Morphological and phylogenetic analyses reveal two new species of Rhodoveronaea (Rhamphoriaceae, Rhamphoriales) from China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
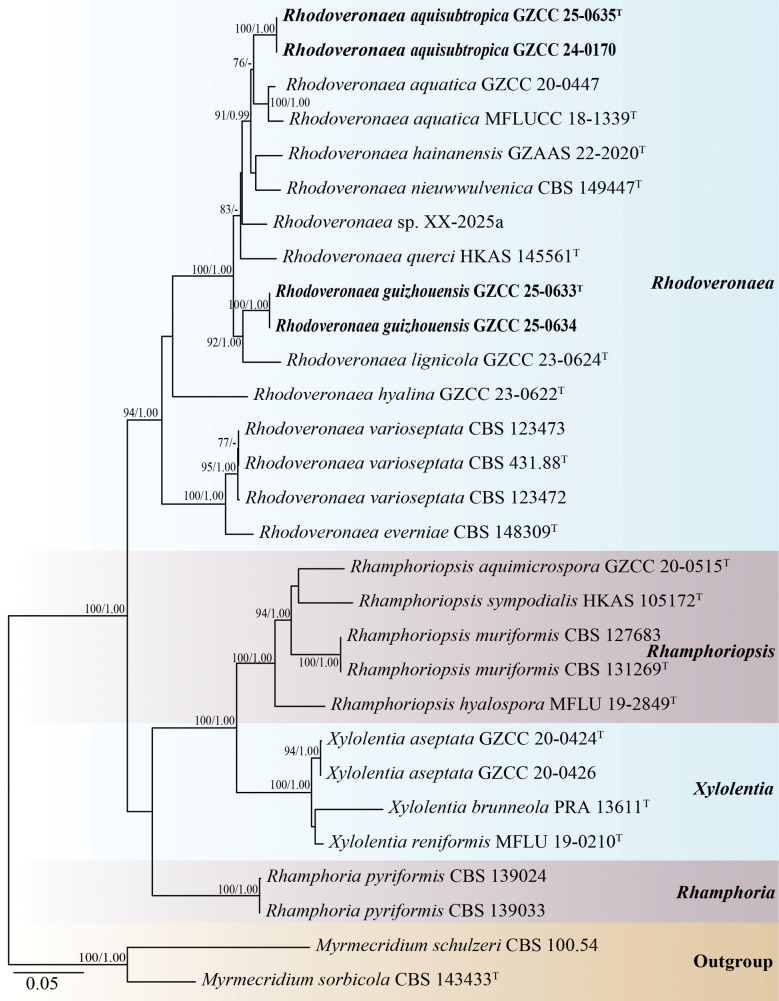 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3| Taxon | Strain | GenBank Accessions | ||||
|---|---|---|---|---|---|---|
|
|
|
|
|
| ||
|
| CBS 100.54 |
|
| - | - | - |
|
| CBS 143433T |
|
| - | - | - |
|
| CBS 139033 |
|
|
| - |
|
|
| CBS 139024 |
|
|
| - |
|
|
|
|
|
|
|
| |
|
| MFLU 19-2849T |
|
| - | - | - |
|
| CBS 127683 |
|
|
| - |
|
|
| CBS 131269T |
| - |
| - |
|
|
| HKAS 105172T |
|
| - | - | - |
|
| MFLUCC 18-1339T |
|
|
|
| - |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
| CBS 148309T |
|
| - | - |
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
|
|
| - | - | - | |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
| CBS 149447T |
|
| - |
|
|
|
| HKAS 145561T |
|
|
|
| - |
| XX-2025aT | - |
|
| - | - | |
|
| CBS 431.88T |
|
| - | - | - |
|
| CBS 123472 |
|
|
| - |
|
|
| CBS 123473 |
|
|
| - |
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
| PRA 13611T |
|
|
| - |
|
|
| MFLU 19-0210T |
|
| - | - | - |
| Species | Distribution | Habitat | Reference |
|---|---|---|---|
|
| China | Freshwater | |
|
| China | Freshwater | This study |
|
| Netherlands | Terrestrial |
|
|
| China | Freshwater | This study |
|
| China | Freshwater |
|
|
| China | Terrestrial |
|
|
| China | Terrestrial |
|
|
| Netherlands | Terrestrial |
|
|
| China | Terrestrial |
|
|
| Czech Republic, France, Sweden | Terrestrial | Arzanlou1 et al. (2007); |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInvertebrate Taxonomy and Ecology · Mollusks and Parasites Studies · Evolution and Paleontology Studies
Introduction
Freshwater fungi represent a taxonomically and ecologically diverse group of organisms that are widely distributed across various geographical regions, particularly in China (Guizhou, Hainan, Hong Kong, Taiwan, and Yunnan) and northern Thailand (Chiang Mai and Chiang Rai) (Zhang et al. 2011; Hyde et al. 2016, 2021; Su et al. 2016; Bao et al. 2018, 2019, 2020, 2021, 2023; Luo et al. 2019; Dong et al. 2020, 2021a, b; Fryar and Catcheside 2023; Fryar et al. 2023; Shen et al. 2024). They play a crucial role in the decomposition of organic matter and nutrient cycling within freshwater ecosystems (Luo et al. 2004; Hyde et al. 2016; Calabon et al. 2022, 2023). These fungi typically colonize fully or partially submerged woody substrates in a variety of freshwater habitats, including ponds, lakes, rivers, streams, and swamps, as well as artificial environments such as pools, reservoirs, dams, drainage ditches, and water-cooling towers (Lu et al. 2018; Xiao et al. 2023; Yang et al. 2023; Ma et al. 2024; Wang et al. 2024a, b; Xu et al. 2025).
