Temporal propagation of neural state boundaries in naturalistic context
Djamari Oetringer, Sarah Henderson, Dora Gözükara, Linda Geerligs

TL;DR
This study explores how neural state boundaries spread across the brain during naturalistic movie watching, finding that they can propagate both top-down and bottom-up depending on the context.
Contribution
The first investigation of neural state boundaries in intracranial data during event segmentation, revealing context-dependent propagation patterns.
Findings
Neural state boundaries aligned with stimulus features and between brain areas.
Top-down propagation was observed at clause onsets and offsets.
Propagation direction varied depending on the stimulus input.
Abstract
Our senses receive a continuous stream of complex information, which we segment into discrete events. Previous research has related such events to neural states: temporally and regionally specific stable patterns of brain activity. The aim of this paper was to investigate whether there was evidence for top-down or bottom-up propagation of neural state boundaries. To do so, we used intracranial measurements with high temporal resolution while subjects were watching a movie. As this is the first study of neural states in intracranial data in the context of event segmentation, we also investigated whether known properties of neural states could be replicated. The neural state boundaries indeed aligned with stimulus features and between brain areas. Importantly, we found evidence for top-down propagation of neural state boundaries at the onsets and offsets of clauses. Interestingly, we did…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
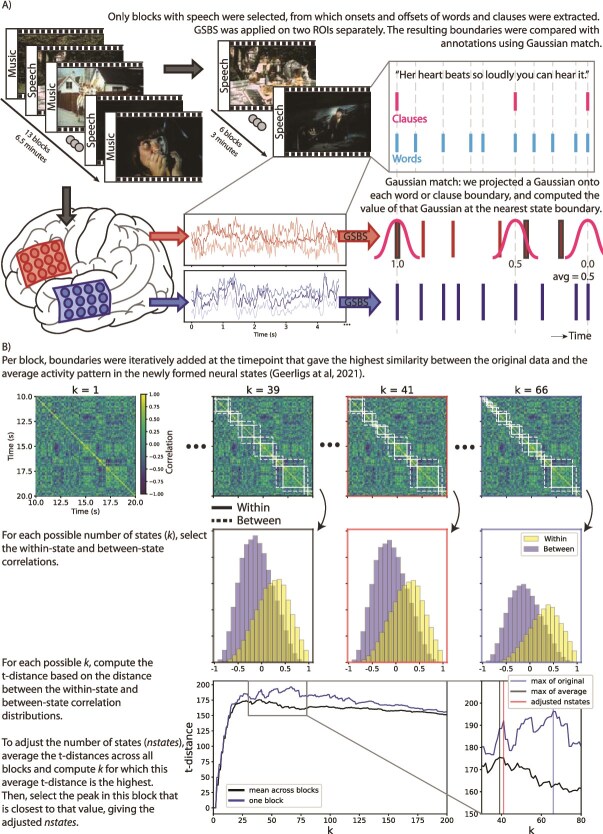 Fig. 1
Fig. 1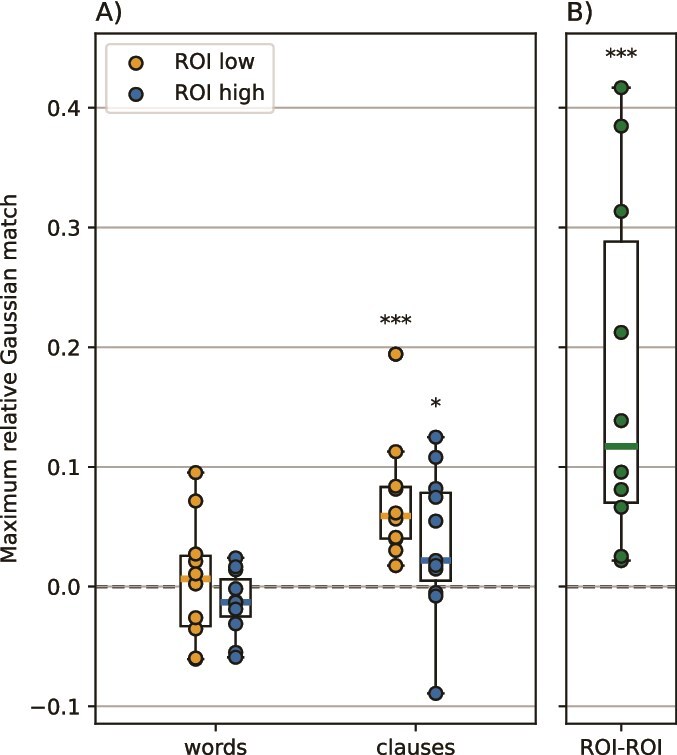 Fig. 2
Fig. 2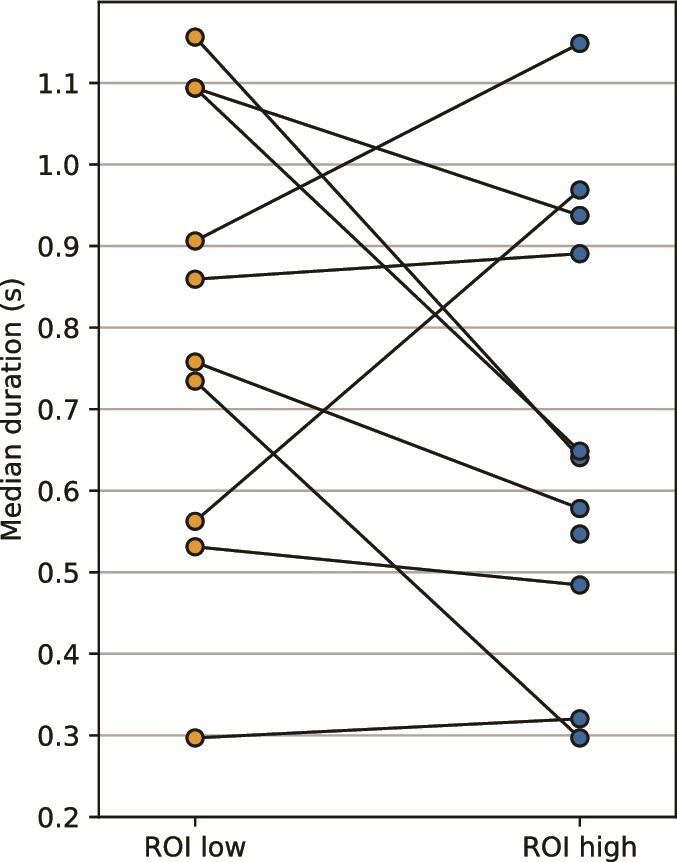 Fig. 3
Fig. 3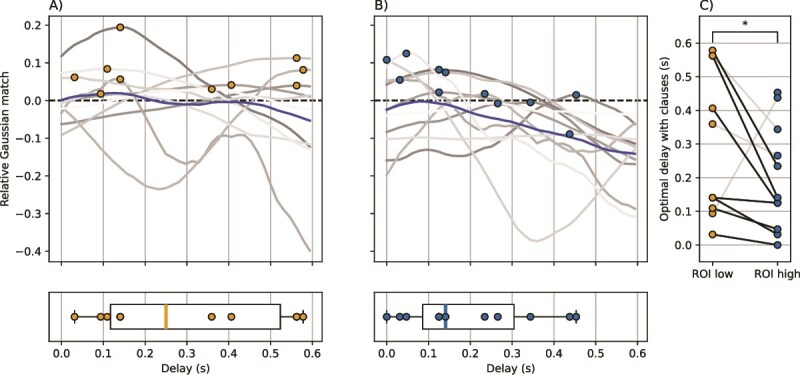 Fig. 4
Fig. 4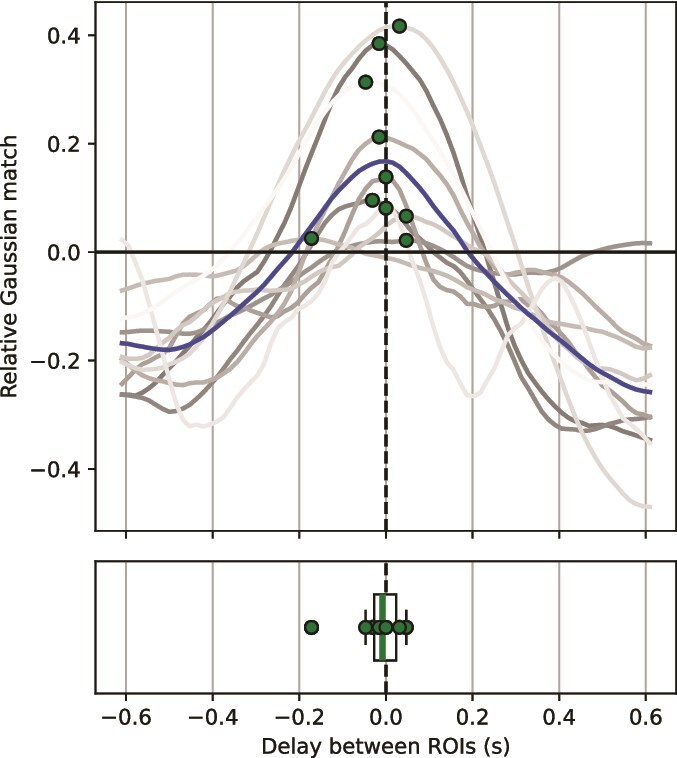 Fig. 5
Fig. 5- —Vidi
- —Netherlands Organization for Scientific Research
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeural dynamics and brain function · Neural Networks and Applications · EEG and Brain-Computer Interfaces
Introduction
The continuous influx of sensory input in our daily lives encompasses a vast amount of information. Despite its complexity, we can effortlessly process and utilize this information. This is due, in part, to our ability to automatically segment information into meaningful units (Zacks et al. 2007; Magliano et al. 2014; Zacks 2020), which supports information processing (Kurby and Zacks 2008), as well as memory encoding and retrieval (Swallow et al. 2009; Sargent 2013; Gold et al. 2017; Bird 2020; Shin and DuBrow 2020). Recently, various studies have investigated neural states as a possible neural underpinning of event segmentation. Neural states are temporarily stable patterns of brain activity in a local brain area, sometimes also termed “events in the brain” or “neural events,” which are observed across the cortex and are usually studied while subjects experience a naturalistic stimulus (Baldassano 2017; Geerligs 2022; Sava-Segal et al. 2023; Oetringer et al. 2025).
