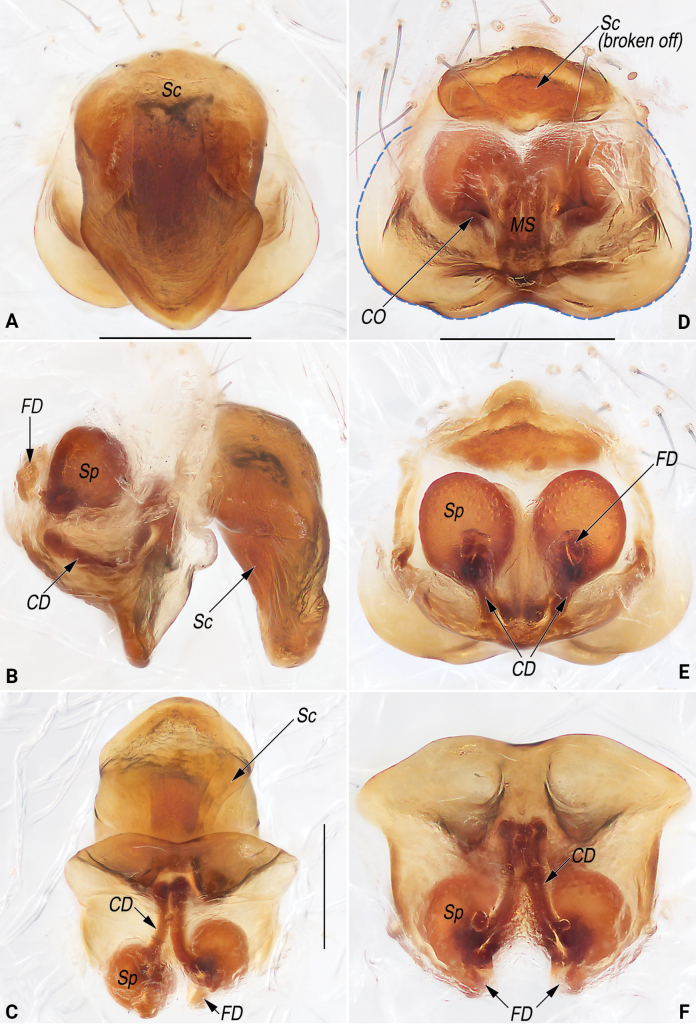 Figure 14
Figure 14| Specimen code | Genus/Species | Sex | Locality | Country | Coordinates | Elevation (m a.s.l.) | |
| YHGY208 | ♂ | Nanjiang Grand Canyon, Kaiyang County, Guiyang City, Guizhou Prov. | China |
| 861 |
| |
| YHGY209 | ♀ | Nanjiang Grand Canyon, Kaiyang County, Guiyang City, Guizhou Prov. | China |
| 861 |
| |
| YHGY428 | ♀ | Nanjiang Grand Canyon, Kaiyang County, Guiyang City, Guizhou Prov. | China |
| 861 |
| |
| YHGY431 |
| ♂ | Dangwu Town, Huaxi District, Guiyang City, Guizhou Prov. | China |
| 1138 |
|
| YHGY432 |
| ♀ | University Town, Huaxi District, Guiyang City, Guizhou Prov. | China |
| 1173 |
|
| YHGY433 |
| ♀ | Dangwu Town, Huaxi District, Guiyang City, Guizhou Prov. | China |
| 1138 |
|
| YHGY495 |
| ♂ | University Town, Huaxi District, Guiyang City, Guizhou Prov. | China |
| 1173 |
|
| YHGY507 |
| ♀ | University Town, Huaxi District, Guiyang City, Guizhou Prov. | China |
| 1173 |
|
| YHGY508 |
| ♀ | University Town, Huaxi District, Guiyang City, Guizhou Prov. | China |
| 1173 |
|
| AT5343 |
| ♂ | Mai Khet, Mueang Prachinburi District, Prachinburi Prov. | Thailand |
|
| |
| AT5337 |
| ♀ | Mai Khet, Mueang Prachinburi District, Prachinburi Prov. | Thailand |
|
| |
| AT5338 |
| ♀ | Mai Khet, Mueang Prachinburi District, Prachinburi Prov. | Thailand |
|
| |
| AT5339 |
| ♀ | Mai Khet, Mueang Prachinburi District, Prachinburi Prov. | Thailand |
|
| |
| AT5340 |
| ♀ | Mai Khet, Mueang Prachinburi District, Prachinburi Prov. | Thailand |
|
| |
| AT5341 |
| ♀ | Mai Khet, Mueang Prachinburi District, Prachinburi Prov. | Thailand |
|
| |
| AT5342 |
| ♂ | Mai Khet, Mueang Prachinburi District, Prachinburi Prov. | Thailand |
|
| |
| AT5345 |
| ♂ | Ban Pathum, Sam Khok District, Pathum Thani Prov. | Thailand |
|
| |
| LJO01 |
| ♂ | Khlong Sam, Pathum Thani Prov. | Thailand |
|
| |
| LJO02 |
| ♀ | Khlong Sam, Pathum Thani Prov. | Thailand |
|
|
| Substitution model | Pmin/Pmax | X | Partition | Prior intraspecific divergence (P) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.001 | 0.0017 | 0.0028 | 0.0046 | 0.0077 | 0.0129 | 0.0215 | 0.0359 | 0.0599 | 0.1000 | ||||
| JC | 0.001/0.1 | 1.5 | Initial | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Recursive | 9 | 9 | 9 | 9 | 6 | 5 | 3 | 3 | 3 | 3 | |||
|
| 0.001/0.1 | 1.5 | Initial | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Recursive | 9 | 9 | 9 | 9 | 6 | 5 | 3 | 3 | 3 | 3 | |||
| Simple | 0.001/0.1 | 1.5 | Initial | 5 | 5 | 5 | 5 | 3 | 3 | 3 | |||
| Recursive | 6 | 6 | 6 | 5 | 4 | 3 | 3 | ||||||
| JC | 0.001/0.1 | 2 | Initial | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Recursive | 9 | 9 | 9 | 9 | 5 | 5 | 3 | 3 | 3 | 3 | |||
|
| 0.001/0.1 | 2 | Initial | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Recursive | 9 | 9 | 9 | 9 | 5 | 5 | 3 | 3 | 3 | 3 | |||
| Simple | 0.001/0.1 | 2 | Initial | 3 | 3 | 3 | 3 | 3 | 3 | 3 | |||
| Recursive | 6 | 5 | 5 | 4 | 3 | 3 | 3 | ||||||
| 0.0001 | 0.0002 | 0.0005 | 0.0013 | 0.0029 | 0.0068 | 0.0159 | 0.0860 | ||||||
| JC | 0.0001/0.2 | 1.5 | Initial | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | ||
| Recursive | 9 | 9 | 9 | 9 | 9 | 6 | 5 | 3 | |||||
|
| 0.0001/0.2 | 1.5 | Initial | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | ||
| Recursive | 9 | 9 | 9 | 9 | 9 | 6 | 5 | 3 | |||||
| Simple | 0.0001/0.2 | 1.5 | Initial | 5 | 5 | 5 | 5 | 5 | 3 | 3 | |||
| Recursive | 6 | 6 | 6 | 6 | 6 | 4 | 3 | ||||||
| JC | 0.0001/0.2 | 2 | Initial | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | ||
| Recursive | 9 | 9 | 9 | 9 | 9 | 5 | 5 | 3 | |||||
|
| 0.0001/0.2 | 2 | Initial | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | ||
| Recursive | 9 | 9 | 9 | 9 | 9 | 5 | 5 | 3 | |||||
| Simple | 0.0001/0.2 | 2 | Initial | 3 | 3 | 3 | 3 | 3 | 3 | 3 | |||
| Recursive | 6 | 6 | 6 | 6 | 5 | 3 | 3 | ||||||