Rhodoveronaea was established by Arzanlou et al. (2007) with R. varioseptata as the type species. Rhodoveronaea species are primarily known from their asexual morphs, which are characterized by reddish-brown, septate, straight or flexuous conidiophores with inflated basal cells, terminally integrated conidiogenous cells with crowded, slightly conspicuous conidium-bearing denticles, pale brown, ellipsoidal to obovoidal and septate conidia with a protruding base and a marginal basal frill (Arzanlou et al. 2007; Réblová 2009; Luo et al. 2019; Crous et al. 2021, 2023; Hyde et al. 2023; Chen et al. 2024; Yang et al. 2025). The first sexual morph characterized by immersed ascomata with subglobose to conical venter bearing conical neck, filamentous, septate paraphyses longer than asci, unitunicate, cylindrical asci with long-stipitate and fusiform, septate, hyalina ascospores (Réblová 2009). Currently, eight species are accepted within the genus Rhodoveronaea, which occur in both freshwater and terrestrial habitats (Chen et al. 2024; Yang et al. 2025).
In this study, four hyphomycetes isolates were obtained from freshwater habitats in Chishui River, Guizhou Province, China. Based on morphological characteristics, illustrations, and multi-gene phylogenetic analyses, two novel species are introduced, namely, Rhodoveronaea aquisubtropica and R. guizhouensis.
Materials and methods
Sample collection, examination, and isolation
Samples of submerged decaying wood pieces were collected from the Chishui River, Chishui City, Guizhou Province in southern China. Samples were taken to the laboratory in plastic bags with the collection details, including localities and dates (Rathnayaka et al. 2024). Fresh specimens were incubated in zip-lock bags and sterile, moist plastic boxes at room temperature for two weeks. The microscopic features were examined and photographed using a stereomicroscope (SMZ-168, Nikon, Japan) and an ECLIPSE Ni compound microscope (Nikon, Tokyo, Japan) with a Canon 90D digital camera. Measurements were made using Tarosoft (R) Image Frame Work software. Photo-plates were assembled using Adobe Photoshop CC 2019 (Adobe Systems, USA).
