Membrane Palmitoylated Protein 7 is Required for Osteogenesis and is Linked with Bone Mineralization and Osteoporosis: The Functional Evaluation of GEFOS GWAS Hit
Petra Malavašič, Jasna Lojk, Marija Nika Lovšin, Radko Komadina, Gregor Haring, Rihard Trebše, Fernando Rivadeneira, David Karasik, Barbara Ostanek, Janja Marc

TL;DR
This study shows that MPP7, a protein linked to bone health, is important for bone formation and its absence can lead to osteoporosis.
Contribution
The study identifies MPP7 as a critical gene for osteogenesis and links it to osteoporosis through human and zebrafish data.
Findings
MPP7 expression is reduced in osteoporotic bone tissue.
MPP7 knockout in HOS cells impairs osteogenic differentiation and mineralization.
MPP7 influences osteoblast function through cell polarity and adhesion.
Abstract
Genome-wide association studies have identified multiple loci associated with bone mineral density, a major determinant of osteoporotic fracture risk. At one such locus, genetic, bioinformatic, and zebrafish knockout data strongly prioritize membrane palmitoylated protein 7 (MPP7) as a candidate gene, although its precise role in bone biology remains poorly defined. MPP7 encodes a member of the p55 Stardust family of membrane-associated guanylate kinase proteins, which are key regulators of epithelial cell polarity and junctional organization. Here, we investigated the functional role of MPP7 in bone biology. We found that MPP7 expression was significantly reduced—by approximately twofold—in bone tissue from osteoporotic patients compared with osteoarthritic patients and non-osteoporotic controls. Furthermore, we generated a CRISPR/Cas9-mediated MPP7 knockout in the human osteosarcoma…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
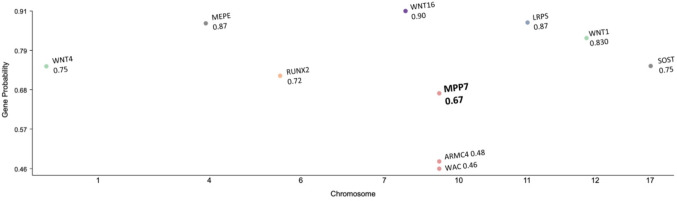 Figure 1
Figure 1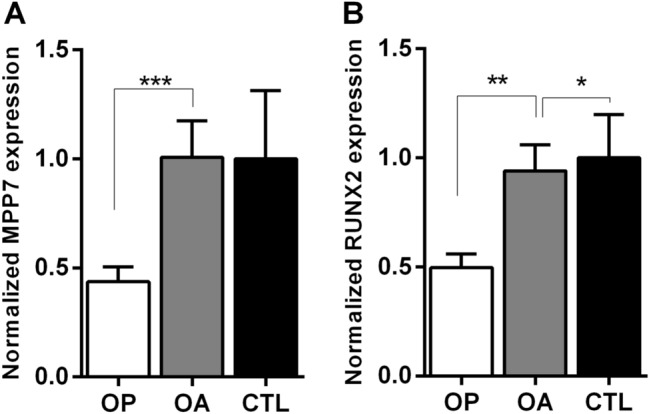 Figure 2
Figure 2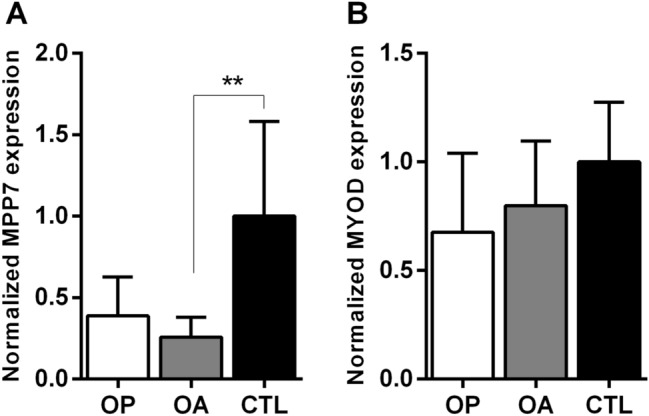 Figure 3