| Putative species | Intraspecific | Closest interspecific | P ID (Liberal) | Closest P ID (Liberal) species | Intra/Inter |
|---|---|---|---|---|---|
| 0/0 | 0.1349/0.1219 | 1.00 (0.86, 1.0) |
| 0.02 | |
|
| 0.0134/0.0132 | 0.1275/0.1151 | 0.97 (0.90, 1.0) |
| 0.12 |
|
| 0.0181/0.0177 | 0.1275/0.1151 | 0.96 (0.86, 1.0) |
| 0.14 |
| Analysis | Clusters (CI) | Entities (EI) | Likelihood (null) | Likelihood (GMYC) | Likelihood ratio | Threshold |
|---|---|---|---|---|---|---|
| Single | 3(1–5) | 3(1–17) | 96.89582 | 98.26415 | 2.736648 | -0.017105236 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSpider Taxonomy and Behavior Studies · Genetic diversity and population structure · Lepidoptera: Biology and Taxonomy
Introduction
Lipocrea Thorell, 1878 is a small spider genus that was originally established to include three species: L. phthisica (L. Koch, 1871) and L. tabida (L. Koch, 1872) from the Australasian Region, and the type species L. fusiformis (Thorell, 1877) from the Oriental Region. An additional Oriental species, L. diluta Thorell, 1887 was subsequently described. However, the validity of Lipocrea as a distinct genus was not recognized by Simon (1887: 187), who treated these species under the genus Larinia Simon, 1874. Of the aforementioned species, L. phthisica has since been transferred to Larinia sensu stricto (Grasshoff 1970). The remaining species, together with Larinia longissima (Simon, 1881) from the Ethiopian Region, were subsequently removed from Larinia and placed in a newly erected genus, LarinopaGrasshoff (1970), by Grasshoff (1970: 226). Although the name Lipocrea remains available under the rules of zoological nomenclature, it has been largely overlooked (Levy 1986). The genus Lipocrea, as redefined by Grasshoff under the name Larinopa, is primarily distinguished from Larinia by the structure of the genitalia, especially the morphology of the male palpal organ.
Currently, the WSC (2025) lists five species under Lipocrea, none of which are known from the Chinese mainland (WSC 2025). However, in several publications, some of these species are considered to belong to Larinia. For example, L. phosop (Tanikawa, Into & Petcharad, 2023) was originally described as Larinia has not yet been formally transferred to Lipocrea. Nevertheless, due to its strong morphological similarity to the type species of Lipocrea, L. phosop has been provisionally placed in Lipocrea by the WSC (2025). Even the type species, L. fusiformis, has frequently been placed in Larinia and redescribed as such by various authors, including Tanikawa (1989, 2007, 2009), Okuma et al. (1993), Barrion and Litsinger (1995), and Chang and Tso (2004). In view of the above-mentioned, the validity of the genus Lipocrea and the current generic placement of its constituent species remains in dispute.
A preliminary genus-level taxonomic molecular analysis of Grasshoff’s (1970) ‘Larinia group’ was carried out based on all available COI sequences of the Larinia group and related genera (69 from NCBI and 9 newly sequenced here) (Suppl. material 1: table S1). According to the results (Suppl. material 1: fig. S1): (1) the monophyly of the genus Lipocrea is well supported; (2) Larinia is polyphyletic, which is consistent with the results of Scharff et al. (2020). Based on this preliminary result and considering that a formal comprehensive revision of the Larinia group is lacking, we agree with Framenau and Castanheira (2022) that the Larinia group requires further systematic study that includes Larinia lineata Lucas, 1846 (the type species of the genus). A review of the Larinia group is not within the scope of this work. Consequently, the present study assigns the two species treated here to the monophyletic genus Lipocrea, rather than to the polyphyletic Larinia sensu lato.