Single spore isolations were performed on PDA plates following the methods described by Chomnunti et al. (2014) and Senanayake et al. (2020), and the germinated conidia were aseptically transferred to fresh PDA plates. Morphological characters of fungal mycelium, including color, shape, and size, were documented. Dried fungal specimens were deposited in the Herbarium of Guizhou Academy of Agriculture Sciences (Herb. GZAAS), Guiyang, China. Pure cultures were deposited in the Guizhou Culture Collection, China (GZCC), Guiyang, China. The MycoBank numbers were obtained as described in https://www.mycobank.org/.
DNA extraction, PCR amplification, and sequencing
Fresh fungal mycelia were scraped from colonies grown on PDA plates and transferred to a 1.5 mL microcentrifuge tube using a sterilized lancet for genomic DNA extraction. Genomic DNA was extracted using the Biospin Fungus Genomic DNA Extraction Kit (BioFlux, China). LR0R/LR5,ITS5/ITS4, NS1/NS4, EF1-983F/EF1-2218R, and fRPB2-5F/fRPB2-7cR were employed to amplify large ribosomal subunit (LSU; Vilgalys and Hester 1990), the internal transcribed spacer (ITS; White et al. 1990), small subunit of the nuclear ribosomal DNA (SSU, Rehner and Samuels 1994), translation elongation factor 1-α (tef1-α; Rehner and Buckley 2005), and RNA polymerase II second largest subunit (rpb2; Liu et al. 1999) sequence fragments, respectively. DNA preparation was conducted in a 50 μL mixture, which included 2 μL ofDNA, 2 μL of each forward and reverse primer, and 44 μL of 1.1 × T3 Supper PCR Mix (Qingke Biotech, Chongqing, China). The conditions for the polymerase chain reaction (PCR) correspond to those reported by Chen et al. (2024). The PCR products were purified and sequenced with the same primers at Beijing Tsingke Biotechnology Co., Ltd.
Phylogenetic analyses
The newly obtained sequences were checked and assembled using BioEdit v.7.0.5.3 (Hall 1999) and SeqMan v.7.0.0 (DNASTAR, Madison, WI, USA; Swindell and Plasterer 1997), respectively. The sequences incorporated in this study were downloaded from GenBank (Table 1; https://www.ncbi.nlm.nih.gov/). Multiple sequences were aligned using MAFFT v.7.473 (https://mafft.cbrc.jp/alignment/server/; Katoh et al. 2019). The dataset was trimmed using trimAl v.1.2rev59 software (Capella-Gutiérrezet et al. 2009). A combined sequence dataset was created using SequenceMatrix-Windows-1.7.8 software (Vaidya et al. 2011).
Maximum likelihood (ML) analysis was carried out using the IQ Tree online website (http://iqtree.cibiv.univie.ac.at/) based on Bayesian Information Criteria (BIC) (Nguyen et al. 2015). The substitution model was automatically tested by the server. Bayesian Inference (BI) analysis was performed by using MrBayes on XSEDE (3.2.7a) via CIPRES (Stamatakis 2014). The aligned fasta file was converted to the nexus format file by using AliView (Daniel et al. 2010). The best-fit evolutionary model for individual dataset was determined using MrModeltest v. 2.3. 10 (Nylander et al. 2008). The GTR+G+I substitution model was selected for LSU, tef1-α, and rpb2, whereas the SYM+I+G and HKY+G models were applied to ITS and SSU, respectively.The posterior probabilities (BYPP) were determined based on Bayesian Markov chain Monte Carlo (BMCMC) sampling (Huelsenbeck and Ronquist 2001). Five simultaneous Markov chains were run for 10,000,000 generations, and trees were sampled every 1,000^th^ generation. The burn-in phase was set at 25%, and the remaining trees were used for calculating posterior probabilities (BYPP).
