Molecular Evaluation of Expression Changes in Genes Associated With Colistin Resistance and Virulence Development in Salmonella enterica From Two Iran Hospitals
Mohammad Darvishi, Shahriar Sepahvand, Hassan Sepahvand, Mohammad Ali Davarpanah, Mahboobeh Madani, Hesam Kamyab, Seyed Sobhan Behrouz, Farzaneh Asmani, Simin Yazdanpanah Ravari

TL;DR
This study examines colistin-resistant Salmonella strains in Iran hospitals, finding increased gene expression linked to resistance and virulence.
Contribution
The study evaluates gene expression patterns in colistin-resistant Salmonella isolates from Iran hospitals.
Findings
16% of Salmonella isolates showed colistin resistance.
CST-resistant isolates had increased pmrA and fliC gene expression.
Multidrug-resistant and colistin-resistant strains are widespread in hospitals.
Abstract
Salmonella entericais one of the most frequent causes of gastroenteritis in humans. The emergence of antimicrobial-resistant strains of the bacterium, especially those which are resistant toward colistin (CST), the latest antibiotic introduced to cure this bacterial disease, has become a severe health problem. Beside pmrA and pmrB genes, CST resistance is determined by mobilized colistin resistance (mcr) genes. Accordingly, in the present study, the expression pattern of these genes and fliC (encoding the flagellin protein) and agfA (encoding the bacterial fimbriae) was evaluated. To this end, a total of 50 S. enterica isolates were collected from two hospitals in Shiraz, Iran, during 2019-2020. The pattern of antimicrobial resistance of the isolates was determined by the disk diffusion method. Then, after the antibiotic sensitivity test, following RNA extraction, the expression of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
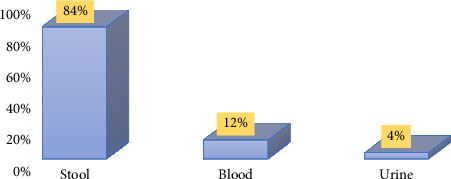 Figure 1
Figure 1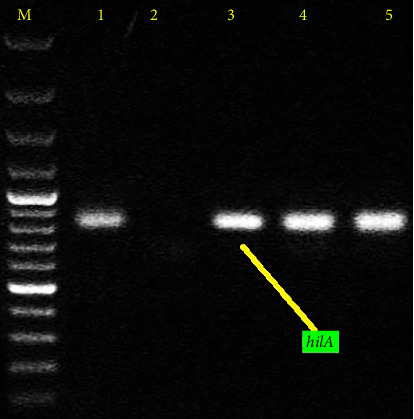 Figure 2
Figure 2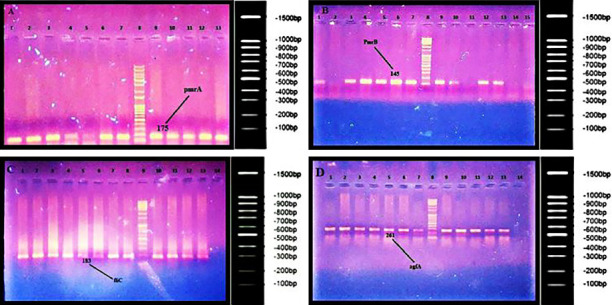 Figure 3
Figure 3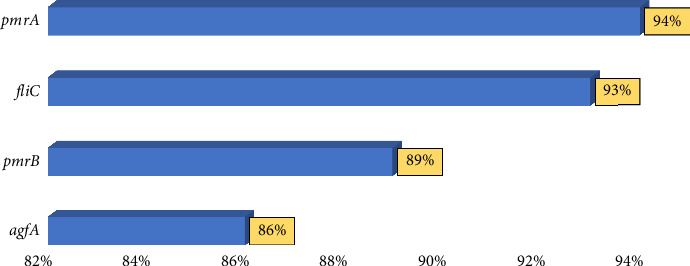 Figure 4
Figure 4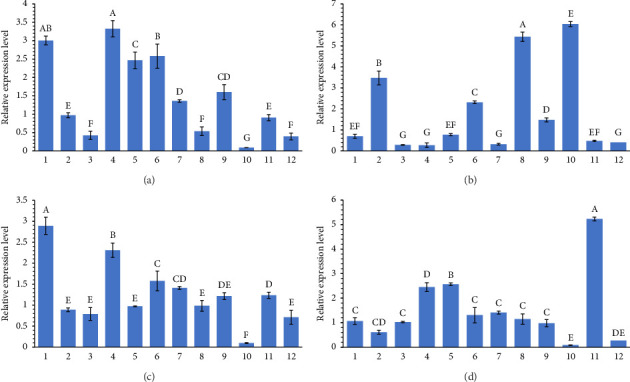 Figure 5
Figure 5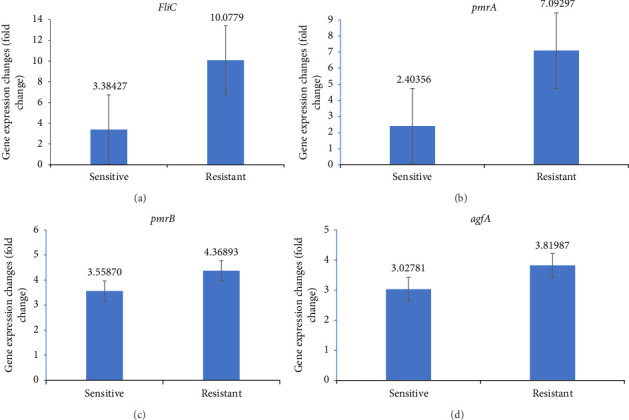 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · Antibiotic Resistance in Bacteria · Vibrio bacteria research studies
1. Introduction