Figure 3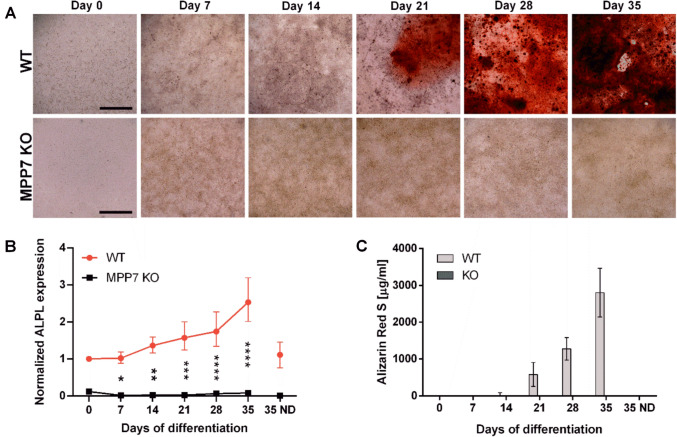 Figure 4
Figure 4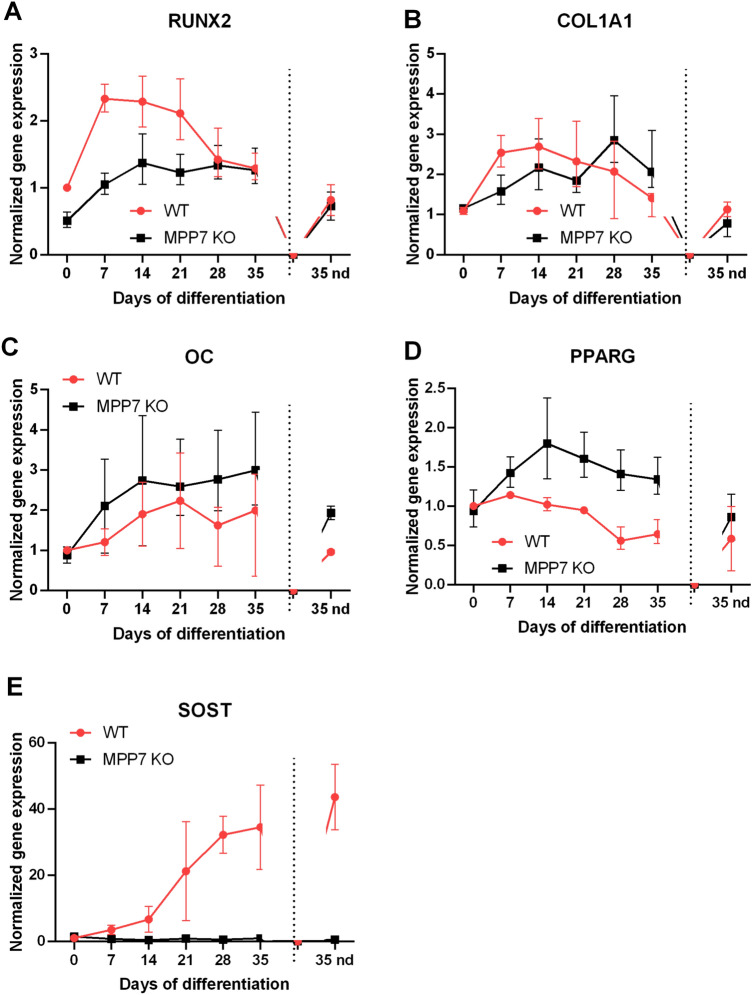 Figure 5
Figure 5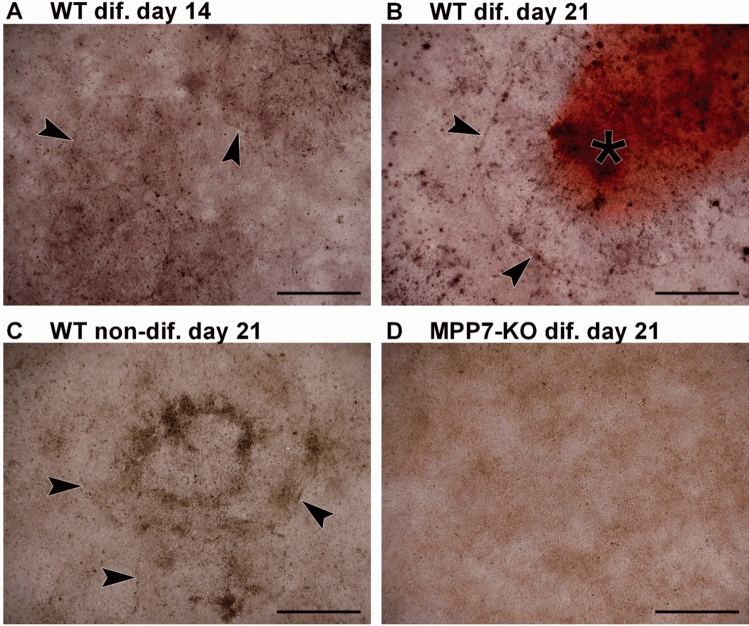 Figure 6
Figure 6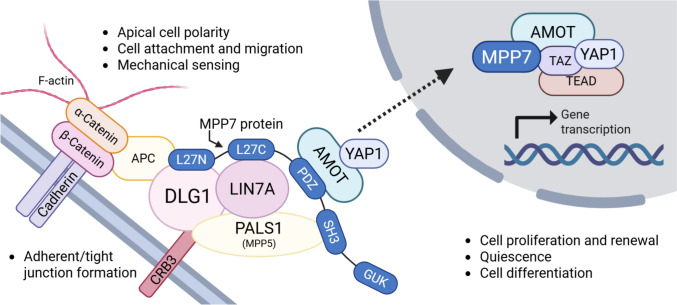 Figure 7
Figure 7- —http://dx.doi.org/10.13039/501100004329Javna Agencija za Raziskovalno Dejavnost RS
- —COST action
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBone health and osteoporosis research · Bone Metabolism and Diseases · Bone and Dental Protein Studies
Lay Summary
Osteoporosis causes bones to become fragile and more likely to break. We studied the gene MPP7, which has been linked to bone mineral density in genetic studies, but whose role in bone biology is unknown. We found that MPP7 levels are lower in bone from osteoporotic patients. When we deleted this gene in bone-forming cells, they were no longer able to initiate mineralization. The cells also changed shape, suggesting that MPP7 helps control their structure and function. These findings show that MPP7 is important for the mineralization process and may be a potential target for treating osteoporosis.
Introduction
Osteoporosis (OP) is the most common skeletal disorder in humans, characterized by decreased bone mineral density (BMD), reduced bone strength, and an increased risk of fractures. It affects more than one-third of postmenopausal women and, to a lesser extent, older men [1]. OP is asymptomatic in itself and is typically diagnosed only after a low-energy fracture, which is often followed by immobility or reduced physical activity, increased morbidity and mortality, and significant changes in patients’ lifestyles [2–4]. As such, OP substantially reduces quality of life, may lead to loss of independence, and poses a major healthcare and economic burden. With the aging global population, there is an urgent need for improved prevention strategies, early diagnostics, and novel treatment options.