Trees were visualized using FigTree v.1.4.4 and edited with Adobe Illustrator CC 2019 (v.23.1.0; Adobe Systems, USA).
Results
The phylogenetic placements of the four new strains were determined by multi-locus phylogenetic analysis. The concatenated sequence matrix comprised 4,318 characters (LSU: 1–860,ITS: 861–1,402, SSU: 1,403–2,409, tef1-α: 2,410–3,309, and rpb2: 3,310–4,318) across 29 taxa. Base frequencies and rates were A = 0.251521, C = 0.250418, G = 0.274699, and T = 0.223362; substitution rates were AC = 1.428534, AG = 2.806428, AT = 1.229261, CG = 1.263192, CT = 7.729689, and GT = 1.000000. The distribution shape parameter α equaled 0.144696. Fig. 1 presents the best-scoring ML tree, which had a final log-likelihood value of -16,381.027.
Phylogenetic tree generated from the RAxML analysis based on the combined LSU, ITS, SSU, tef1-α, and rpb2 sequence data. Bootstrap support values of RAxML (ML) equal to or greater than 75%, and Bayesian posterior probabilities (PP) equal to or greater than 0.95 are given near the nodes as ML/BYPP, respectively. The Maximum Likelihood (ML) and Bayesian Inference (BI) analyses yielded similar tree topologies. Hyphen (“-”) indicates a posterior probability lower than 0.95 for Bayesian.Myrmecridium schulzeri (CBS 100.54) and M. sorbicola (CBS 143433) were selected as outgroups. Ex-type strains are denoted with “T” and newly obtained isolates are in bold black fonts.
Based on the multi-gene phylogenetic tree (Fig. 1), our collections represent two species of Rhodoveronaea within the family Rhamphoriaceae. Our isolates, GZCC 24-0170 and GZCC 25-0635, group together and this clade forms a distinct lineage with Rhodoveronaea aquatica (GZCC 20-0447 and MFLUCC 18-1339) with 76% ML support. In addition, GZCC 25-0633 and GZCC 25-0634 form a clade together and are sister to Rhodoveronaea lignicola (GZCC 23-0624) with 92% ML and 1.00BYPP support.
Taxonomy
Rhodoveronaea
aquisubtropica
Taxon classificationFungiRhamphorialesRhamphoriaceae
H. Zhang & J. Ma sp. nov.
CD56C901-161C-5128-B95F-D4A03F0794A6
904169
Etymology.
“aqui-’’ refers to aquatic habitat of this fungus, and ‘‘-subtropica’’ means the climate type where the fungus was collected.
Holotype.
GZAAS 25-0665.
Description.
Saprobic on decaying wood in a freshwater habitat. Sexual morph Undetermined. Asexual morph Hyphomycetous. Colonies on wood effuse, hairy, scattered or aggregated, brown. Mycelium partly superficial, partly immersed, composed of branched, septate, guttulate, smooth-walled, hyalina to brown hyphae. Conidiophores 204–270 × 4.9–7 μm (x̄ = 241.5 × 6 μm, n = 20), macronematous, mononematous, erect, flexuous, solitary, cylindrical, smooth-walled, septate, unbranched, black brown, mid brown, paler towards the apex. Conidiogenous cells polyblastic, integrated, terminal, determinate, sympodial, forming a rachis with subdenticulate loci, flexuous, pale brown to subhyalina, pigmented, with inconspicuous denticles. Conidia 10.5–13.5 × 4–5.5 μm (x̄ = 12.5 × 4.7 μm, n = 25), acropleurogenous, aggregated in slimy masses, ellipsoidal to narrowly obovoid, 1–3-septate, mostly 3-septate, smooth-walled, pale yellowish brown, guttulate, thin-walled, sometimes slightly constricted at the septa.