Although it has been shown that transitions in neural states (ie neural state boundaries) in different brain regions co-occur forming a nested hierarchy (Baldassano 2017; Geerligs 2022), it is not known how neural states are propagated across the cortex and what the role of bottom-up versus top-down signaling is. Looking at event segmentation, there is substantial evidence that both major low-level perceptual changes (Heusser et al. 2018) and changes in high-level features of a narrative, such as a change in the goals of a character (Zacks et al. 2009; Cutting 2014), can result in the perception of an event boundary. Similarly, both low-level and high-level features can lead to co-occurring neural state boundaries in feature-relevant areas (Oetringer et al. 2025). Given our previous observation of the nested hierarchy of boundaries, this suggests that boundaries could propagate across the cortex in a bottom-up but also in a top-down manner. For example, when there is a deviation from a social script that is detected in a higher-level brain area, such as a person introducing themselves with a high five rather than a handshake, this may result in a change in expectation for upcoming perceptual input, resulting in a neural state boundary in a lower-level area. Alternatively, a strong change in perceptual features could induce a boundary in a low-level area, which could then propagate to a high-level area when that perceptual change also results in changes in high-level representations of the scene.
Of particular interest when studying different levels of information and their interaction in the context of event segmentation is naturalistic language, which forms a nested hierarchy akin to events and neural states: multiple phonemes together form words, words form sentences, and sentences form narratives. Lexical processing is more apparent in lower-level auditory areas (eg anterior superior temporal gyrus), whereas the more complex syntactic integrations are more evident in higher-level areas (eg inferior frontal gyrus; Friederici 2002). Although less rich in sensory information, reading narratives also results in transient increases in activity around event boundaries in a network of brain regions that is similar to those found when watching audiovisual movies (Speer et al. 2007). Indeed, when reading a narrative, readers perceive temporal shifts as event boundaries (Speer and Zacks 2005). Large language models such as GPT-2 and GPT-3 have also been found to be able to segment text-only narratives similar to human event segmentation by providing instructions as one would do with human subjects (Michelmann et al. 2025), or by extracting measures of surprise and comparing those to the timings of human-annotated event boundaries (Kumar 2023).
One way to infer the presence of top-down and bottom-up propagation of neural state boundaries is by investigating timing differences between neural states in different brain regions and in relation to particular stimulus features. Studies so far have been unable to investigate these timings as they were mostly using functional magnetic resonance imaging (fMRI), which has very high spatial resolution but low temporal resolution. Instead, an electrophysiological method would be necessary to study the exact timings of neural state boundaries. Some recent studies have already explored the notion of neural states in electrophysiological data. In particular, Mariola et al. (2022) showed that neural states can be extracted from electroencephalogram (EEG) data, and that neural state boundaries align with event boundaries in the stimulus. Silva et al. (2019) related neural states in EEG to memory, by showing that neural state patterns during movie watching were reactivated during free recall of that movie. Finally, Henderson et al. (2025) further investigated the relationship between neural states and memory in different age groups, and found that greater distinction between neural states relates to better subsequent memory performance independent of age. Taken together, these studies show that neural states in electrophysiological methods show at least some similar characteristics as found in fMRI and, more importantly, could offer valuable insights into neural states that may not be possible in fMRI.
In this study, we intended to study the timing differences between neural state boundaries and the stimulus, and between the neural states of multiple brain areas. To this end, we used electrocorticography (ECoG), because, unlike EEG and MEG, it avoids issues of low spatial specificity by measuring local signals directly from different brain regions (Dubey and Ray 2019). As we were interested in natural language comprehension, we looked at neural states in a high-level and a low-level language area in an open intracranial electroencephalography (iEEG) dataset, which included ECoG data, and in which participants watched a movie. To first test whether neural states can be found and studied in ECoG data, we aimed to replicate neural state features observed in fMRI. We tested whether neural state boundaries in ECoG data also align with stimulus features, as well as align between different brain areas, and whether they show the expected temporal hierarchy of high-level areas exhibiting longer neural states than low-level areas. We hypothesized that these 3 strongly supported findings in fMRI are found in ECoG data as well, and that a top-down and/or bottom-up flow of information would be visible in the exact timing of the neural state boundaries. More specifically, Chang et al. (2022) reported a bottom-up between-network lag gradient in naturalistic language processing that was absent during rest, which is why we expected a bottom-up propagation of boundaries in general. However, although bottom-up processes could be dominant, others have reported the presence of top-down predictions during natural language processing in MEG (Heilbron et al. 2022). Thus, we also expected to see top-down propagation at moments in time that are important for high-level processing, such as the end of clauses. To preface our results, we indeed replicated the finding that neural states align with stimulus features and between brain areas. However, we did not find that neural states in higher-level areas are longer than those in lower-level areas, which was unexpected given the duration hierarchy observed in fMRI data. In addition, we found evidence for a top-down flow of information by looking at the exact timing of neural state boundaries in different areas in relation to the stimulus.
Materials and methods
Data acquisition
To investigate neural states with high temporal resolution and high spatial specificity, we used an open iEEG dataset (Berezutskaya 2022). In this dataset, 46 epilepsy patients were implanted with electrode grids (ECoG; exposed diameter of 2.3 mm, inter-electrode distance of 10mm) and watched a short audiovisual movie. The data were sampled at a rate of 512 Hz or higher. Some subjects (additionally) had depth electrodes and/or a high-density grid implanted, but these types of recordings were not used in the current study. More details about data acquisition can be found in Berezutskaya (2022).
Participants
According to the accompanying literature (Berezutskaya 2022), all participants were admitted to the University Medical Center Utrecht, Netherlands, for diagnostic procedures related to epilepsy. For some participants, the movie stimulus with ECoG measurements were part of clinical function mapping procedures, and these participants gave a written permission to use their clinical data for research purposes. Others participated as part of scientific research, in which case they gave their written informed consent to participate in research tasks. All participants gave their consent to share their de-identified data publicly. The study was approved by the Medical Ethical Committee of the University Medical Center Utrecht in accordance with the Declaration of Helsinki (2013). No additional ethical approval was required, as these are openly available data of which the ethical approval was established during data gathering by a previous study, and we did not gather any new data.