Guiyang, the provincial capital of Guizhou Province (Fig. 1A), is the first city to receive the accolade of ‘the national forest city’ in China, famous for more than 55% forest coverage (Zhong 2018). Guiyang is also known as the ‘city of a thousand gardens’, with 1025 urban parks, and is considered one of China’s most biodiverse provincial capitals (Yang 2020; Cheng and Chen 2022). However, spiders can be regarded as being poorly represented in Guiyang, with only 64 species from 25 families recorded or described to date (JZ and HY, unpubl. data). Of these, 21 species are endemic, 16 were newly described, and 15 were reported as new records in recent years (Yu et al. 2018; Yu and Zhang 2019; Xin et al. 2020; Yan et al. 2021; Long et al. 2022; Zhang et al. 2022a, b, 2023; He et al. 2023; Li et al. 2023; Yang et al. 2023a, b, c; Ding et al. 2024; Pan et al. 2024; Jiang et al. 2025; Zhang et al. 2025). This estimate of spider diversity is assumed to be far from the true diversity within this city.
Locality of Guiyang City (A) and distribution records of Lipocrea species in Guiyang (B).
As mentioned above, spiders are poorly studied in Guiyang, including Araneidae. Only five species have been recorded: Cyrtarachne bufo (Bösenberg & Strand, 1906), Hypsosinga pygmaea (Sundevall, 1831), Neoscona xishanensis Yin, Wang, Xie & Peng, 1990, Nephila pilipes (Fabricius, 1793) and Pronoides brunneus Schenkel, 1936. Recently, short but intensive field collections in Guiyang have been conducted by the staff of the Guizhou Normal University. During these surveys, we have found some Lipocrea specimens that belong to at least two morphospecies (Figs 1–3): one is new to science, and the other one has been identified as L. fusiformis, a new record for mainland China. The sympatric distribution with L. fusiformis and the high intraspecific morphological variation in females of the new species pose significant challenges for sexual pairing and species identification. We therefore generated DNA barcode data to devise a specimen phylogeny and used five molecular species delimitation methods to test morphology-based species identification.
Living specimens of Lipocrea guiyang J. Zhang, Yu & Mi, sp. nov. A. Male; B. Female. Photographs by Q Lu (Shenzhen).
Living specimens of Lipocrea fusiformis (Thorell, 1877). A. Male; B, C. Female. Photographs by Q Lu (Shenzhen).
The goal of this paper is to: 1) use the consensus results of an integrated morphological and molecular approach to delimit Lipocrea spiders from Guiyang; 2) describe the new species under the name of Lipocrea guiyang J. Zhang, Yu & Mi, sp. nov.; 3) re-illustrate L. fusiformis based on new material from Guiyang, including supplementary micrographs; and 4) provide a distribution map of Lipocrea species in Guiyang City.
Materials and methods
Taxon sampling
Specimens in this study were collected by hand and by beating vegetation, and directly fixed in absolute ethanol. The right legs were removed and stored at −80 °C for subsequent DNA extraction. The remainder of the specimens was preserved in 80% ethanol for identification and morphological examination. A total of 11 adults were obtained, examined, and processed for DNA extraction but only nine individuals yielded useable DNA (Table 1). All voucher specimens (including types of the new species) are deposited in the Museum of Guizhou Normal University, Guiyang, China (MGNU).
Molecular protocols
Total genomic DNA was extracted using the Cell & Tissue Genomic DNA Isolation Kit (Bioteke, Beijing, China) following the manufacturer’s protocols. We amplified cytochrome c oxidase subunit I (COI) using the primer pairs LCO1490/HCO2198 (Folmer et al. 1994) and standard polymerase chain reaction (PCR) settings (Wheeler et al. 2016). PCR products were transported to the Beijing Tsingke Biotech Co., Ltd. (Chongqing, China) for sequencing using the same PCR primers. We manually edited the sequences using Geneious Prime 2024 (Kearse et al. 2012), translated nucleotide sequences into amino acids to check for stop codons, and ensured the proper configuration of codon positions.
Phylogenetic analyses
For phylogenetic analyses, the COI data of 17 specimens of Lipocrea were used as the in-group, including eight sequences of L. phosop downloaded from GenBank (Tanikawa et al. 2023; NCBI 2025). To root the tree, we included two specimens of Larinia joei Tanikawa & Petcharad, 2021 from NCBI (2025) as the outgroup (Table 1). We performed maximum-likelihood (ML) analyses using IQ-TREE v. 2.3.1 (Minh et al. 2020) based on the best COI substitution model (GTR+I+G) in jMODELTEST v. 2.1.10 (Darriba et al. 2012). Branch support was estimated with ultrafast bootstrapping with 1000 replicates (Hoang et al. 2018). Bayesian-inference (BI) was performed with MrBayes v. 3.2.1 (Ronquist et al. 2012) using one independent chain for 50 million generations. The first 10% of trees from each run were discarded as burn-in. Finally, we used FigTree v. 1.4.4 (Rambaut 2012) to visualize and manipulate trees and used Photoshop CC 2018 to summarize them.
Molecular species delimitation
To delimit three morphospecies of Lipocrea based on an accompanying morphological study of the genus, we used two genetic distance-based methods: the DNA barcoding gap (Barrett and Hebert 2005) and ABGD (Puillandre et al. 2012), as well as three methods based on the inferred tree, GMYC (Pons et al. 2006), P ID (Liberal), and mPTP (Kapli et al. 2017).
Because the P ID (Liberal) and DNA barcoding gap (Barrett and Hebert 2005) methods require a priori designation, we assigned 17 Lipocrea individuals to three putative species based on a combination of phylogenetic topology and morphological characteristics. With the DNA barcoding gap, we used the overlap between the interspecies and intraspecies Kimura two-parameter (K2P) and uncorrected p-distance for each candidate species calculated in MEGA X (Kumar et al. 2018). The P ID (Liberal) method tests species delimitation by relying on defining the putative species groups. We used the BI tree as a guide to test species hypothesis (Xu et al. 2017).