The genus Salmonella belongs to the Enterobacteriaceae family [1]. The bacteria of this genus are Gram-negative bacilli, most of which are motile. Bacteria belonging to genus Salmonella could be both aerobic and anaerobic with optimal growth temperature of 37°C [2]. There are over 2650 serotypes of Salmonella, some of which are important enteric pathogens [3]. Salmonella species can infect humans and animals like chickens, turkeys, cows, and pigs. These species cause gastrointestinal disorders and are considered among the most important foodborne pathogens [4] and the main causative agents of the etiology of severe systemic diseases such as typhoid and paratyphoid fevers [5].
Virulence factors in Salmonella are divided into two major categories based on their genetic control: chromosomal and plasmid virulence factors. The importance of flagellum is that it allows movement, chemotaxis, and the establishment of bacteria in tissues. Flagellin also stimulates the immune system via the toll-like receptor 5 (TLR5) [6–8]. FliC, together with FljB, represents the structural components of the Salmonella flagellum highly expressed on the cell surface and regulated by transcription repressor FljA [9–12]. Fimbriae play a critical role in virulence by allowing bacteria to interact with host cells and other solid substrates [13]. Aggressive factor A (agfA) gene is also involved as a gene contributing to the production of fimbriae in the bacterial colonization in the intestine and systemic infection in the host [14].
The emergence of multidrug-resistant (MDR) bacterial strains has become a significant challenge for the health community [15] that can affect patients, economics, and the health system. It has considerable implications on public health. MDR is associated with prolonged length of stay, increased cost, and mortality [16]. Various bacterial pathogens have gained the ability to resist different antimicrobials. MDR Salmonella strains have evolved since the 1980s, and many salmonellosis outbreaks have been reported worldwide, especially from developing countries [17–19]. This issue becomes worse when some strains become resistant to new generations of antimicrobials or old efficient antibiotics. Colistin (CST) is an old antibiotic (over 50 years old) that is efficient against MDR Gram-negative bacteria, including Salmonella [20]. It should be noted that it is used as a last-line therapy against infections caused by these bacteria and is considered an antibiotic of critical importance in human clinical settings by the World Health Organization (WHO) [21]. However, due to the excessive consumption of this antibiotic by humans and the widespread use of this antibiotic in the veterinary medicine and livestock/animal farming [22], CST resistance is increasing among Gram-negative bacteria, including Pseudomonas aeruginosa, Acinetobacter baumannii, Escherichia coli, and S. enterica [21, 23].