Low BMD—the diagnostic marker of OP [5], is highly heritable and polygenic in nature, with heritability estimates ranging from 0.6 to 0.85 [6–8]. To date, genome-wide association studies (GWAS) and their meta-analyses have identified numerous loci associated with BMD [9–15]. However, the roles of many candidate genes within these loci in bone physiology remain largely uncharacterized. One such gene, located on chromosome 10, is membrane palmitoylated protein 7 (MPP7; also known as MAGUK p55 subfamily member 7), which has been associated with heel [14, 16–18] and lumbar spine [10, 19] BMD measurements. GWAS have identified the MPP7 locus (10p11.23) as being significantly associated with estimated bone mineral density (eBMD) (rs3905706) [10]. In addition, site specificity has been reported: MPP7 is associated with lumbar spine but not femoral neck BMD, consistent with trabecular versus cortical bone differences between these sites [15]. Beyond bone phenotypes, MPP7 has been robustly associated with lean mass in a large GWAS (n = 683,692, p = 1.5 × 10^−8^), pointing to a broader musculoskeletal role that further motivated its investigation in the context of bone–muscle cross-talk [19, 20]. Skeletal functional evidence is also available, as knockdown of MPP7 in zebrafish resulted in skeletal abnormalities, including impaired vertebral mineralization, which were partially rescued by human MPP7 mRNA [21]. Furthermore, MPP7 is constitutively expressed in human bone-derived cells during osteogenesis localizing to the plasma membrane and cytoplasm in osteoblasts [21]. Taken together, these data provide strong support for prioritizing MPP7 at this locus, although contribution of nearby genes cannot be completely excluded. MPP7 is a recently identified member of the membrane-associated guanylate kinase (MAGUK) family [22], primarily known for its role in membrane polarization and cell attachment. It interacts with LIN7A/C and DLG1 to form the MPP7–DLG1–LIN7A/C complex [23, 24], which co-localizes with E-cadherin at cell–cell junctions and is essential for tight junction formation [24, 25]. Similarly, MPP7 interacts with CRB3 and PALS1 (MPP5) to form the DLG1–MPP7–CRB3–MPP5 complex, which also localizes to adherens and tight junctions [25]. Interestingly, both DLG1 [16–18] and LIN7C [10, 15, 17, 18, 26] have been associated with BMD in multiple GWAS, highlighting the importance of cell adhesion and polarity in the regulation of BMD. Beyond its structural role, MPP7 is involved in regulating cell proliferation and stem cell fate during tissue regeneration. By forming a complex with angiomotin (AMOT) and yes-associated protein 1 (YAP1) following cell detachment, MPP7 modulates the entry of satellite cells into the cell cycle. Silencing MPP7 in muscle satellite cells reduces proliferation and forces premature differentiation, thereby depleting the satellite cell pool [27, 28].
Adequate proliferation, stem cell renewal, and osteogenic differentiation are central to bone homeostasis and skeletal integrity. Osteoblasts, the bone-forming cells, differentiate from mesenchymal stem cells (MSCs), which also give rise to myoblasts, chondrocytes, and adipocytes [29]. MSC commitment is governed by tightly regulated signaling pathways, particularly the Wnt and Hippo pathways, along with lineage-specific transcription factors that orchestrate terminal differentiation and functional maturation, including mineralization. To date, only one prior functional study used morpholino-mediated knockdown of MPP7 in zebrafish embryos, resulting in > 70% transcript suppression. The knockdown animals exhibited a characteristic ventral bending of the tail, impaired swimming ability, and markedly reduced vertebral bone mass and mineralization compared with wild-type (WT) controls and a pronounced deficit in vertebral mineralization. Co-injection of human MPP7 mRNA partially rescued the phenotype, reinforcing the specificity of the observed skeletal defect. These findings constitute the first in vivo validation of MPP7’s role in bone development [21].
The aim of this study was to validate the involvement of MPP7 in bone biology and to explore the underlying mechanisms by which it may influence BMD. We investigated whether MPP7 could serve as a potential therapeutic target or biomarker in bone diseases. Here, we report that MPP7 expression is reduced in osteoporotic bone and in muscle tissue from both osteoporotic and osteoarthritic patients compared to healthy controls. Furthermore, using CRISPR/Cas9-mediated MPP7 knockout (KO) in the human osteosarcoma HOS cell line, we demonstrate that MPP7 is critical for mineralization via regulation of ALPL expression. Our findings suggest that MPP7 is involved in the regulation of bone mineralization and may contribute to the development of OP.
Materials and Methods
Bioinformatic Tools for MPP7 Evaluation
To evaluate the functional relevance of MPP7 at the 10p11.23 locus, we performed in silico analyses using publicly available GWAS summary statistics for BMD. Bayesian analysis (HuGE calculator) [30] was used to compute Bayes Factors for causality, while Gene Sifter (Knowledge Portal, Gene Sifter) was applied to assess tissue-specific expression enrichment. In addition, the Gene Effector Index (GEI) [31] was used to integrate GWAS, chromatin, and eQTL data and estimate the probability of MPP7 as the effector gene at this locus.