The other three methods that we used do not require terminals to be a priori assigned to putative species. ABGD calculates all pairwise distances in the data set, evaluates intraspecific divergences, and then sorts the terminals into candidate species with calculated P values. We performed the ABGD analysis on a web server (https://bioinfo.mnhn.fr/abi/public/abgd/) using three different models: Jukes-Cantor (JC69; Jukes and Cantor 1969), K2P (Kimura 1980), and simple distance (p-distance; Nei and Kumar 2000). We analyzed the data using two different values for the parameters Pmin (0.001 and 0.0001), Pmax (0.1 and 0.2), and relative gap width (X = 1.5 or 2), with the other parameters set to default values. We used the BI tree as a guide to test the species hypothesis (Xu et al. 2017). Two runs of 100 million steps were used for the mPTP logging every 1 million steps, discarding the first 2 million steps. Each run was started from a random delimitation. The GMYC methodology (Pons et al. 2006) analysis was conducted using the single-threshold model in the “splits” package (Ezard et al. 2009) for R 4.2.2 (R Development Core Team 2024). BEAST 2.6.7 (Bouckaert et al. 2014) was used to produce an ultrametric tree for the GMYC analysis. Analyses were run for 50 million steps with 10% of the trees in each chain discarded as burn-in.
Morphological protocols
Specimens were examined using an Olympus SZX7 stereomicroscope. Further details were studied under a CX41 compound microscope. Male and female copulatory organs were examined and illustrated after dissection. Epigynes were removed and cleared in lactic acid or a warm 10% potassium hydroxide (KOH) solution. Images were captured with a Canon EOS 70D digital camera (20.2 megapixels) mounted on an Olympus CX41 compound microscope and assembled using Helicon Focus v. 6.80 image-stacking software. All measurements were obtained using an Olympus SZX7 stereomicroscope and are given in millimeters. Eye diameters were measured at the widest part. The total body length does not include the chelicerae or spinnerets. Leg lengths are given as total length (femur, patella+tibia, metatarsus, tarsus). The terminology used in the text and figure legends follows Grasshoff (1970), Framenau and Scharff (2008), Framenau and Castanheira (2022), Tanikawa and Petcharad (2021), and Tanikawa et al. (2023).
References to figures in the cited papers are listed in lowercase (fig. or figs); figures from this paper are noted with an initial capital (Fig. or Figs). The abbreviations used in the text are:
AER anterior eye row;
ALE anterior lateral eyes;
AME anterior median eyes;
C conductor;
CD copulatory duct;
CO copulatory opening;
Cy cymbium;
Em embolus;
FD fertilisation duct;
HP hook-shaped process of MA;
KP knob-like projection;
MA median apophysis;
MS median septum;
MOQ median ocular quadrangle;
Pc paracymbium;
PLE posterior lateral eyes;
PME posterior median eyes;
R radix;
RA radix apophysis;
RER posterior eye row;
Sc scape;
Sp spermatheca;
St subtegulum;
T tegulum;
TA terminal apophysis;
TA I terminal apophysis I;
TA II terminal apophysis II;
TAA terminal apophysis appendix;
TE tegular extension.
The distribution map was generated with ArcGIS v. 10.5 (Environmental Systems Research Institute, Inc.).
Results and discussion
Based on traditional morphological characters and experience (matching of males and females we had hypothesized mainly on the basis of co-occurrence and compatibility of epigynes with male pedipalpal structures), all examined materials could be identified as at least two morphospecies: one belongs to an undescribed species new to science: L. guiyang sp. nov.; the other one was identified as L. fusiformis (Grasshoff 1970; Chang and Tso 2004; Tanikawa et al. 2023). However, some morphological variation is exhibited in females of L. guiyang sp. nov. Five molecular species delimitation methods were employed to test the validity of the morphology-based identification of the three Lipocrea species and the accuracy of the proposed sex matching.
The COI matrix of 17 Lipocrea individuals analyzed in this study had a sequence length of 629 bp, with 132 variable and 116 parsimony-informative sites. For COI, phylogenetic inference from BI and ML analyses yielded similar topologies with high support (Fig. 4; posterior probability, PP = 1; bootstrap value, BS = 100). The trees clearly divided the samples into four deeply divergent clades (Fig. 4).
Bayesian CO1 gene tree for 17 terminals of Lipocrea, with the results of three different species delimitation approaches. Numbers above branches show bootstrap supports and posterior probability (black), and values above show mean intraspecific (red), calculated as Lipocrea two-parameter (K2P)/uncorrected p-distance. Species names (for species codes, see Table 1) according to consensus results of species delimitation approaches.
When considering three species of Lipocrea, interspecific distances were higher than intraspecific distances. Interspecific distances range from 3.27 to 11.69% for K2P (Fig. 5A) and from 3.18 to 10.65% for uncorrected p-distance (Fig. 5B). The lowest mean interspecific distance was 12.75% / 11.51% (K2P / uncorrected p-distance) found between L. phosop and L. fusiformis, and the highest mean intraspecific distance (1.81% / 1.77% K2P / uncorrected p-distance) was estimated for L. fusiformis. The barcoding gap range identified three species, one of which is new (Fig. 4).
DNA barcoding gap for Lipocrea. Histograms show the division of intraspecific (gray) and interspecific (black) CO1 sequence variation based on the Kimura two-parameter (K2P) (A) and uncorrected p-distance (B).
The ABGD results varied based on different parameter combinations of both the initial and recursive partitions. Based on the barcoding gap results, a distinct gap was observed among the three Lipocrea species, demonstrating substantial genetic divergence. Therefore, the value of X=2 was selected as the relative gap width (Table 2). The initial three species partition was consistent with the three morphospecies, while the recursive partition regime yielded more species (Fig. 4; Table 2). However, all analyses under different assumptions revealed three hypothetical species which is entirely consistent with the morphological identification (Fig. 4).
Our P ID (Liberal) results revealed high P ID (Liberal) values > 0.95 (0.86–1.0) (Table 3), thereby also supporting the taxonomy of three putative species (Fig. 4).
The results of our mPTP analysis indicated that, when only monophyletic species are considered, our three hypothetical species were identified as anticipated (Fig. 4). The mPTP model strongly supports three species: L. guiyang sp. nov., L. phosop and L. fusiformis (PP = 1.0).
GMYC produced the same results as the four methods mentioned above (Fig. 4). The single-threshold model GMYC resulted in three clusters (confidence intervals: 1–5) and three entities (confidence intervals: 1–17) (Table 4).