Rhodoveronaea aquisubtropica (GZAAS 25-0665, holotype). a, b. Colonies on the host surface; c–f. Conidiophores, conidiogenous cells and conidia; g, h. Conidiogenous cells and conidia; i–l. Conidia; m. Germinated conidium; n, o. Colonies on PDA from above and below after 36 days of incubation at room temperature. Scale bars: 50 μm (c–f); 20 μm (g, h); 5 μm (i–m).
Culture characteristics.
Conidia germinating on PDA within 12 hours, producing germ tubes from the conidial body. Colonies on PDA are circular with a raised surface and entire margin, reaching 30 mm in diameter after 36 days at room temperature (approximately 25 °C), and are brown to reddish brown in color on both the surface and reverse sides.
Material examined.
China • Guizhou Province, Chishui City, Chishui River, on decaying submerged wood in a freshwater habitat, 10 October 2024, Hong Zhang & Jian Ma, CS12 (GZAAS 25-0665, holotype), ex-type living cultures GZCC 25-0635; Ibid., CS18 (GZAAS 24-0083, paratype), living culture GZCC 24-0170.
Notes.
In our phylogenetic tree (Fig. 1), our isolates (GZCC 25-0635 and GZCC 24-0170) formed a sister clade to Rhodoveronaea aquatica (GZCC 20-0447 and MFLUCC 18-1339) with 76% ML support. A comparison of LSU, ITS, SSU, and tef1-α sequences between Rhodoveronaea aquatica (MFLUCC 18-1339) and R. aquisubtropica (GZCC 25-0635) and reveals their nucleotide differences of 3/774 bp (0.4%, without gap),20/502 bp (4%, including 4 gaps), 5/852 bp (0.6%, including 3 gaps), and 29/882 bp (3.3%, without gap), respectively, indicating that they are distinct species. Morphologically, Rhodoveronaea aquisubtropica (GZAAS 25-0665) differs from R. aquatica (MFLU 18-1593, ex-type) by its narrower conidiophores (204–270 × 4.9–7 μm*vs.*182–310 × 9–13 μm), and smaller conidia (10.5–13.5 × 4–5.5 μm vs. 23–27 × 9–11 μm) (Luo et al.2019). Therefore, we introduce Rhodoveronaea aquisubtropica as a novel species based on the multi-gene phylogenetic analysis and morphological differences.
Rhodoveronaea
guizhouensis
Taxon classificationFungiRhamphorialesRhamphoriaceae
H. Zhang & J. Ma sp. nov.
029CFEE4-3E6E-58A2-A1EC-243B40E6BD88
904170
Etymology.
The specific epithet ‘guizhouensis’ refers to the locality “Guizhou Province”, from where the holotype was collected.
Holotype.
GZAAS 25-0663.
Description.
Saprobic on decaying submerged wood in a freshwater habitat. Sexual morph Undetermined. Asexual morph Hyphomycetous. Colonies on wood effuse, hairy, scattered or aggregated, brown. Mycelium partly superficial, partly immersed, composed of branched, septate, guttulate, smooth-walled, hyalina to brown hyphae. Conidiophores 211–268 × 4.5–6.3 μm (x̄ = 234 × 5.2 μm, n = 25), macronematous, mononematous, erect, flexuous, solitary, cylindrical, smooth-walled, septate, unbranched, black brown, mid brown, paler towards the apex. Conidiogenous cells polyblastic, integrated, terminal, determinate, sympodial, forming a rachis with subdenticulate loci, flexuous, pale brown to subhyalina, pigmented, with inconspicuous denticles. Conidia 11.5–17 × 4.3–6 μm (x̄ = 13.8 × 5.2 μm, n = 25), acropleurogenous, aggregated in slimy masses, ellipsoidal to narrowly obovoid, 1–3-septate, mostly 3-septate, smooth-walled, pale yellowish brown, guttulate, thin-walled, sometimes slightly constricted at the septa.
Culture characteristics.
Conidia germinating on PDA within 11 hours, producing germ tubes from the conidial body. Colonies on PDA are circular with a raised surface and entire margin, reaching 27 mm in diameter after 32 days at room temperature (approximately 25 °C), and are white, reddish brown to black in color on both the surface and reverse sides.