Bone and Muscle Tissue Collection
Human bone and muscle tissue samples were collected from Slovenian patients with osteoarthritis (OA) and OP, as previously described [32]. Bone tissue samples (intertrochanteric region of the proximal femur) and skeletal muscle samples (gluteus medius) were obtained from 84 individuals undergoing hip arthroplasty—37 with OA and 47 with OP—at the Department of Traumatology, General Hospital Celje, and the Orthopedic Hospital Valdoltra. BMD of the contralateral hip and lumbar spine was assessed using dual-energy X-ray absorptiometry (DEXA; Hologic QDR-1000, Waltham, MA, USA) either before or after surgery. OP was diagnosed clinically since all patients sustained hip fractures; we also measured BMD at the hip, femoral neck, and lumbar spine by DXA, according to the WHO definition (t-score ≤ − 2.5). OA was diagnosed clinically and radiographically. Bone samples were obtained from the proximal femur during hip replacement surgery (OP and OA groups) or post-mortem (controls). All patients were treatment-naïve, and exclusion criteria were identical across groups, including systemic/metabolic diseases or medications known to affect bone metabolism. OA patients were recruited as a disease-control group to distinguish OP-specific changes from those related to other skeletal pathology. Controls consisted of 10 donors (mean age 68.1 years), in whom skeletal or metabolic disorders were excluded. The demographic and clinical characteristics of all groups, including age, sex, BMI, and BMD parameters, are outlined in Table 1 [33]. Bone and muscle tissue samples were obtained post-mortem from age-matched donors during routine autopsies at the Institute of Forensic Medicine, Faculty of Medicine, University of Ljubljana. Micro-CT scans of bone samples were performed as described previously [32], and the results showed that cadaver bone samples have normal bone structure, confirming that these samples are not osteoporotic and thus suitable as control samples. The study was approved by the Republic of Slovenia National Medical Ethics Committee (Reference Numbers: 0120-523/2016-2, KME 45/10/16). All participants provided written informed consent in accordance with the Declaration of Helsinki. Table 1. The demographic and clinical characteristics of all study groupsOA (n** = 37)OP (n = 47)K (n = 10)p (OA–OP)Age (years)71.1 (49–87)76.3 (53–88)68.1 (56–87)0.2513Women24380Men13910BMI (kg/m^2^)29.3 (22.8–43.7)25.1 (19.3–33.9)25.3 (19.0–312) < 0.001Hip BMD (g/cm^2^)0.909 (0.541–1.314)0.696 (0.402–0.974)na < 0.001Hip t-score − 0.629 (− 3.600 to 1.900) − 2.261 (− 4.400 to 0.000)na < 0.001Femoral neck BMD (g/cm^2^)0.791 (0.474–1.179)0.396 (0.386–0.798)n.i** < 0.001Femoral neck t-score − 1.192 (− 4.160 to 1.800) − 2.681 (− 4.200 to − 0.500)na < 0.001Lumbar spine BMD (g/cm^2^)1.015 (0.583 to 1.401)0.837 (0.601 to 1.461)na < 0.001Lumbar spine t-score − 0.465 (− 4220 to 2.800) − 2.006 (− 4.100 to 3.800)na < 0.001
RNA Isolation from Bone and Muscle Tissue
Following sample collection, 200 mg of bone and 100 mg of gluteus medius muscle tissue were immediately snap frozen in liquid nitrogen. Frozen bone and muscle samples were pulverized in liquid nitrogen and homogenized in 1 mL of TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Following the addition of 400 µL chloroform, samples were vigorously mixed and centrifuged at 12,000×g for 15 min at 4 °C. The aqueous phase was collected, and total RNA was isolated using the peqGOLD Total RNA Kit (PeqLab, VWR, Germany) as per the manufacturer’s protocol. RNA was resuspended in RNase-free water. RNA concentration and purity were assessed using a NanoDrop spectrophotometer (Thermo Fisher Scientific, USA), and RNA integrity (RIN) was evaluated using the Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA, USA).
Cell Culturing
The human osteosarcoma cell lines HOS (ATCC® CRL-1543™) and MG-63 (ATCC® CRL-1427™) were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). Both cell lines were cultured in low-glucose Dulbecco’s modified Eagle medium (DMEM; Gibco, Thermo Fisher Scientific, USA) supplemented with 10% fetal bovine serum (FBS), 2 mM l-glutamine, and 1% antibiotic/antimycotic solution (Gibco). Cells were maintained at 37 °C in a humidified atmosphere containing 5% CO_2_.
Osteogenic Differentiation of HOS Cells
For osteogenic differentiation, HOS and MG-63 cells were seeded at 5 × 10^4^ cells/cm^2^. After 24 h, differentiation was initiated by replacing the medium with osteogenic induction medium: complete growth medium supplemented with 100 nM dexamethasone, 50 µg/mL 2-phospho-l-ascorbic acid trisodium salt, and 10 mM β-glycerophosphate disodium salt hydrate (all from Sigma-Aldrich, USA). Medium was refreshed every 2–3 days. Non-differentiated control cells were maintained in complete medium for the full duration of the 35-day experiment. Samples were collected at day 0 (prior to induction) and then every 7 days. All experiments were performed in three independent biological replicates.
Alizarin Red S staining
Mineral deposition was assessed by Alizarin Red S staining as described previously [26]. At each time point, cells were fixed in 4% formalin for 10 min at room temperature, washed with distilled water, and stained with 2% Alizarin Red S solution (pH 4.2; Sigma-Aldrich) for 30 min. After extensive washing, stained cultures were visualized under a light microscope (EVOS XL Core Imaging System, Thermo Fisher Scientific). For quantification, the stain was extracted using 10% (v/v) acetic acid, and absorbance was measured at 405 nm.
Generation of MPP7 Knockout HOS Cells
Stable MPP7 KO HOS and MG-63 cells were generated using CRISPR/Cas9 gene editing. A guide RNA targeting exon 4 of the MPP7 gene (5′-CCAGCTTTGTCAACGGGATCTGG-3′; Ensembl ENST00000398795, GRCh38.p2) was cloned into the PX459 vector [pSpCas9(BB)-2A-Puro, Addgene #48139; gift from Feng Zhang] [30] using the BbsI restriction site (New England Biolabs, MA, USA). The construct was verified by DNA sequencing (Eurofins Scientific, Luxembourg). Transfection was performed using PolyJet™ In Vitro DNA Transfection Reagent (SignaGen Laboratories, MD, USA), with a PolyJet:DNA ratio of 3:1. After 24 h, cells were selected in 0.5 µg/mL puromycin (Sigma-Aldrich) for 2 weeks. Clonal populations were obtained using cloning rings, and following sufficient expansion, MPP7 KO was confirmed by Western blotting (Figure S1) as described previously [34]. Briefly, WT and MPP7-KO cells were trypsinized, counted and an equal number of cells was lysed and resolved by SDS-PAGE. Primary antibodies against MPP7 (1:1000 dilution, overnight incubation at 4 °C; Proteintech, Cat# 12983-1-AP, Rosemont, IL, USA) were detected by secondary anti-rabbit antibodies (1:10,000 dilution, 3-h incubation at room temperature; Cell signaling, Cat #7074), glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as loading control (1:1000 dilution; rabbit polyclonal, Sigma-Aldrich, #G9545).