**Table 4.: Results of the general mixed Yule-coalescent (GMYC) analyses (*P < 0.001).
In conclusion, all analyses support three morphological species, with terminals within the same clade being conspecific. In L. guiyang sp. nov., the results from five molecular species delimitation analyses indicate that, despite considerable morphological variation among individuals, they should be classified as the same species: (1) Both the ABGD and GMYC methods, whether using multiple parameter setting s or visualizing the data tree, support a single species (Fig. 4, Tables 2, 4); (2) these results are also supported by both the P ID (Liberal) and mPTP methods, with a P ID (Liberal) value of 1.00 (0.86, 1.0) (> 95%) (Table 3) and a posterior probability of one in mPTP; (3) our barcoding gap results (3.27% – 11.69% / 3.18% – 10.65% / K2P/uncorrected p-distance) further support that the average genetic distance (0% / 0% K2P/uncorrected p-distance) of L. guiyang sp. nov. represents the intraspecific genetic distance (Figs 4, 5). After careful re-examination of all specimens, the variation is concluded to be intraspecific differences. We also confirm sex pairing accuracy of both morphospecies based on an integrated morphological and molecular approach, including L. guiyang sp. nov. The molecular and morphological identifications are therefore congruent.
As can be concluded from the above, when identifying certain species of the genus Lipocrea, relying solely on morphological characters may lead to over-splitting of species. Nevertheless, our results indicate that even single-locus analyses based on the COI barcodes, when integrated with morphological data and collection experience, may provide sufficiently reliable species delimitation.
Taxonomic accounts
Family Araneidae Clerck, 1757
Lipocrea
Taxon classificationAnimaliaAraneaeAraneidae
Genus
Thorell, 1878
540FDC1F-ABAD-5393-91BC-A719732BC7B9
Type species.
Meta fusiformis Thorell, 1877, from Indonesia (Sulawesi), India, Myanmar, Vietnam, Japan, Bangladesh, Philippines, Thailand, China, New Guinea, Australia (Grasshoff 1970; Tanikawa 1989; Okuma et al. 1993; Barrion and Litsinger 1995; Chang and Tso 2004).
Amended diagnosis.
Lipocrea differs from Larinia sensu stricto by: the broad epigynal base with auricular lateral edges and a bipartite posterior margin (vs epigynal base not widening and posterior margin not bipartite) (cf. Figs 10C, 14D and Grasshoff 1970: fig. 6d; Gaymard and Lecigne 2018: fig. 9B; Morano 2023: figs 81, 82); conductor broadly fused with the tegular extension, forming a continuous structure (conductor separated from tegular extension) (cf. Figs 8A–C, 9A–C, E, 13B, C and Grasshoff 1970: fig. 6b, c, Gaymard and Lecigne 2018: fig. 9D, E, Morano 2023: fig. 80); median apophysis with a conspicuous spine-like or hook-shaped process with a sharp apex (vs. process absent) (cf. Figs 8A–C, 9A–C, E, 13A–C and Grasshoff 1970: fig. 6b, c, Alioua et al. 2020: 2, fig. 9, Gaymard and Lecigne 2018: fig. 9D, E, Morano 2023: fig. 80); radix with a keel-like ridge (vs no ridge) (cf. Figs 8A, C, 9A, C, E, 13A, C and Grasshoff 1970: fig. 6a, b, Alioua et al. 2020: 2, fig. 9).
Lipocrea guiyang J. Zhang, Yu & Mi, sp. nov., male holotype (YHGY208), habitus (A–C) and frontal view of cephalothorax (D). A. Dorsal; B. Ventral; C. Lateral; D. Male. Scale bars: 1 mm (A–C).
Composition and distribution.
The genus Lipocrea currently comprises five species (WSC 2025): L. diluta Thorell, 1887, distributed from Myanmar to Indonesia; L. epeiroides (O. Pickard-Cambridge, 1872), occurring from Spain and Italy (including Sardinia and Sicily) to Malta, Cyprus, Turkey, Israel, Yemen, and India; L. fusiformis (Thorell, 1877), with a distribution extending from India to Japan, the Philippines, and Indonesia (Sulawesi); L. longissima (Simon, 1881), found throughout central, eastern, and southern Africa; and L. phosop (Tanikawa, Into & Petcharad, 2023), currently known only from Thailand. This paper reports the sixth member of the genus, L. guiyang sp. nov., currently endemic to Guiyang City, China.
Based on preliminary molecular phylogenetic results (Suppl. material 1: fig. S1) and morphological evidence (see diagnosis above), L. fusiformis, L. guiyang sp. nov., and L. phosop are confidently placed in the genus Lipocrea (Figs 8–10B–F, 11, 13, 14; Tanikawa et al. 2023 figs 3–14). In addition, although not included in our molecular analyses, L. epeiroides (O. Pickard-Cambridge, 1872) and L. longissima (Simon, 1881) exhibit characteristics typical of Lipocrea and are thus justifiably assigned to this genus (Bosmans and Colombo 2015: figs 43–48; Grasshoff 1970: figs 12a, b, 14a–c). However, due to the absence of a spine-like or hook-shaped process on the median apophysis, a keel-like ridge on the radix, and a broad epigynal base in L. diluta (Grasshoff 1970: fig. 17a–c), there remains considerable uncertainty regarding the placement of this species within Lipocrea.
Lipocrea
guiyang
Taxon classificationAnimaliaAraneaeAraneidae
J. Zhang, Yu & Mi sp. nov.
958E9D71-6833-5B8E-8ABF-5E6B0CE739CE
https://zoobank.org/200B8EC9-2032-49B4-9483-91D65AC0887D
Type material.
Holotype • ♂ (YHGY208), China: Guizhou Prov.: Guiyang City, Kaiyang Co., Nanjiang Grand Canyon, 26.94°N, 106.97°E, c. 861 m, by hand, 7 VI 2022, H. Yu & Q. Lu leg. Paratypes: • 2♀ (YHGY209, YHGY428), same data as holotype.
Other material examined.
• 1♂1♀ (YHGY218, YHGY318), same data as holotype.
Diagnosis.