As an antibiotic, CST targets lipid A moiety of the lipopolysaccharide (LPS), which is a surface antigen located in the outer membrane (OM) of Gram-negative bacteria [24]. LPS plays an essential role in the manipulation of OM permeability [25]. Through electrostatic interactions, positively charged moieties of CST (diaminobutyric acid residues) bind to lipid A. These interactions destabilize LPS and make the OM more porous. As a result, transportation via OM is disrupted, which will, in turn, lead to cell death.
By reducing the negative charge of LPS, mainly by the addition of positively-charged molecules such as phosphoethanolamine (PEtn) or 4-amino-4-deoxy-L-arabinose (L-Ara4N), the interaction between LPS and CST will be reduced; hence, the cell permeability will not be disrupted, and the bacteria will survive [26]. CST resistance in Salmonella is mainly chromosomal; however, plasmids might play a role in this phenomenon. In chromosomal resistance, two different regulatory systems, PmrA/PmrB and PhoP/PhoQ, make a significant contribution in this regard. PmrA/PmrB is involved in the synthesis of L-Ara4N, while PhoP/PhoQ is responsible for the biosynthesis of PEtn [27]. Specific mutations in the Salmonella genome led to the permanent expression of the components of these two systems. The addition of positively charged molecules, L-Ara4N and PEtn, to lipid A makes the CST–lipid A interaction less favorable. In addition, other strategies destabilize the CST–lipid A interaction, such as PagL-mediated lipid A deacylation [28]. The mobilized colistin resistance (mcr)-1 gene was mostly detected in Salmonella, but mcr-2, mcr-3, mcr-4, mcr-5, and mcr-9 were also detected. Since it has been reported for other bacteria, the direct relationship between the presence of mcr genes and CST resistance in Salmonella has not been conclusively established. These genes have also been found in sensitive strains [29].
In the present study, the presence and expression of two resistance related genes, i.e., pmrA and pmrB, and two virulence genes encoding fimbriae and flagellin proteins, agfA and fliC genes, respectively, were evaluated in patients with salmonellosis.
2. Materials and Methods
2.1. Informed Consent
All human participants consciously participated in this study, and sampling was performed with the patients' complete satisfaction. The study samples were collected from two hospitals in Shiraz, Iran: Ali Asghar Hospital and Ghadir Mother and Child Subspecialized Hospital during 2019 and 2020.
2.2. Isolation and Identification of Bacterial Strains
This cross-sectional descriptive study was conducted on patients suspected of infection with Salmonella enterica. According to the standard method, clinical specimens, including stool, blood, body fluids, etc., were taken from the suspected cases of salmonellosis infection. After sampling, the stool samples were immediately transferred to the selenite F medium for the selective enrichment of Salmonella species. The samples were incubated at 24°C for 6 hours. Then, the bacteria were transferred to selective culture media, such as Salmonella–Shigella (SS) agar, sulfite agar Bismuth, and other media and incubated at 37°C for 24 h. Blood samples were first cultured in the biphasic (solid and liquid) medium and then transferred to the selective culture media. The following day, Salmonella-suspected colonies were isolated and identified using biochemical tests. According to the manufacturer's instructions, colonies were further confirmed using a Microgen microbial identification kit (Microgen, UK).
2.3. Antibiotic Susceptibility Testing
Antibiogram test was performed using the disc agar-diffusion method [30] (in order to investigate the susceptibility of the isolates to different antibiotics (MAST UK Company), including tetracycline (TET), nalidixic acid (NAL), cefazolin (CFZ), ampicillin (AMP), chloramphenicol (CHL), and CST. Salmonella enterica subsp. enterica serovar Enteritidis ATCC 13076 strain was used as the positive control. The inhibition (nongrowing) zones were measured using a ruler and compared with existing Clinical and Laboratory Standards Institute (CLSI) tables for different antibiotics.
2.4. DNA Extraction
For DNA extraction, each isolate was grown in Luria Bertani broth (LBH). After 16 h of incubation at 37°C, DNA extraction and purification were performed using a commercial kit (CinnaGen, Iran) according to the manufacturer's instructions.