RNA Expression Analysis by RT-qPCR
Total RNA was isolated and cDNA was synthesized with the High-Capacity cDNA Reverse Transcription Kit with RNase Inhibitor (Applied Biosystems, CA, USA) following manufacturers’ instructions. Quantitative PCR (RT-qPCR) was performed using the LightCycler 480 (Roche Diagnostics, Switzerland) with 2.5 ng/µL cDNA and Hot FirePol EvaGreen qPCR Supermix (Solis BioDyne, Estonia) according to manufacturers’ instructions. Cycling conditions were as follows: 95 °C for 15 min (initial denaturation), followed by 40 cycles of 95 °C for 5 s, 60 °C for 20 s, and 72 °C for 20 s. Gene expression was determined for the following markers; ALPL (alkaline phosphatase, biomineralization associated), collagen type I alpha 1 chain (COL1A1), RUNX family transcription factor 2 (RUNX2), osteocalcin (OC; bone gamma-carboxyglutamate protein), peroxisome proliferator-activated receptor gamma (PPARG), sclerostin (SOST), myogenic differentiation 1 (MYOD), and MPP7. EF1A1 and YWHAZ were used as reference genes for HOS cells, TBP for MG-63 cells [35], and RPLP0 for patient samples. Primers (Table S1) were designed using NCBI Primer-BLAST and synthesized by Macrogen Europe B.V. (Amsterdam, The Netherlands). Primer efficiency was validated with a standard curve, and specificity was confirmed by melting curve analysis. Gene expression was quantified in duplicate, and relative expression levels were normalized using the ΔΔCt method to the geometric mean of reference genes where applicable.
Statistical Analyses
Statistical analyses were performed using Prism 7.0 (GraphPad Software, La Jolla, CA, USA). Data are presented as mean ± SEM. Statistical significance was determined using Student’s t test, one-way or two-way ANOVA, followed by Bonferroni or Tukey’s post hoc tests. Statistical significance is displayed as follows: ns—not significant (P > 0.05); *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001; ****P ≤ 0.0001. MPP7.
Results
In Silico Prioritization of MPP7 at the 10p11.23 Locus
To further evaluate the relevance of MPP7 among candidate genes at the 10p11.23 locus, we performed bioinformatic analyses. Bayesian fine-mapping using the HuGE calculator assigned a Bayes Factor of 45 to MPP7 for lumbar spine and heel BMD, supporting a high probability of causality at this locus. Consistently, MPP7 scored significantly for bone tissue expression in Gene Sifter analysis (P = 0.0037). Finally, prioritization using the GEI highlighted MPP7 as the most likely effector gene in this region, with a higher probability of causality than neighboring genes (Fig. 1).Fig. 1. Gene Effector Index prioritization at the 10p11.23 locus. The Gene Effector Index analysis supports MPP7 as the primary effector gene underlying the GWAS association with lumbar spine BMD, when compared to other screened genes. Expression of MPP7 is decreased in human osteoporotic bone tissue
MPP7 has been identified as a strong candidate gene in multiple GWAS, examining heel [14, 16–18] and lumbar spine [10, 19] BMD. To assess whether MPP7 expression is associated with bone pathology, we analyzed its expression in bone tissue samples from individuals with OP, OA, and control donors (healthy cadavers) using RT-qPCR. MPP7 expression was significantly reduced in OP bone tissue compared to both OA and control (CTL) bone, with a 2.30-fold and 2.28-fold decrease, respectively (P = 0.0007; Fig. 2A). Similarly, we observed a significant reduction in the expression of RUNX2, a master transcription factor essential for osteoblast differentiation, matrix production, and bone mineralization. RUNX2 expression was approximately twofold lower in OP samples compared to controls (P = 0.002), with a comparable decrease also observed in comparison with OA bone tissue (Fig. 2B), consistent with our previous findings [32]. These results confirm that both MPP7 and RUNX2 are reduced in osteoporotic bone and are most probably involved in the pathological processes.Fig. 2. Expression of A MPP7 and B RUNX2 is decreased in osteoporotic (OP) but not osteoarthritic (OA) human bone samples. Gene expression was quantified by RT-qPCR, using RPLP0 as the internal control. Data are presented as mean fold change ± SEM. Statistical significance was assessed by one-way ANOVA followed by Tukey’s post hoc test, *P < 0.05; **P < 0.01; ***P < 0.001
MPP7 Expression is Decreased in Osteoporotic Muscle Tissue
Given the close developmental and functional relationship between bone and muscle—mediated through a shared MSC origin and continuous biomechanical and biochemical cross-talk—we next investigated whether MPP7 gene expression is also altered in skeletal muscle tissue from OP and OA patients. Total RNA was isolated from gluteus medius muscle samples, and the expression of MPP7 and myogenic differentiation 1 (MYOD1) gene was evaluated by RT-qPCR. Interestingly, MPP7 expression was significantly decreased in both OP and OA muscle tissue compared to controls, with a 2.6-fold and 3.9-fold reduction, respectively (P = 0.004; Fig. 3A). In contrast, expression of MYOD1, a key transcription factor regulating myogenic differentiation, exhibited only a slight, non-significant decrease. This reduction