Only subjects with ECoG recordings were included. We excluded participants that were younger than 18 (20 subjects) to ensure that the developmental differences in younger brains did not affect the reliability of our results. Additionally, we excluded participants who had electrodes only in the non-language hemisphere (5 subjects), or had major issues in their brain anatomy such as tumors (1 subject), missing a substantial amount of brain tissue (2 subjects), or ventricles of an abnormal size indicative of severe atrophy (2 subjects). After preprocessing, an additional 5 subjects were excluded due to the presence of atypical activity based on visual inspection (see Supplementary Section A for visualizations and discussions). Of the remaining subjects ( \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} N\end{document} = 11, aged 19 to 52 yr, 4 male, 7 female), 6 saw the stimulus once, while another 5 had seen the stimulus before, with several days or weeks between viewings. Analyses were performed on all 11 subjects together.
Stimulus
Subjects watched an audiovisual movie of 6.5 min that was originally Swedish but dubbed into Dutch. They were free to move their eyes. The movie consisted of 7 music blocks, in which background music was presented and no character was talking, and 6 speech blocks in which the movie was presented as usual, including characters having conversations. These block types were interleaved, starting with music, and each block was 30 s. In this study, we only used the speech blocks in our analyses as our analyses focused on language. The dataset additionally included detailed speech annotations of various language features, of which we used clauses and words (see Fig. 1A, top). A “boundary timeline” for each of these features was created by having 1’s when there was an onset and/or offset of a clause/word, and 0’s at any other timepoint. For words, there were 540 1’s in total, of which 94 (= 17.4%) were offsets without the onset of a new word at the exact same time. For clauses, there were 153 1’s in total, of which 61 (= 39.9%) were offsets of a clause without the onset of a new clause.
A) Visualization of the experimental procedure, with subjects watching 1 uninterrupted movie consisting of music and speech blocks of 30 s each. B) Visualization of GSBS and computing the number of states. The time by time correlation matrix is actual data of 1 subject, cut to 10 s for visualization purposes. The visualized results are all based on a block of 30 s. Top left figure contains screenshots of the audiovisual stimulus, © The Astrid Lindgren Company. Used under fair use/fair dealing for scientific illustration.
Region of interest definitions
To be able to investigate the hierarchy of neural states within the language network, we defined 1 high-level language region of interest (ROI) and 1 low-level language ROI per subject. Specifically, we divided the language network as defined in Heilbron et al. (2022) into 2 parts. The high-level ROI consisted of Brodmann areas 38, 39, 40, 44, 45, 46, 47, which include Broca’s area and temporal pole. The low-level ROI consisted of Brodmann areas 20, 21, 22, 41, 42, which include the primary auditory cortex. The Brodmann area per electrode was determined by segmenting the subject-level anatomy into Brodmann areas using the PALS-B12 atlas (Van Essen 2005) in Freesurfer (version 7.3.2). See Supplementary Section B for the number of electrodes per ROI per subject. One subject did not have any electrodes in the low-level ROI, and was thus excluded from any analysis that required this ROI.
Preprocessing
To clean the data, we first applied a procedure based on Zada (2024) and Goldstein (2022) in all electrodes within a subject. We first dropped the bad electrodes as labeled by Berezutskaya (2022), who visually inspected the data with respect to artifacts and outliers and checked whether any electrodes were placed on top of others. Then the data were high-pass filtered at 0.1 Hz. Spikes in the data were automatically detected and interpolated. Then, data were re-referenced using either a common average filter or an independent component analysis (ICA)-based method, depending on the noise profile of the subject. In case of ICA-based re-referencing, we used the InfoMax algorithm and omitted bad spikes during fitting. Bad components were then selected based on visual inspection: first the mixing matrix was inspected to identify components that were shared among many electrodes, and then the timeseries of these components were inspected and identified as “bad” if it indeed looked non-neural. After re-referencing, a notch filter with a width of 2 Hz was applied at 50 Hz to remove line noise. Then the electrodes were selected per ROI as described above. Additional bad electrodes were identified based on visual inspection of the timecourses of that particular ROI and excluded from analysis. The data were then downsampled to 64 Hz, which was a balance between improving computational complexity and keeping a high enough temporal resolution for our analyses, then clipped at 3 times the standard deviation, and finally z-scored to have a mean of zero for each electrode.
Data-driven detection of neural state boundaries
To identify the exact locations of neural state boundaries, we used Greedy State Boundary Search (GSBS; Geerligs et al. 2021) on single-subject data, per speech block and per ROI (Fig. 1A, bottom). Given time by space data, GSBS defines neural states iteratively in a fully data-driven manner, visualized in Fig. 1B. In most research so far, GSBS is applied to group-averaged fMRI data, but ECoG data cannot be averaged across subjects due to the subject-specific electrode locations. Notably, previous literature has demonstrated that GSBS works for single-subject electrophysiological data as well (Mariola et al. 2022; Henderson et al. 2025). The parameters were configured according to the original recommendations by Geerligs et al. (2021): the maximum number of neural states was set to half the number of time points in each run, and fine-tuning was set to 1, allowing each neural state boundary to be adjusted by 1 timepoint after each iteration for a better fit if necessary. We specifically utilized the states-GSBS algorithm, which is an enhanced and more reliable version of GSBS introduced by Geerligs (2022), capable of potentially placing a full neural state (ie 2 boundaries) in a single iteration instead of always placing just 1 boundary at a time.