2.5. Tracking the hilA Gene via Polymerase Chain Reaction (PCR)
A pair of specific primers was used to identify the hilA gene. The forward primer sequence was 5′-CGG AAG CTT ATT TGC GCC ATG CTG AGG TAG-3′ and the reverse primer sequence was 5′-GCA TGG ATC CCC GCC GGC GAG ATT GTG-3′.
2.6. Amplification of the Selected Genes
PCR was used to amplify pmrA, pmrB, fliC, and agfA genes. The quality of the designed/applied primers in this study (Table 1) was checked on the NCBI web (https://www.ncbi.nlm.nih.gov/tools/primer-blast/). The primers' sequences and source are illustrated in Table 1. PCR amplification was performed in a total reaction volume of 25 μL containing 2.5 μL of 10 × buffer (Fermentas, Canada), 250 μM of each primer, 0.2 mM of each dNTP, 2.5-mM MgCl2, 0.75 U of Taq DNA polymerase ((Fermentas, Canada), 2 ng of purified DNA (as the template), and variable distilled water. PCR cycle condition was as follows: 94°C for 2 min; 35 cycles of 93°C for 1 min (denaturation step); 50°C–60°C for 1 min (annealing step); 72°C for 1 min (extension step); and 72°C for 10 min. After performing PCR, 5 μL of each reaction product was run on a 1% agarose gel, and the results were recorded using BioDoc Analyzer (Biometra, Germany).
2.7. RNA Extraction
RNA extraction was performed using bacterial RNA Isolation Kit Trizol Max (Invitrogen Life Technologies) following the provided protocol. Bacterial cells were harvested, resuspended in 200-μL accelerating factor, and incubated at 95°C for 4 min. After that, the cells were lysed in 1-mL Trizol. The lysed cells were incubated in the Trizol for 5 min at room temperature. A total of 100 μL of chloroform was added to the mixture and strongly shaken for 15 s. The mixture was incubated at room temperature for 2-3 min. The phases were separated by centrifugation at 12, 000 × g for 15 min, and 90% of the liquid phase (transparent color) was transferred to the Eppendorf tubes containing 500-μL isopropanol to precipitate the RNA. RNA was precipitated by mixing and incubating the tubes at room temperature for 10 min and then centrifugation at 12, 000 × g for 10 min. The precipitants were washed once with ethanol 70% and centrifuged at 7500 × g for 5 min. The precipitated RNA was solved in the nuclease-free water, and the concentration was determined using a spectrometer ND-1000 (NanoDrop Technologies, USA).
2.8. Real-Time PCR (RT-PCR)
RT-PCR was performed to investigate the expression level of the selected genes in CST-susceptible and -resistant isolates. RT-PCR was done using a One-Step Kappa Biosystem Kit (USA). The materials used in the final volume of 25 μL include 5 μL of RNA with a concentration of 5 ng, 0.5 μL of forward primer, and 0.5 μL of reverse primer with a concentration of 0.15 μM and 12.5 μL of master mix containing KAPA SYBR FAST, which was performed in a Corbett machine in Australia. The temperature program used in RT-PCR included 95°C for 5 min, 95°C for 30 s, 60°C for 30 s, and a final hold at 30°C for 40 cycles. Finally, the relative expression of pmrA, pmrB, fliC, and agfA genes was calculated by the ΔΔCт method. The 16S rRNA was used as the housekeeping gene. Used primers for each gene are presented in Table 1.
2.9. Statistical Analysis
The SPSS-21 software was used for data analysis. The normalization of differential expression of target genes was defined by REST software. One-way ANOVA was used to analyze the statistical differences among samples.
3. Results
3.1. Bacterial Isolates
In this study, 50 S. enterica isolates were isolated from Ali Asghar Hospital and Ghadir Mother and Child Hospital. Most isolates (84%) were from stool samples; a low percentage of isolates were from blood and urine samples (12% and 4%, respectively) (Figure 1).