Rhodoveronaea guizhouensis (GZAAS 25-0663, holotype). a, b. Colonies on the host surface; c–f. Conidiophores, conidiogenous cells, and conidia; g–i. Conidiogenous cells and conidia; j. A germinated conidium; k–o. Conidia; p, q. Colonies on PDA from above and below after 32 days of incubation at room temperature. Scale bars: 60 μm (c–f); 30 μm (g, h); 20 μm (i); 10 μm (j); 5 μm (k–o).
Material examined.
China • Guizhou Province, Chishui City, Chishui River, on decaying submerged wood in a freshwater habitat, 10 October 2024, Hong Zhang &Jian Ma, CSF2 (GZAAS 25-0663, holotype), ex-type living cultures GZCC 25-0633; Ibid., CSF9 (GZAAS 25-0664, paratype), living culture GZCC 25-0634.
Notes.
In our phylogenetic tree (Fig. 1), our isolates (GZCC 25-0633 and GZCC 25-0634) formed a sister clade to Rhodoveronaea lignicola (GZCC 23-0624) with 92% ML and 1.00BYPP support. Rhodoveronaea guizhouensis (GZAAS 25-0663) can be distinguished from R. lignicola (GZAAS 23-0612) by its longer conidiophores (211–268 μm*vs.75–125 μm), longer conidia (up to 17 μmvs.*9–13.5 μm) (Chen et al. 2024). Moreover, base pair comparison of Rhodoveronaea guizhouensis (GZCC 25-0633) and R. lignicola (GZCC 23-0624) shows 30/528 bp differences in ITS (5.7%, gaps 5 bp), 7/862 bp differences in LSU (0.8%, gaps 2 bp), 3/910 bp differences in SSU (0.3%, without gap), 39/887 bp differences in tef1-α (4.4%, without gap), and 69/803 bp differences in rpb2 (8.6%, without gap). Therefore, based on DNA molecular data and morphological characteristics, we introduce Rhodoveronaea guizhouensis as a novel species.
Discussion
The genus Rhodoveronaea currently comprises ten species, including the two new species described in the present study (Arzanlou et al. 2007; Luo et al. 2019; Crous et al. 2021, 2023; Hyde et al. 2023; Chen et al. 2024; Yang et al. 2025). Among them, four species are found in freshwater habitats and six in terrestrial habitats (Table 2), with primary distributions in Guizhou, Yunnan, and Hainan provinces of China.Rhodoveronaea species are distributed in China, Czech Republic, France, Germany, Netherlands, and Sweden (Arzanlou et al. 2007; Réblová 2009; Luo et al. 2019; Crous et al. 2021, 2023; Hyde et al. 2023; Chen et al. 2024; Yang et al. 2025). They occur as saprobes on bamboo stick, Bertia moriformis, Carpinus betulus, Evernia prunastri, and decaying wood in both freshwater and terrestrial habitats (Arzanlou et al. 2007; Réblová 2009; Luo et al. 2019; Crous et al. 2021, 2023; Hyde et al. 2023; Chen et al. 2024; Yang et al. 2025).
Based on multi-gene phylogenetic analyses, considerable morphological variation can occur even within the same Rhodoveronaea species. For example, two collections (MFLU 18-1593 from decaying submerged wood in Yunnan Province and HKAS 112574 from a similar substrate in Guizhou Province, China) have both been identified as the same species, namely Rhodoveronaea aquatica (Luo et al. 2019; Yang et al. 2023). However, MFLU 18-1593 exhibits significantly larger conidiophores (182–310 × 9–13 μm) and longer conidia (23–27 × 9–11 μm) compared to those of HKAS 112574 (conidiophores: 70–147 × 3–4.5 μm; conidia: 10–13 × 4–6 μm) (Luo et al. 2019; Yang et al. 2023).