Within GSBS, the optimal number of states is determined using the t-distance metric, which represents how similar within-state activity patterns are and how dissimilar activity patterns between consecutive states are. The optimal number of states is then determined by taking the number of states that gives the highest t-distance. However, we have found that this approach results in the number of states being highly variable across blocks in this particular dataset, with for example 1 subject having 20 to 31 states per 30 s for 5 blocks, and having 152 states for the remaining 30-s block. This discrepancy may be because of the lower signal-to-noise ratio in single-subject ECoG data, as compared to group-averaged fMRI data for which GSBS was originally developed and tested. To achieve a more stable state segmentation across blocks within each participant, we adjusted our procedure for selecting the optimal number of states, visualized in Fig. 1B, bottom. First, we averaged the t-distance metric across blocks, resulting in an average optimal number of states per 30 s by taking the highest t-distance from this averaged distribution. Next, we fine-tuned the number of states per block, by finding a peak in the t-distance metric per block that is closest to the chosen optimal number of states across all blocks. Here, a peak is defined as any point, or a pair of points with the exact same value, with a lower value on both the left and the right side.
Gaussian match
To determine the extent of having a “match” between 2 boundary timelines, such as when comparing neural state boundaries in 1 ROI to the other, we developed a new metric called Gaussian match. Previously used match metrics applied to fMRI data such as counting the number of overlapping boundaries (Baldassano 2017; Geerligs 2022) or correlations (Oetringer et al. 2025) are not applicable here, since the much higher temporal resolution of ECoG data requires more flexibility in the relative timing of 2 matching boundaries. We therefore developed a measure that weighs the amount of match depending on the temporal distance between 2 boundaries, rather than having an all-or-nothing approach. We did this by projecting a Gaussian (SD = 332 ms, mean of 0), of which the amplitude has been scaled between 0 and 1, onto each boundary in 1 timeline. Then, to compute the match of the current boundary, the closest boundary in the other timeline was selected and the match was determined using the amplitude of the projected Gaussian at the timepoint of this closest boundary (Fig. 1A, bottom-right). Thus, the further away the closest boundary in the other timeline is located, the lower the match, with a maximum of 1 when the 2 boundaries occur at the exact same point in time. If the SD of the projected Gaussian is too big, there would be a high match for chance-level timings as well, thus it would be hard to detect anything above chance-level. An SD that is too small would lead to boundaries that are actually close enough to be called a (weighted) match, but are not actually seen as a match. The SD was thus chosen in an attempt reflect a good balance between too many false positives (missing boundaries that actually match) and too many false negatives (an increase in chance-level match). However, we did not test different SD values to avoid double dipping.
To determine the overall match between 2 timelines, 1 timeline is chosen as the seed. Then for each boundary in this seed timeline, the match is computed using the closest boundary in the other timeline. Then, the average match across all boundaries is computed as the Gaussian match. Thus, the Gaussian match metric is non-commutative, since choosing the other timeline as the seed gives a different Gaussian match value. Therefore, it is important to consider which timeline becomes the seed timeline. When investigating whether boundaries in stimulus features co-occur with neural state boundaries, we used the stimulus feature timeline as seed since having extra neural state boundaries should not have a negative impact on the match. When comparing neural state boundaries between different ROIs, we took the timeline with the lowest number of boundaries as the seed boundary, to prevent 1 area having more boundaries than the other from negatively impacting the match value.
Our method is similar to the one presented in Henderson et al. (2025) for investigating boundary overlap in EEG data. They convolved the neural state boundary timeline and subsequently correlated the result with the “event boundary density,” which was a non-binary measure for event boundaries, to alleviate the challenge of identifying the exact timing of event boundaries. In our case, we have precise timings of words and clauses and therefore chose not to convolve those with a Gaussian. Instead, we used a projection of a Gaussian to quantify the match between 2 binary and temporally precise timelines.
Accounting for delays
Because of the high temporal resolution of electrophysiological data, a delay may be detected between the stimulus and corresponding activity in a specific brain area, or between 2 brain areas. Such delays have been estimated before in ECoG (Flinker 2015), with an onset as early as 34 ms in the temporal cortex and 240 ms in Broca’s area, but the exact delay will depend on the specific electrode locations, which differ per subject. When computing the match between 2 timelines, we therefore computed the Gaussian match over various delays. When comparing stimulus features to neural features, we shifted the stimulus feature timeline forward in time ranging from a delay of 0 to 600 ms, with steps of 1/64 s ( \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} \approx 16\end{document} ms). When comparing 2 timelines of neural features, the possible delays instead ranged from roughly −600 to 600 ms, with steps of 1/64 s. The optimal delay was then defined as the delay that gave the maximum Gaussian match.
Relative Gaussian match
The Gaussian match in the data ( \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} match{_}data\end{document} ) will always be above zero, also when boundaries are not aligned with each other. The Gaussian match that can be expected by chance is dependent on the number of neural states and their durations in each timeline. In order to get a measure for the match relative to chance-level, we computed a null distribution of the maximum Gaussian match by shuffling the neural states in 1 timeline 1,000 times, and re-computing the maximum Gaussian match over all delays per permutation. The chance-level Gaussian match is then defined by taking the mean of all the maximum Gaussian matches across permutations (giving us \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} match{_}null\end{document} ). The relative Gaussian match was then defined as \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} (match{_}data - match{_}null)/(1-match{_}null)\end{document} , giving a value of around 0 when the Gaussian match in the data is at chance-level, and 1 when this match is perfect. Group-level statistics can then be computed over the maximum relative Gaussian match per subject using the Wilcoxon signed-rank test.