was more pronounced in OP muscle tissue, but did not reach statistical significance (Fig. 3B). These findings indicate that while MPP7 expression is reduced in muscle tissue from both OP and OA patients, this downregulation does not appear to significantly affect the transcriptional status of muscle differentiation.Fig. 3. Expression of A MPP7 is significantly decreased in skeletal muscle samples from osteoporotic (OP) and osteoarthritic (OA) patients, whereas B MYOD1 shows only a slight, non-significant reduction compared to controls. Human gluteus medius muscle samples were obtained from patients with OP (n = 7), OA (n = 11), and healthy autopsy donors (CTL, n = 6). Gene expression was measured by RT-qPCR using RPLP0 as the internal control. Data are presented as mean fold change ± SEM. Statistical significance was assessed by one-way ANOVA with Tukey’s post hoc test,**P < 0.01
Deletion of MPP7 Abolishes Mineralization in Human Osteosarcoma Cells
Analysis of MPP7 expression on WT HOS cells showed that MPP7 mRNA increases during differentiation process up to day 14 and then slowly decreases to day 35, indicating that MPP7 is involved in osteogenic differentiation (Figure S2). To further confirm the role of MPP7 in osteogenic differentiation and mineralization, we generated a CRISPR/Cas9-mediated MPP7 KO in the human osteosarcoma HOS and MG-63 cell lines and assessed their osteogenic potential in vitro. WT and MPP7-KO cells were cultured in osteogenic differentiation medium for 35 days. Differentiation was evaluated by measuring the expression of osteogenic markers, while mineralization was assessed both microscopically and spectrophotometrically using Alizarin Red S staining. No mineral deposition was observed in MPP7-KO cells throughout the 35-day differentiation period, as demonstrated by the absence of Alizarin Red S staining (Fig. 4A, C for HOS cells and Figure S3, for MG-63 cells). In contrast, WT cells of both cell lines showed detectable mineralization by day 21, with the mineralized matrix progressively expanding to cover the entire cell layer by day 35 (Fig. 4A). These findings indicate that MPP7 is essential for mineralization in HOS and MG-63 cells. To further characterize the mechanism underlying the mineralization defect, we quantified the expression of ALPL, an early osteoblast differentiation marker. In MPP7-KO cells, ALPL mRNA expression was completely abolished and did not increase at any time point during osteogenic induction (Fig. 4B for HOS and Figure S3B for MG-63). In contrast, HOS WT cells exhibited a gradual increase in ALPL expression starting at day 7, reaching a maximum by day 35 (Fig. 4B), while in MG-63, ALPL expression peaked at day 7 and gradually decreased till day 35 (Figure S3A). Consistent with its mRNA expression, ALP activity in MG-63 gradually increases till day 21 and then gradually decreases. These results suggest that loss of MPP7 disrupts the osteogenic mineralization signaling cascade upstream of ALPL expression, ultimately leading to a complete block in mineralization.Fig. 4. Mineralization and ALPL expression are abolished in MPP7-KO cells. Wild-type (WT) and MPP7-KO HOS cells were cultured in osteogenic differentiation medium and analyzed every 7 days up to day 35. A Representative light microscopy images of differentiating WT and MPP7-KO cells stained with Alizarin Red S to detect calcium deposits (red). Scale bars represent 1 mm. B Relative mRNA expression of alkaline phosphatase (ALPL) as determined by RT-qPCR. C Quantification of Alizarin Red S staining by spectrophotometry. Data represent mean ± SD of three independent biological replicates (N = 3), *P ≤ 0.05; **P ≤ 0.01
Deletion of MPP7 Affects Expression of Osteogenic Differentiation Markers in Osteosarcoma Cells
To identify which osteogenic and functional differentiation markers are affected by MPP7 deletion, we measured the mRNA expression levels of RUNX2, COL1A1, OC, and the adipogenic marker PPARG during the differentiation process. RUNX2 expression significantly increased in HOS WT cells during the early phase of osteogenic differentiation, peaking at day 7 and gradually returning to baseline by week 3 (Fig. 5A). In contrast, RUNX2 expression in HOS MPP7-KO cells was markedly reduced—2.2-fold lower than in HOS WT cells at day 7—and remained consistently lower throughout the first 21 days of differentiation, increasing only modestly over time. By week 4, RUNX2 levels were comparable between KO and WT cells. Interestingly, in MG-63 cell line, although RUNX2 expression increased with time of differentiation in both WT and KO cells, it was significantly higher in MPP-KO cell compared to WT MG-63 cells at all observed time points (Figure S4A). The expression of COL1A1, an early osteogenic marker, followed a similar trend in both cell lines for both WT and KO cells. In HOS cells expression increased until day 14, then gradually declined (Fig. 5B), while in MG-63 cells the highest expression was detected already at day 7, followed by gradual decline, expression in KO cells being slightly higher at all time points (Figure S4B). In MPP7, there were no statistically significant differences between KO and