3.2. Antibiogram Test
Chi-square test was used to compare the level of sensitivity between six antibiotics. Based on the results of this test, there was a significant difference in the level of sensitivity between six antibiotics (p < 0.001). The results of follow-up comparisons revealed that there was no significant difference in sensitivity between the two antibiotics NAL and CFZ (p < 0.05) and the sensitivity in these two was significantly lower than the antibiotics CHL, AMP, TET, and CST (p > 0.05). There was no significant difference in the sensitivity of the three antibiotics CHL, AMP, and TET (p < 0.05), and the sensitivity of these three antibiotics was significantly lower than the sensitivity to CST (p < 0.05) (Table 2). As CST is the last-resort treatment option for MDR S. enterica since there are few reports of CST-resistant Salmonella species in Iran and other countries, the resistance of these bacteria to CST is considered as a failure in an efficient treatment program. In examining the minimum inhibitory concentration for the CST antibiotic, 15% of the isolates were resistant (≥ 4 μg/mL), whereas 85% of the strains were susceptible (≤ 2 μg/mL).
3.3. The Results of hilA Gene Investigation
The amplification of the hilA gene via PCR revealed an 854-bp band in all samples tested, confirming the presence of Salmonella bacteria (Figure 2).
3.4. The Results of PCR for Investigating the Presence of pmrA, pmrB, fliC, and agfA Genes
The presence of the selected genes (pmrA, pmrB, fliC, and agfA) was investigated using the PCR method. The PCR products were then analyzed using gel electrophoresis (Figures 3(A), 3(B), 3(C), and 3(D)). The 94%, 89%, 93%, and 86% of the isolates carried pmrA, pmrB, fliC, and agfA genes, respectively (Figure 4).
3.5. Expression Analysis of the Selected Genes Using RT-PCR
The expression level of pmrA, pmrB, fliC, and agfA is presented in Figures 5(a), 5(b), 5(c), and 5(d). The fold-chain expression of all selected genes is presented in Figure 6. As can be seen, the fold-chain expression of all genes was significantly higher in CST-resistant isolates than those in the CST-susceptible ones (p < 0.01).
4. Discussion
S. enterica is the most important cause of salmonellosis in humans, and the prevalence of this infection has increased in recent decades [5]. The emergence of the MDR strains of this bacterium, especially CST-resistant ones, has become a significant health problem. The present study investigated the presence of the CST-resistant S. enterica isolates in clinical samples of two hospitals in Shiraz, Iran. Furthermore, the expression level of two CST-resistance associated genes, pmrA and pmrB, and two virulence genes, fliC and agfA, in hospital-acquired S. enterica samples were studied using RT-PCR.
In 2000, the WHO issued a significant warning about antibiotics referred to as “magic bullets” to end the misfortunes that infections brought to humans [15]. Antibiotic resistance in the community is present among the bacteria that cause primary infections. The community-acquired antibiotic resistance is a serious problem (e.g., resistance to penicillin and macrolide in Streptococcus pneumonia and some MDR strains of Staphylococcus aureus). The situation of antibiotic resistance in hospitals is more critical than that in the community, which affects the provision of health system services. S. enterica are among the bacteria that can acquire resistance to antimicrobial agents in many ways and transfer that trait to the following generations and even to other intestinal bacteria. Accordingly, they can spread and cause many epidemics. Excessive consumption of antibiotics in humans, veterinary, agriculture, and animal food products resulted in the emergence of antibiotic-resistant strains of S. enterica, a major epidemiological problem throughout the world. In MDR strains, CST is the last option for the treatment of the infection [21]. It means that the resistance to this antibiotic is a failure in the treatment of Salmonella-related diseases. This study indicates that CST resistance is considerable among isolated strains, attributed to improper and excessive consumption of antibiotics [31].
CST exerts its bactericidal activity by binding to bacterial cell membrane and disrupting its permeability, which results in the leakage of intracellular components [26]. Various mechanisms have been proposed for the CST resistance of S. enterica. In this bacterium, CST chromosomal resistance includes the activity of two-component regulatory systems (TCRSs) of PmrA/PmrB and PhoP/PhoQ responsible for biosynthetic changes of L-Ara4N and PEtn [34]. Activation of these systems is associated with environmental stimuli, such as the low concentration of Mg2+ or specific mutations in the genes encoding TCRSs. These mutations lead to the practical expression of PmrA/PmrB and PhoP/PhoQ. Plasmid-mediated CST resistance is conferred by MCR genes encoding PEtn [35], which add PEtn to lipid A and, as in chromosomal resistance, and contribute to decreased CST–LPS interactions. Plasmid-mediated resistance is less common in S. enterica.