Results
Because this is the first study of neural state boundaries in ECoG data, we first set out to replicate the findings of previous fMRI studies. Specifically, we investigated whether boundaries align with stimulus features, whether they overlap across brain regions, and whether a duration hierarchy is present. Next, we investigated whether there was any evidence of top-down and bottom-up propagation of neural state boundaries. To do so, we investigated the timing differences across regions in their alignment to the stimulus and to each other.
Neural state boundaries align with speech
To assess whether neural states in ECoG data are related to the stimulus, we investigated the alignment between neural state boundaries and stimulus features, specifically the on- and offsets of words and clauses. In the low-level ROI, subjects had an average of 236.7 neural state boundaries across 3 min (standard deviation of 130.5) and 285.1 boundaries in the high-level ROI (standard deviation of 140.1), which are both much lower than the number of onsets and offsets of words (538), and higher than the number of onsets and offsets of clauses (152). See Supplementary Section C for example visualizations of the timelines. Per ROI and per stimulus feature, we computed the maximum Gaussian match relative to chance-level between the neural state boundaries in the ROI and the onsets and offsets of the stimulus feature. Using a one-tailed Wilcoxon signed-rank test on the maximum relative Gaussian matches per subject (N = 11 for high-level ROI, N = 10 for low-level ROI), we indeed found a significant match between neural states and the on- and offsets of clauses (Fig. 2A; low-level ROI: \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} P < 0.001\end{document} , mean = 0.072, SD = 0.049; high-level ROI: \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} P = 0.034\end{document} , mean = 0.036, SD = 0.045). For words, both the low-level ROI ( \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} P = 0.065\end{document} , mean = 0.005, SD = 0.049) and the high-level ROI ( \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} P = 0.139\end{document} , mean = −0.014, SD = 0.026) did not show such an alignment. One possibility is that having many more onsets and offsets of words than the number of neural state boundaries could explain this difference between words and clauses. We therefore performed a post hoc analysis in which we extracted the non-optimal GSBS-solution that had the same number of neural state boundaries as the number of onsets and offsets of words, but there was still no significant alignment (low-level ROI: \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} P = \end{document} 0.99; high-level ROI: \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} P = \end{document} 0.98; results not visualized). Together, these results indicate that neural states in ECoG data are indeed aligned with the stimulus, specifically with the on- and offset of clauses. The lack of alignment with words could be due to the electrode locations being too widespread in ECoG to capture such a relatively low-level language feature.
*Neural state boundaries in both low- and high-level ROI aligned with clauses but not with words. Additionally, neural state boundaries between the low-level and the high-level ROI significantly align with each other, with each subject having a match above chance. A) Maximum relative Gaussian match per subject for words and clauses, and in both ROIs. Within 1 box plot, each dot is 1 subject. B) Maximum relative Gaussian match per subject between the 2 ROIs. Each dot is 1 subject. A and B) *\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} \end{document}, **\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} \end{document}, **\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} \end{document}.
Neural boundaries between areas overlap
Based on previous studies with fMRI data, we expected to observe significant overlap between the boundaries in the different ROIs (Baldassano 2017; Geerligs 2022). To investigate this boundary alignment, we computed the maximum relative Gaussian match between the low-level and high-level ROI per subject (Fig. 2B). We found that all individual subjects had a maximum relative Gaussian match of above chance-level. Using a one-tailed Wilcoxon signed-rank test on these maximum relative Gaussian matches per subject ( \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} N=10\end{document} ), we found that the match between the neural state boundaries of the 2 ROIs was significantly above zero ( \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} P < 0.001\end{document} ). This indicates that boundaries indeed showed significant overlap between the 2 ROIs.
Duration hierarchy is absent
Previous literature has found a relatively consistent neural state duration hierarchy in fMRI data: low-level areas have shorter neural states than high-level areas (Baldassano 2017), forming a gradient across the cortex (Geerligs 2022). We accordingly expected our high-level language ROI to have longer neural states than the low-level language ROI, and we therefore computed the median neural state length per subject per ROI, taking all speech blocks together. However, we found very inconsistent results across subjects (N = 10): 6 subjects had longer neural states in the low-level ROI, and 4 subjects had longer neural states in the high-level ROI (Fig. 3). To investigate the possibility that this inconsistency could be due to taking many anatomically separated Brodmann areas together to define the high-level language ROI, we re-ran GSBS and the analysis on smaller subsets of high-level electrodes that were anatomically close to each other in Supplementary Section D. However, the inconsistency remained and we therefore conclude that the neural state duration hierarchy is not present. This inconsistency with previous literature could perhaps be due to the size and shape of the ROIs. Previous fMRI studies have used small clusters of voxels, while our ECoG electrodes within 1 ROI encompass a much larger area of 5 or 7 Brodmann areas. Extracting neural states from such a large area could affect the detected state durations in different ways. On the one hand it could diminish neural state boundaries in various smaller areas within the ROI, creating longer neural states overall. On the other hand it could merge multiple neural state boundaries from smaller areas together, creating shorter neural states overall. Alternatively, perhaps the hierarchy of neural state durations is only observable at the timescale of fMRI and not that of electrophysiology. Additionally, previous studies that demonstrated this duration hierarchy of neural states were based on group-averaged data, which makes the state boundaries fully stimulus driven (Baldassano 2017; Geerligs 2022). In contrast, for this dataset, boundaries could have also been driven by endogenous factors, such as mind wandering. This may result in a less robust duration hierarchy.