Males of L. guiyang sp. nov. can be easily distinguished from all other congeners with the exception of L. phosop by having a bifurcate terminal apophysis (TA); a terminal apophysis appendix (TAA) that is relatively short, with its apex not extending beyond the apex of the TA and directed distally; and a relatively long conductor (C) that is nearly as long as the tegular extension (TE) (Figs 8A–C, 9A–E; Tanikawa et al. 2023: figs 3–5, 9–11) (TA not bifurcate, TAA tongue-shaped, wrapping around TA, apex directed proximally, C rostrate and distinctly shorter than TE, such as in L. epeiroides, L. fusiformis and L. longissima as in Fig. 13A–C, Levy 1986: figs 32–34, Tanikawa 2009: figs 191, 192, Grasshoff 1970: figs 12a, b,15a, b) but can be easily differentiated from L. phosop by: (1) TA distinctly bifurcating into two apophyses (vs slightly bifurcate and not forming two processes) (cf. Figs 8B, C, 9A, B, D, E and Tanikawa et al. 2023: figs 3–5, 9–11); (2) TAA digitiform (vs papilliform) (cf. Figs 8B, C, 9B, C, E and Tanikawa et al. 2023: figs 3, 9–11); (3) process of median apophysis (HP) hook-shaped, distinctly curved (vs nearly triangular) (cf. Figs 8A–C, 9A–C, E and Tanikawa et al. 2023: figs 3–5, 9–11); (4) C shaped like a ox horn (vs finger-shaped) (cf. Figs 8A–C, 9A–C, E and Tanikawa et al. 2023: figs 3–5, 9–11). Females of L. guiyang sp. nov. also resemble those of L. phosop by the presence of a knob-shaped projection (KP) (epigyne with a scape instead of a KP in all other congeners, such as L. fusiformis; as in Fig. 14A–D), but it can be easily differentiated from L. phosop by the following features: (1) epigyne nearly trapeziform or disc-shaped in ventral view (vs inverted heart shape) (cf. Figs 10C, 11A, C and Tanikawa et al. 2023: figs 6, 12); (2) KP partly membranous and translucent (vs KP more sclerotized, non-transparent) (cf. Fig. 10C and Tanikawa et al. 2023: figs 6, 12); (3) copulatory openings (CO) located on comma-shaped or circular windows, with the posterior margin clearly separated from the posterior margin of epigyne (vs windows shaped like a horizontally oriented check mark, posterior margin of CO close to posterior margin of epigyne) (cf. Figs 10C, 11A, C and Tanikawa et al. 2023: figs 6, 12).
Description.
Male (holotype, YHGY208). Measurements. Total length 6.09. Carapace 2.70 long, 1.77 wide. Abdomen 3.55 long, 1.52 wide. Sternum 1.30 long and 0.76 wide. Labium 0.19 long and 0.39 wide. Endites 0.55 long and 0.35 wide. Clypeus height 0.10. Both margins of chelicerae with four teeth. Eye sizes and interdistances: AME 0.17, ALE 0.12, PME 0.14, PLE 0.11, AME–AME 0.20, ALE–AME 0.12, PME–PME 0.03, PME–PLE 0.26. MOQ 0.43 long, anterior width 0.47, posterior width 0.28. Leg measurements: I missing, II 12.63 (3.07, 4.55, 3.91, 1.10), III 6.22 (2.09, 2.03, 1.45, 0.65), IV 10.53 (3.11, 3.53, 3.08, 0.81).
Habitus (Fig. 6A–D). Carapace nearly oval, basically yellowish white, with a narrow red midline extending from just behind the PME almost to the black fovea; a faint dark spot present approximately at the midpoint of the midline; pars cephalica distinctly narrowed; cervical groove and radial grooves invisible; tegument smooth. AER distinctly recurved, PER nearly as wide as AER, almost straight in dorsal view. Sternum bright yellow, shield shaped; anterior margin nearly straight, posterior region strongly protruding between coxae III. Chelicerae colored as carapace, with reddish fangs. Labium and endites colored as carapace; labium nearly triangular, concave laterally; endites depressed posteriorly, slightly convergent anteriorly, with dense scopulae on inner margin. Legs uniformly colored as carapace. Abdomen elongate-oval; dorsum reddish brown, with a sword-shaped median band extending along its entire length, bordered with yellow lines and bearing a prominent large black spot anteriorly; laterally with two distinct yellow longitudinal lines, each line accompanied by approximately four small black spots; venter grayish, without distinct pattern; spinnerets yellow.
Palp (Figs 8A–D, 9A–E). Cymbium (Cy) navicular, ~2.2× longer than wide, dorsally with sparse, long setae (all detached in ethanol), basoretrolaterally with a thumb-like paracymbium (Pc). Pc moderately large, about 1/5 length of cymbium, apex blunt, slightly curved and pointing retrolatero-distally. Tegulum (T) disc-shaped, slightly wider than cymbium, with distinct sperm duct along anterior margin, proximally covered by broad subtegulum (St). Tegular extension (TE) laminar, extending dorsally, almost completely concealed by conductor (C) in ventral view. St ~1/2 cymbium length, partly membranous, surface wrinkled and ribbed, with numerous diagonal ridges. Radix (R) leaf shaped, ~½ the width of the subtegulum length, distally with a triangular apophysis (RA). RA hyaline, nearly as long as radix, apex sharp and pointing distally. Median apophysis (MA) heavily sclerotized, located prolaterally to tegulum, consisting of a broad base and a hook-shaped process (HP); base navicular, ~2/5 the width of the subtegulum in length; HP nearly as long as base, apex sharp, distinctly curved and pointing retrolaterally. Terminal apophysis (TA) hidden behind tegulum, extending distally, apex surpassing the tegulum and bifurcating into two apophyses, forming a C-shape in anterior view; both terminal apophysis I (TA I) and terminal apophysis II (TA II) heavily sclerotized, with blunt apices pointing prolaterally; TA I relatively large, its length nearly equal to the width of the tegulum; TA II smaller and humble, ~½ the length of TAI. Terminal apophysis appendix (TAA) membranous, digitiform, accompanied by terminal apophysis, hidden behind tegulum, extending distally. C originating from dorsal-anterior portion of tegulum, proximally fused to weakly sclerotized TE; tip distinctly curved, shaped like an ox horn, with a sharp apex pointing dorso-distally. Embolus (Em) spine-shaped, nearly as long as the hook-shaped process of the median apophysis, originating centrally in anterior view, extending distally, surrounded by the RA, MA, TA, and C.