WT groups at any time point. Expression of OC, a marker of mature osteoblasts, was also induced during osteogenic differentiation but remained slightly lower in MPP7-KO HOS cells compared to WT (Fig. 5C), indicating impaired terminal differentiation, while no differences were detected in MG-63 cells (Figure S4C). In MPP7-KO cells, SOST mRNA expression, which is an osteocyte marker, was completely abolished and did not increase at any time point during osteogenic differentiation (Fig. 5E). In contrast, HOS WT cells exhibited a gradual increase in SOST expression starting at day 14, reaching a maximum by day 35. These results are in line with the previously demonstrated requirement of MPP7 for mineralization as shown by Alizarin Red staining and ALPL expression. They suggest that osteocyte formation is impaired, resulting in the absence of SOST expression. Interestingly, PPARG, the master regulator of adipogenic differentiation, showed a significant increase in HOS MPP7-KO cells during osteogenic induction (Fig. 5D), while its expression steadily decreased in WT cells, as expected. These findings suggest that loss of MPP7 alters the expression dynamics of key transcription factors involved in lineage commitment. Reduced RUNX2 and elevated PPARG expression in MPP7-deficient cells may indicate a shift away from the osteogenic toward adipogenic potential.Fig. 5MPP7 knockout affects the expression of osteogenic and adipogenic differentiation markers. Expression of A RUNX2, B COL1A1, C OC (osteocalcin), D PPARG, and SOST (E) was assessed in wild-type (WT) and MPP7 knockout (MPP7-KO) HOS human osteosarcoma cells during 35 days of osteogenic differentiation. Cells were collected weekly (ND not determined), and mRNA levels were quantified by RT-qPCR. Expression data were normalized to the geometric mean of EF1A1 and YWHAZ reference genes. Data represent mean ± SEM from three independent biological replicates (N = 3), *P ≤ 0.05; **P ≤ 0.01
Knockout of MPP7 in HOS Cells Results in Loss of Cell-Layer Morphology
Microscopic analysis of cultures stained with Alizarin Red S revealed distinct changes in cell-layer morphology during osteogenic differentiation in WT compared to MPP7-KO HOS cells, while no changes to cell-layer morphology were observed in MG-63 cells. In HOS WT cells, once confluence was reached, the monolayer began to organize into circular structures composed of margin cells and morphologically distinct central cells. These circular formations progressively increased in number and size over time (Fig. 6A; see also Fig. 4A). By day 21, mineral deposits began to appear in the center of these structures (Fig. 6B), suggesting that they may serve as nucleation sites for mineralization. As mineralization progressed, the deposits expanded to cover the entire cell layer, and the circular structures were no longer discernible. Interestingly, similar circular arrangements were also observed in WT cultures maintained in standard growth medium (i.e., without osteogenic supplements) for the full 35-day period. However, no mineralization occurred in these non-differentiated cultures (Fig. 6C), indicating that the formation of circular structures is a part of normal monolayer development upon confluence but does not lead to mineralization in the absence of osteogenic stimuli. Strikingly, these circular structures were entirely absent in MPP7-KO cultures throughout the differentiation period. MPP7-deficient cells retained a uniform morphology, and the monolayer remained morphologically homogenous (Fig. 6D; see also Fig. 4A). These observations suggest that MPP7 contributes to the spatial organization and structural remodeling of the osteoblast monolayer. The inability of MPP7-KO cells to form morphologically distinct domains may underlie their failure to initiate mineralization, pointing to a potential role for MPP7 in orchestrating osteoblast polarity and microenvironmental cues required for matrix deposition.Fig. 6MPP7 knockout disrupts cell-layer morphology during osteogenic differentiation. Wild-type (WT) and MPP7-KO HOS cells were cultured under osteogenic conditions, and morphological changes in the developing cell layer were assessed. Representative images show: A WT cells at day 14 of differentiation, B WT cells at day 21, C non-differentiated WT cells at day 21, and D MPP7-KO cells at day 21 of differentiation. Arrowheads indicate the margins of circular structures forming within the cell layer, and asterisks denote mineral deposits detected by Alizarin Red S staining. Scale bars represent 1 mm
MPP7 (MAGUK P55 scaffold protein 7) has emerged as a strong candidate gene for OP in several GWAS [14, 16]. To confirm its direct involvement in bone metabolism, we examined the role of MPP7 in bone mineralization and OP by analyzing its expression in human bone and muscle tissues and by generating a CRISPR/Cas9-mediated MPP7 KO in a human osteosarcoma cell model of osteogenesis [36]. Our results demonstrate that MPP7 expression is significantly reduced in osteoporotic bone tissue (Fig. 2) and that MPP7 plays a critical role in osteogenic differentiation and mineralization by modulating ALPL and RUNX2 expression (Figs. 4, 5).