The expression results indicated that the pmrA gene expression was significantly higher in CST-resistant strains than that in the susceptible ones. It is well established that the level of the pmrA expression is associated with CST-resistant phenotype [36]. However, no significant changes were observed in pmrB in CST-resistant and -susceptible strains in the present study. This issue may be due to the high expression level of pmrB but not pmrA in susceptible strains. In other studies, where the expression levels of pmrA and pmrB in CST-resistant and CST-susceptible S. enterica have been investigated, these two genes had been upregulated. This finding (in this work) may be related to the low number of susceptible isolates.
This study also observed that the flagellin-coding gene, fliC, is associated with the CST-resistant phenotype as well. To the authors' best knowledge, it is the first study that investigated the association between the fliC expression level of fliC and CST-resistant phenotype in S. enterica. Bacteria have different molecular identification patterns, such as lipoprotein (peptidoglycan), flagellin, DNA, and RNA, which are identified by various types of toll-like receptors (TLRs) [37]. Salmonella flagellin gene fliC is detected by TLR5, which activates NFkB to produce inflammatory cytokines such as IL-8, IL-10, pro-IL-1, and pro-IL18. Finally, neutrophils are called to the infection site to eliminate the organism [38]. Accordingly, perhaps mutations in the fliC gene cause a change in the structure of bacterial flagellin such that the immune system cannot detect and eliminate the bacterium. This vital issue is associated with increased expression of this gene in CST-resistant strains compared with susceptible ones. This study investigated the association between the agfA gene and CST resistance; agfA gene is one of the genes encoding the presence of the fimbriae, which also have properties related to pathogenesis and autoaggregation. Still, no significant change was observed in the expression level of the agfA gene in resistant and susceptible S. enterica strains.
5. Conclusions
The findings of this study show the relatively high rate of multidrug- and CST-resistant strains of S. enterica in patients, and it is an actual warning for the health community.
The results also showed that CST resistance in S. enterica is associated with increased expression of the pmrA and fliC genes. Still, it is not associated with pmrB and agfA genes. In the future, the suppression of these genes could contribute to their role in CST resistance and resistance elimination in the treatment process. Future studies can be performed on the suppression of these genes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Monte D. F. M. Sellera F. P. Salmonella Emerging Infectious Diseases 2020261210.3201/eid 2612.ET 2612 · doi ↗
- 2Banavandi S. Javad M. Shahhosseiny M. H. Selective Amplification of prt, tyv and inv A Genes by Multiplex PCR for Rapid Detection of Salmonella typhi Iranian Biomedical Journal 2005
- 3Kipper D. Hellfeldt R. M. De Carli S. Salmonella Serotype Assignment by Sequencing Analysis of Intergenic Regions of Ribosomal RNA Operons Poultry Science 201998115989599810.3382/ps/pez 2852-s 2.0-8507294544831134273 · doi ↗ · pubmed ↗
- 4Popa G. L. Popa M. I. Salmonella Spp. Infection-A Continuous Threat Worldwide Germs 2021111889610.18683/germs.2021.124433898345 PMC 8057844 · doi ↗ · pubmed ↗
- 5Rabsch W. Tschäpe H. Bäumler A. J. Non-typhoidal Salmonellosis: Emerging Problems Microbes and Infection 20013323724710.1016/s 1286-4579(01)01375-22-s 2.0-1714445527511358718 · doi ↗ · pubmed ↗
- 6Gulig P. A. Curtiss R. Plasmid-associated Virulence of Salmonella typhimurium Infection and Immunity 198755122891290110.1128/iai.55.12.2891-2901.19873316027 PMC 260003 · doi ↗ · pubmed ↗
- 7Josenhans C. Suerbaum S. The Role of Motility as a Virulence Factor in Bacteria International Journal of Medical Microbiology 2002291860561410.1078/1438-4221-001732-s 2.0-003622157212008914 · doi ↗ · pubmed ↗
- 8Milan C. Timm C. D. Fatores de Virulência Associados à Formação de Biofilme POR Salmonella enterica Science and Animal Health 201531 p. 94