Female (YHGY428). Total length 7.30. Carapace 2.83 long, 1.81 wide. Abdomen 4.96 long, 2.67 wide. Sternum 1.25 long and 0.84 wide. Labium 0.28 long and 0.49 wide. Endites 0.61 long and 0.43wide. Clypeus height 0.10. Both margins of chelicerae with four teeth. Eye sizes and interdistances: AME 0.15, ALE 0.13, PME 0.13, PLE 0.11, AME–AME 0.23, ALE–AME 0.19, PME–PME 0.02, PME–PLE 0.35. MOQ 0.44 long, anterior width 0.48, posterior width 0.27. Leg measurements: I 14.53 (3.33, 5.36, 4.54, 1.30), II 13.15 (3.26, 4.89, 3.86, 1.14), III 6.96 (2.31, 2.27, 1.58, 0.80), IV 11.80 (3.28, 4.39, 3.14, 0.99).
Habitus (Fig. 7A–C, 10A). Similar to males, but the dorsum of the abdomen lacks the prominent anterior black spot.
Lipocrea guiyang J. Zhang, Yu & Mi, sp. nov., female paratypes, YHGY428 (A–C) and YHGY209 (D–F), habitus. A, D. Dorsal; B, E. Ventral; C, F. Lateral. Scale bars: 1 mm.
Male palp of the holotype of Lipocrea guiyang J. Zhang, Yu & Mi, sp. nov. A. Dorsal; B. Ventral; C. Prolateral; D. Retrolateral. Abbreviations: C = conductor; Cy = cymbium; Em = embolus; HP = hook-shaped process of MA; MA = median apophysis; Pc = paracymbium; R = radix; RA = radix apophysis; St = subtegulum; T = tegulum; TA I = terminal apophysis I; TA II = terminal apophysis II; TAA = terminal apophysis appendix; TE = tegular extension. Scale bar: 0.2 mm.
Male palpal bulb of the holotype of Lipocrea guiyang J. Zhang, Yu & Mi, sp. nov. A. Dorsal; B. Ventral; C. Prolateral; D. Retrolateral; E. Anterior. Abbreviations: C = conductor; Em = embolus; HP = hook-shaped process of MA; MA = median apophysis; R = radix; RA = radix apophysis; St = subtegulum; T = tegulum; TA I = terminal apophysis I; TA II = terminal apophysis II; TAA = terminal apophysis appendix; TE = tegular extension. Scale bar: 0.2 mm.
Frontal view of cephalothorax (A.) and macerated epigyne (B–F) of the paratype (YHGY428) of Lipocrea guiyang J. Zhang, Yu & Mi, sp. nov. A. Female; B. Lateral; C. Ventral (blue dashed line showing the lateral and posterior margins of the epigynal base); D. Dorsal; E. Ventro-posterior; F. Dorso-anterior. Abbreviations: CD = copulatory duct; CO = copulatory opening; FD = fertilization duct; KP = knob-like projection; MS = median septum; Sp = spermatheca. Scale bars: 1 mm (A), 0.2 mm (B–F).
Epigyne of the paratype (YHGY209) of Lipocrea guiyang J. Zhang, Yu & Mi, sp. nov. A. Intact, ventral; B. Macerated, lateral; C. Macerated, ventral; D. Macerated, dorsal; E. Macerated, ventro-posterior; F. Macerated, dorso-anterior. Abbreviations: CD = copulatory duct; CO = copulatory opening; FD = fertilization duct; KP = knob-like projection; MS = median septum; Sp = spermatheca. Scale bar: 0.2 mm.
Genitalia (Fig. 10B–F). Epigyne strongly sclerotized with large postero-lateral lobes, distinctly wider than long, nearly trapeziform in ventral view and inverted triangular in lateral view. Knob-shaped projection (KP) represented by a small, partly membranous tubercle, located at anterior portion of epigynal plate. Copulatory openings (CO) large, located on the comma-shaped window (or pockets with chitinous posterior margins) is at the postero-lateral portion of the epigynal plate, separated by indistinct median septum (MS). Copulatory ducts (CD) short, diverging and ascending obliquely, forming V-shaped course in dorsal view, finally entering anteriorly located spermathecae. Spermathecae (Sp) oval, ~1.2× longer than wide, relatively large, ~2/3 of epigyne length; two spermathecae close together, separated by ~2/3 of their width. Fertilization ducts (FD) membranous, relatively long, ~2/3 of spermathecae length, located on dorsal-basal surface of spermathecae.
Distribution.
Known only from the type locality (Fig. 1).
Etymology.
The specific epithet is derived from the name of the type locality; noun in apposition.
Comments.
In spite of the stable morphology of the male palp and the consistent coloration of the male habitus, considerable morphological variation is observed among female individuals, primarily related to epigynal structures. These variations involve features such as the presence or absence of a knob-shaped projection (KP), the shape of the copulatory openings (CO), and whether the median septum (MS) is distinct or indistinct. For example, in some females (e.g., YHGY428, as in Fig. 10B, C, E), the KP is distinct, the CO is situated on a comma-shaped window (or within pockets with chitinous posterior margins), and the MS is indistinct. In contrast, in other individuals (e.g., YHGY209, as in Fig. 11A–C, E), the KP is broken off, the CO is positioned on a nearly circular window (or within pockets bordered anteriorly, internally, and posteriorly), and the MS is distinct. In addition, some variation related to the abdominal pattern is also observed: the dorsum of the abdomen bears a median band extending along its entire length in some individuals (e.g., YHGY428), whereas in others (e.g., YHGY209), the median band is restricted to the posterior quarter of the dorsum (cf. Fig. 7A and Fig. 7D). However, the morphological variation was determined to be intraspecific variation based on the molecular species delimitation analysis.