Median state duration in the low- and high-level ROIs. The durations of neural states are longer in the higher-level ROI than in the low-level ROI for 4 subjects, but longer in the low-level ROI for 6 subjects. Each line is 1 subject.
Optimal delay with speech is longer in high-level areas
To investigate the presence of top-down and bottom-up propagation of neural state boundaries, we investigated whether the optimal delay between the stimulus and the neural state boundaries would be different for the 2 ROIs. The low-level ROI having a shorter optimal delay with speech than the high-level ROI would imply that speech-related boundaries arrive in the low-level ROI before they arrive in the high-level ROI, indicating a bottom-up information flow. Alternatively, the high-level ROI having a shorter optimal delay with speech would be indicative of top-down processes, as speech-related boundaries would then occur in the high-level ROI before they occur in the low-level ROI. Although we had previously shown that the maximum relative Gaussian match between neural state boundaries and clauses was significantly above chance-level at the group level, the subject-level curves of match over delay, as well as the optimal delays, highly varied across subjects (Fig. 4A and B). To test whether the optimal delays differed between the low-level and the high-level ROI, we subtracted the optimal delay with clauses in the low-level ROI from the optimal delay in the high-level ROI, and applied a 2-tailed Wilcoxon signed-rank test. We only applied this analysis to the optimal delays with the onsets and offsets of clauses, and not of words, as we did not observe a significant alignment between neural state boundaries and word onsets and offsets. We additionally only included subjects with an above-chance relative Gaussian match between neural state boundaries and clauses in both ROIs ( \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} N = 7\end{document} ), as the optimal delay is meaningless in subjects with an alignment at or below chance-level. We indeed found that the optimal delay is significantly longer in the low-level ROI relative to the high-level ROI (Fig. 4C; \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} P = 0.016\end{document} ; mean difference = 123 ms, SD = 0.204 ms). Of the 7 included subjects, 5 had a surprisingly fast optimal delay in the high-level ROI of under 180 ms. Given that all included subjects have a shorter optimal delay with clauses in the high-level ROI than in the low-level ROI, we can conclude that neural state boundaries related to the start and/or end of clauses occur in the high-level ROI before occurring in the low-level ROI, and thus that the alignment with clauses capture a top-down flow of information.
*Although the alignment between the onset and offsets of clauses and neural state boundaries was significantly above chance-level (Fig. 2A), the optimal delay with those onsets and offsets varied across subjects. When comparing the optimal delays within each subject, the optimal delay with clauses was shorter in the high-level ROI than in the low-level ROI, implying that clause-related neural state boundaries occur in the high-level ROI before the low-level ROI. A) Relative Gaussian match with clauses over delay in the low-level ROI. Each gray line is 1 subject with a colored marker at the maximum, and the blue line is the average across subjects. B) Relative Gaussian match with clauses over delay in the high-level ROI. C) Optimal delay with clauses per subject. Each line is 1 subject. The gray lines are subjects with a Gaussian match at or below chance level in at least 1 ROI. The black lines are subjects with a Gaussian match above chance in both ROIs. \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{upgreek} \usepackage{mathrsfs} \setlength{\oddsidemargin}{-69pt} \begin{document} \end{document}.
Inconsistent delay of boundaries between low- and high-level areas
To investigate whether this top-down flow of information is present across all timepoints or whether it is instead specific to the on- and offsets of clauses, we investigated the delay between boundary timings across the ROIs, irrespective of the stimulus. We did not observe evidence for a consistent delay in the boundary timing across the ROIs (Fig. 5). Instead, most subjects had an optimal delay between the 2 ROIs of around zero with 3 subjects showing a bottom-up delay (low-level ROI before the high-level ROI), 5 subjects showing a top-down delay (boundaries in the high-level ROI before the low-level ROI), and 2 subjects having an optimal delay of exactly 0 s. Given that the alignment with clauses did show a consistent delay, these results suggest that some boundaries (eg those related to higher-level features of the stimulus) may be propagated in a top-down fashion, while other boundaries are propagated in a bottom-up fashion. The latter could include boundaries that are based on more low-level stimulus features, or boundaries that are driven by endogenous changes in brain activity that are not stimulus-driven (eg task-irrelevant thoughts or a moment of distraction). Another potential reason for the inter-subject variable delays between the 2 ROIs could be that different participants had variable electrode locations within each ROI. To sum up, these analyses show that, although the alignment with clauses imply a top-down flow of information, this direction of information flow is not observed across all boundaries.
Relative Gaussian match over delay between the low-level and high-level ROI. A negative delay implies that high-level boundaries were generally earlier than low-level boundaries. Each gray line is a subject, and the blue line is the average curve across subjects. The green dots are the maximum relative Gaussian matches per subject.