Bone remodeling involves a dynamic balance between bone formation and resorption, and MPP7 may contribute to both processes. Xiao et al. reported increased MPP7 expression during osteogenesis of human bone-derived cells [21], which was also observed in WT HOS cell line (Figure S2), suggesting a potential role in osteoblast development. To test this directly, we knocked out MPP7 in HOS and MG-63 cells and found that mineralization was completely abolished in KO cells (Fig. 4), confirming a pivotal role for MPP7 in the mineralization process (Fig. 4). Mechanistically, MPP7 may contribute to mineralization by regulating intercellular junctions, Wnt signaling, or both. Previous studies have shown that cell–cell adhesion and junctional integrity are essential for osteoblast differentiation [37, 38]. For instance, inhibition of cadherin-mediated interactions suppresses BMP-2-induced ALPL activity and impairs matrix mineralization [39]. MPP7 has been shown to be essential for tight junction formation [24] and is a component of protein complexes involved in maintaining cell polarity and junctional organization [24, 25, 27] (Fig. 7). Specifically, MPP7 is part of the Crumbs complex, which plays a crucial role in epithelial polarity [25]. Thus, MPP7 may influence osteoblast differentiation via structural roles in cell–cell contact formation and polarity establishment [25, 40]. Supporting this hypothesis, MPP7-KO HOS cells exhibited altered monolayer morphology and failed to form the circular structures seen in WT HOS cultures—structures that appeared to serve as initiation sites for mineral deposition (Fig. 6). Interestingly, these structures also formed in non-differentiated WT cells cultured in growth medium, suggesting that they arise during confluency and may be required, though not sufficient, for mineralization. No such changes or any cell-layer organization was observed in neither WT nor MPP7-KO MG-63 cell line (Figure S3A).Fig. 7A schematic representation of presumed model of protein interactions of MPP7. MPP family proteins are adaptor molecules that assemble protein complexes consisting of protein families such as Crb homologues (e.g., CRB1, CRB3), DLT homologues, and LIN7 homologues (e.g., LIN7A, LIN7B, or LIN7C), together forming a cell adhesion signaling apparatus involved in establishment and maintenance of apical polarity at sites of adherent or tight junctions. MPP7 protein consists of several domains (i.e., L27N, L27C, PDZ, SH3, and GUK domains) that serve as docking sites for other proteins. While the DLG1–LIN7A–MPP7 protein complex is mainly involved in stabilization of the protein junctions, it also serves as a docking site for AMOT and YAP1 proteins, forming the MPP7–AMOT–YAP1 complex. Based on cell activity and signals form the extracellular environment, this complex can detach and serve as a shuttling complex for YAP1 nuclear entry, where it is also involved in formation of the YAP1/TAZ–TEAD transcriptional regulator complex, which regulates cell growth, proliferation, stem cell renewal, and differentiation. MPP7 protein is thus involved in establishment and stabilization of cell apical polarity through adherent and tight junctions, as well as in processes of cell proliferation, quiescence, and cell differentiation. DLG1 disks large homolog 1, LIN7 Lin-7 homolog, AMOT angiomotin, YAP1 yes-associated protein 1, TAZ (also WWTR1)—Tafazzin family protein, TEAD TEA domain transcription factor 1, APC adenomatous polyposis coli, CRB3 crumbs cell polarity homolog-3, PALS1 (also MPP5) protein associated with LIN7 1
Muscle tissue also plays a crucial role in skeletal health, not only by providing mechanical support but also by sharing the common MSC and cross-communication through both physical and biochemical signals [41]. Our results show that MPP7 expression is decreased in muscle tissue of OP and OA patients, but it does correlate with MYOD1,* the transcription factor* required for muscle cell differentiation. In the study by Andersen et al., polymorphisms in several muscle-related genes were shown to influence bone quality, supporting the concept of genetic interplay between muscle and bone. However, polymorphisms in MYOD1 did not demonstrate any significant association with BMD or fracture risk, suggesting that genetic variation in this myogenic regulator does not directly impact skeletal outcomes [42]. To further investigate the molecular pathways affected by MPP7 deletion, we analyzed the expression of key osteogenic markers including COL1A1, ALPL, OC, and RUNX2 and SOST (Figs. 4, 5). Strikingly, ALPL expression was absent in both HOS and MG-63 MPP7-KO cells (Fig. 4B), suggesting that MPP7 acts upstream of ALPL. As a critical enzyme for bone mineralization, ALPL contributes both to matrix deposition and signaling regulation. Its absence could directly explain the failure of mineralization in MPP7-KO cells.
Supporting this, Liu et al. [43] demonstrated that ALPL KO in murine osteoblasts downregulates Wnt signaling—a pathway essential for MSC commitment to the osteoblast lineage. Suppressed Wnt activity can lead to a shift from osteogenesis to adipogenesis [44]. In our HOS model, increased expression of PPARG, a master regulator of adipogenesis, was observed in MPP7-KO cells (Fig. 4D), suggesting a potential lineage shift. However, since Wnt signaling may not be the only affected pathway, additional mechanisms likely contribute. One such pathway may be Hippo signaling, as MPP7 interacts with AMOT and YAP1 in satellite stem cells to regulate proliferation and regeneration [27, 28] (Fig. 6). A limitation of our study is that HOS and MG-63 cells, although widely used for osteogenesis studies, are already committed osteoblast-like cells of fibroblast/epithelial origin. Thus, this model does not allow direct assessment of lineage commitment. Still, some degree of adipogenic gene expression has been reported in WT HOS cells [45], supporting the relevance of our observations.
Despite the fact that RUNX2 transcription factor regulates expression of COL1A1 and ALPL, there was no direct correlation between the expressions of these factors. RUNX2 transcription factor is involved during all stages of osteoblast differentiation, matrix production, and mineralization. Decreased RUNX2 expression results in abnormal bone development, characterized especially by lower osteoblast differentiation and consequent lower bone ossification [46]. While RUNX2 was moderately reduced, COL1A1 expression remained unaffected, and ALPL was completely absent (Figs. 3B, 4). This suggests that additional regulatory mechanisms influence the expression of these genes independently of RUNX2.
Conclusion
In conclusion, this study strongly suggests an association between the *MPP7 *gene expression and OP. Importantly, we provide evidence that deletion of MPP7 hinders mineralization of osteoblasts indicating that MPP7 has a direct role in bone formation mechanisms. Moreover, we show that MPP7 is a candidate biomarker for diagnosis of OP. These findings offer new insight into the complex mechanisms of bone pathophysiology and warrant further studies to elucidate the precise molecular pathways by which MPP7 regulates bone mineralization.
Potential Limitations of the Study
This study has several limitations. One major limitation is the lack of data on BMD on control (age- and sex-matched healthy cadavers) obtained through DEXA. The quality of the bone was instead confirmed through micro-CT, which showed that control samples have normal bone structure. Where necessary, BMD data from AO patients were used as a phenotypic and mechanistic “control” for OP patients in terms of bone density [47], as studies have shown that OA is often associated with normal or elevated BMD, particularly in subchondral regions [48, 49]. For other analyses, control (cadaver) samples were used.
Another limitation of the study is the low number of collected samples especially in control group, which combined with high heterogeneity of patient samples resulted in low statistical power. The role and effects of MPP7 should thus be confirmed and further explored on a bigger sample size.
Supplementary Information
Below is the link to the electronic supplementary material.Supplementary file1 (DOCX 551 KB)