Lipocrea
fusiformis
Taxon classificationAnimaliaAraneaeAraneidae
(Thorell, 1877)
79C3FD6A-E5F2-5664-B148-B1F22769C556
Meta fusiformis Thorell, 1877: 431 (♀). Lipocrea fusiformis : Thorell 1878: 6. Larinia quadrinotata Simon, 1889: 340 (juv.); Simon 1909: 105 (♀). Larinia lutescens Thorell, 1898: 342 (♂♀). Larinopa fusiformis : Grasshoff 1970: 231, figs 15a, b, 16a–e (♂♀, transfer from Larinia, synonym of Larinia lutescens and L. quadrinotata); Tanikawa 1989: 35, figs 8–14 (♂♀); Chang and Tso 2004: 28, figs 5–8 (♂♀); Tanikawa et al. 2023: 56, figs 15–18.
Note.
For full list of taxonomic references, see WSC (2025).
Material examined.
• 1♂, 3♀ (YHGY432, YHGY495, YHGY507, YHGY508), China: Guizhou Prov.: Guiyang City, Huaxi District, University Town, 26.38°N, 106.65°E, c. 1173 m, by beating, 7 VII 2022, Q. Jiang & Q. Du leg; • 1♂, 1♀ (YHGY431, YHGY433), Guiyang City, Huaxi District, Dangwu Town, 26.38°N, 106.60°E, c. 1138 m, by hand, 19 V 2022, H. Yu & Q. Lu leg; Guiyang City, Kaiyang Co., Nanjiang Grand Canyon, 26.94°N, 106.97°E, c. 861 m, by hand, 7 VI 2022, H. Yu & Q. Lu leg.
Diagnosis and description.
See Tanikawa (1989). Living specimens as in Fig. 3A–C, male habitus as in Fig. 12A–C, male palp as in Fig. 13A–D, female habitus as in Fig. 12D–F, genitalia as in Fig. 14A–F.
Lipocrea fusiformis (Thorell, 1877), male (YHGY431, A–C) and female (YHGY432, D–F), habitus. A, D. Dorsal; B, E. Ventral; C, F. Lateral. Scale bars: 1 mm (A–C), 2 mm (D–F).
Male palp of Lipocrea fusiformis (Thorell, 1877). A. Dorsal; B. Ventral; C. Prolateral; D. Retrolateral. Abbreviations: C = conductor; Cy = cymbium; Em = embolus; HP = hook-shaped process of MA; MA = median apophysis; Pc = paracymbium; R = radix; RA = radix apophysis; St = subtegulum; T = tegulum; TA = terminal apophysis; TAA = terminal apophysis appendix; TE = tegular extension. Scale bar: 0.2 mm.
Macerated epigynes of females of Lipocrea fusiformis (Thorell, 1877), YHGY432 (A–C) and YHGY507 (D–F). A. Ventral, with scape; B. Lateral, with scape; C. Ventro-posterior, with scape; D. Ventral, without scape (blue dashed line showing the lateral and posterior margins of the epigynal base); E. Dorsal, without scape; F. Ventro-posterior, without scape. Abbreviations: CD = copulatory duct; CO = copulatory opening; FD = fertilization duct; MS = median septum; Sc = scape; Sp = spermatheca. Scale bars: 0.2 mm (A, B, D–F).
Distribution.
China (Guizhou, new record for mainland China; Taiwan), India, Burma, Vietnam, Thailand, Japan, Philippines, Sulawesi, New Guinea, Australia.
Comments.
Among all examined female specimens, the epigynal scape is broken off in some individuals (e.g., YHGY507, as in Fig. 14D–F), but it remains intact in others (e.g., YHGY432, as in Fig. 14A–C). Aside from this difference, all other morphological features are consistent. In addition, results from molecular evidence confirm that these specimens belong to the same species (Fig. 4; for details, see the results and discussion section).
Supplementary Material
XML Treatment for Lipocrea
XML Treatment for Lipocrea guiyang
XML Treatment for Lipocrea fusiformis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alioua Y Bosmans R Kherbouche O Bissati S (2020) Distribution of the genus Larinia in the Maghreb, with the first record of Larinia chloris in Algeria (Araneae: Araneidae).Arachnologische Mitteilungen 59(1): 1–4. 10.30963/aramit 5901 · doi ↗
- 2Barrett RDH Hebert PDN (2005) Identifying spiders through DNA barcodes.Canadian Journal of Zoology 83(3): 481–491. 10.1139/z 05-024 · doi ↗
- 3Barrion AT Litsinger JA (1995) Riceland Spiders of South and Southeast Asia.CAB International, Wallingford, 700 pp. [16 pls.]
- 4Bosmans R Colombo M (2015) New species of spiders from Sardinia (Araneae), with ecological notes on Lipocrea epeiroides (O. Pickard-Cambridge, 1872) (Araneae: Araneidae).Arachnology 16(9): 319–332. 10.13156/arac.2015.16.9.319 · doi ↗
- 5Bouckaert R Heled J Kuhnert D Vaughan T Wu CH Xie D Suchard MA Rambaut A Drummond AJ (2014) BEAST 2: A software platform for Bayesian evolutionary analysis. P Lo S Computational Biology 10(4): e 1003537. 10.1371/journal.pcbi.1003537 PMC 398517124722319 · doi ↗ · pubmed ↗
- 6Chang YH Tso IM (2004) Six newly recorded spiders of the genera Araneus, Larinia, Eriophora, Thanatus, Portia and Dolichognatha (Araneae: Araneidae, Philodromidae, Salticidae and Tetragnathidae) from Taiwan.Acta Arachnologica 53(1): 27–33. 10.2476/asjaa.53.27 · doi ↗
- 7Cheng H Chen JY (2022) Guiyang: City of a thousand parks, city of forests.Society and Public Welfare 8: 25–27.
- 8Darriba D Taboada GL Doallo R Posada D (2012) j Model Test 2: More models, new heuristics and parallel computing. Nature Methods 9(8): e 772. 10.1038/nmeth.2109 PMC 459475622847109 · doi ↗ · pubmed ↗