The two Rhodoveronaea species, R. aquisubtropica and R. guizhouensis, isolated from the Chishui River, exhibit highly similar morphological characteristics. However, multi-gene phylogenetic analyses (Fig. 1) provide strong support for their recognition as distinct species. This divergence may be attributed to molecular evolutionary processes, during which the two species independently developed convergent morphological traits in response to comparable environmental conditions. These findings highlight the limitations of morphology-based taxonomy alone and emphasize the importance of molecular data for accurate species delimitation. Moreover, this study underscores the potential for discovering novel fungal taxa in riverine ecosystems, particularly in ecologically rich yet underexplored regions.
Supplementary Material
XML Treatment for Rhodoveronaea aquisubtropica
XML Treatment for Rhodoveronaea guizhouensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arzanlou M Groenewald JZ Gams W Braun U Shin HD Crous PW (2007) Phylogenetic and morphotaxonomic revision of Ramichloridium and allied genera.Studies in Mycology 58: 57–93. 10.3114/sim.2007.58.0318490996 PMC 2104745 · doi ↗ · pubmed ↗
- 2Bao D Luo Z Liu J Bhat D Sarunya N Li W Su H Hyde K (2018) Lignicolous freshwater fungi in China III: Three new species and a new record of Kirschsteiniothelia from northwestern Yunnan Province.Mycosphere 9: 755–768. 10.5943/mycosphere/9/4/4 · doi ↗
- 3Bao D Su H Maharachchikumbura S Liu J Nalumpang S Luo Z Hyde K (2019) Lignicolous freshwater fungi from China and Thailand: Multi-gene phylogeny reveals new species and new records in Lophiostomataceae.Mycosphere 10: 1080–1099. 10.5943/mycosphere/10/1/20 · doi ↗
- 4Bao DF Mc Kenzie EHC Bhat DJ Hyde KD Luo ZL Shen HW Su HY (2020) Acrogenospora (Acrogenosporaceae, Minutisphaerales) appears to be a very diverse genus. Frontiers in Microbiology 11: 1606. 10.3389/fmicb.2020.01606 PMC 739373732793142 · doi ↗ · pubmed ↗
- 5Bao DF Hyde KD Mc Kenzie EHC Jeewon R Su HY Nalumpang S Luo ZL (2021) Biodiversity of lignicolous freshwater hyphomycetes from China and Thailand and description of sixteen species.Journal of Fungi 7(8): 669. 10.3390/jof 708066934436208 PMC 8399276 · doi ↗ · pubmed ↗
- 6Bao DF Hyde KD Maharachchikumbura SSN Perera RH Thiyagaraja V Hongsanan S Wanasinghe DN Shen HW Tian XG Yang LQ Nalumpang S Luo ZL (2023) Taxonomy, phylogeny and evolution of freshwater Hypocreomycetidae (Sordariomycetes).Fungal Diversity 121: 1–94. 10.1007/s 13225-023-00521-8 · doi ↗
- 7Calabon MS Hyde KD Jones EBG Luo ZL Dong W Hurdeal VG Gentekaki E Rossi W Leonardi M Thiyagaraja V Lestari AS Shen HW Bao DF Boonyuen N Zeng M (2022) Freshwater fungal numbers.Fungal Diversity 114: 3–235. 10.1007/s 13225-022-00503-2 · doi ↗
- 8Calabon MS Hyde KD Jones EBG Bao DF Bhunjun CS Phukhamsakda C Shen HW Gentekaki E Al Sharie AH Barros J Chandrasiri KSU Hu DM Hurdeal VG Rossi W Valle LG Zhang H Figueroa M Raja HA Seena S Song HY Dong W El-Elimat T Leonardi M Li Y Li YJ Luo ZL Ritter CD Strongman DB Wei MJ Balasuriya A (2023) Freshwater fungal biology.Mycosphere 14: 195–413. 10.5943/mycosphere/14/1/4 · doi ↗