Familiarity did not shift neural state boundaries
Having seen the same movie stimulus before affects neural activity patterns. In the context of neural states, Lee et al. (2021) showed that neural state boundaries shift earlier in time the more often subjects had watched the stimulus. Since 5 of our subjects had seen the exact same stimulus before, this could impact our results. To ensure that any difference in boundary timing is not due to multiple exposures, we studied whether the familiarity impacted boundary timing. This analysis is discussed in Supplementary Section E, together with optimal delay results if we were to divide the subjects into 2 groups based on familiarity. In summary, we did not find the anticipation effect in these ECoG data. This could perhaps be due to the amount of time between viewings, which was several days up to several weeks. With such a long period, many details are likely to be forgotten, making the second viewing less predictable than when viewed on the same day or more than twice. Taking everything together, we conclude that the significant difference in optimal delay with speech between the low-level and high-level ROI (Figure S5B,C,E), as well as the inconsistent optimal delays between ROIs (Figure S5D), cannot be driven by a subset of subjects having watched the stimulus before.
Discussion
Neural states have previously been investigated as a possible neural underpinning of event cognition. It has been proposed that each area along the cortex segments incoming information into distinct neural states. This proposal is supported by various fMRI and EEG studies that have investigated state boundary alignment with the stimulus as well as the relationship between neural states and memory. Using open ECoG data, here we studied the exact timings of neural state boundaries in multiple brain areas to specifically investigate the presence of bottom-up and top-down propagation of boundaries. We found support for the presence of top-down propagation, extending our understanding of information segmentation throughout the cortex. We additionally replicated multiple attributes of neural states previously observed in fMRI, further supporting the potential of studying neural states through electrophysiological data.
High temporal resolution uncovers top-down processes
Utilizing the high temporal resolution of ECoG, we studied precise timing differences in neural state boundaries to investigate the presence of top-down and bottom-up processes. Indeed, we found that the optimal delay with the onsets and offsets of clauses is shorter in the high-level than in the low-level area, consistently across subjects. This indicates that clause-relevant neural state boundaries occur in high-level areas before low-level areas.
The exact timing differences we observed between stimulus features and neural states suggest that the higher-level brain areas may be able to predict the onset and offset of clauses. Previous research has shown that predictability makes neural state boundaries occur earlier in time, and boundaries in higher-level areas are affected more than those in lower-level areas (Lee et al. 2021). Many subjects had an optimal delay of under 180 ms in the high-level language area, while Goldstein (2023) found an optimal delay of 292 ms in the IFG. Additionally, Flinker (2015) reported that the first activation in Broca’s area (part of IFG) in a word-repetition task occurs at 240 ms post stimulus onset, indicating that the clause-related boundaries in our study occur before the IFG has actually received the bottom-up end/start-of-clause information. In the low-level language ROI however, all but 1 subject had an optimal delay that was longer than that found by previous literature of 55 ms (Goldstein 2023) and 34 ms (Flinker 2015), indicating that the clause-related neural state boundaries occur some time after receiving the relevant bottom-up information. These opposite results in the 2 ROIs are in line with the finding that, during speech comprehension, temporal regions are involved in bottom-up processing of speech, while frontal regions (including Broca’s area) are more involved in the top-down mechanisms (Zekveld et al. 2006).
Taking these numbers together, we conclude that the onsets and/or offsets of clauses are likely predicted, leading to neural state boundaries in high-level areas that are earlier in time than what can be expected from a pure bottom-up flow of sensory input. Indeed, language studies have often reported prediction effects during language comprehension, including the possibility of using information from high-level representations to facilitate the processing of incoming information at the level of syntactic structure (eg Arai and Keller 2013) as well as incoming low-level perceptual information (Dikker et al. (2009); see Kuperberg and Jaeger (2016) for a review). Other studies have argued that this prediction could be implemented through a top-down signal that leads to pre-activation (eg lexical pre-activation (Fruchter et al. 2015), pre-activation of syntactic gender in spoken language comprehension (Van Berkum et al. 2005), and semantic pre-activation (Altmann and Mirković 2009)). Such pre-activation in high-level language areas, perhaps initiated by higher-level multimodal areas during movie watching, could result in an earlier neural state boundary that relates to the onset or offset of a clause. Although we were only able to show such top-down processes at one level of the cortical hierarchy due to the constraints in this particular study, such processes could be present at other levels as well, creating a hierarchy of linguistic predictions, shown to be present in natural language comprehension using EEG and MEG (Heilbron et al. 2022) and in fMRI (Schmitt 2021; Caucheteux et al. 2023). This prediction hierarchy could go beyond language comprehension and instead encompass information processing in general in a multimodal naturalistic environment (Schmitt 2021), in line with the idea of event segmentation and a hierarchy of neural states across the cortex (Baldassano 2017). Predictability was however not controlled in this study, and we therefore cannot make any claims about it beyond speculation. Instead, future electrophysiological studies could study these propagation dynamics while controlling the predictability, for example by presenting the same stimulus multiple times (similar to Lee et al. 2021).
Noncontinuous top-down processing may indicate global updating of events
Although we did find support for top-down processes at the onsets and offsets of clauses, such processes may not be continuous. Namely, we did not find a consistent delay between the high- and low-level ROI when we looked at all neural state boundaries, irrespective of the stimulus. This suggests that the top-down direction of boundary propagation does not generalize to all boundaries. Together, our results imply that top-down processes happen at or around the onsets and offsets of clauses, rather than all the time. When we generalize this observation beyond our specific stimulus, these results suggest that boundaries can propagate in a top-down manner when there are relevant changes in context that could affect the processing of information in lower-level areas. This is in line with some studies finding bottom-up processes to be dominant (Chang et al. 2022), while others find top-down processes during natural language comprehension (Heilbron et al. 2022).
Given that the top-down processes seem to happen mostly at clauses and not across all neural state boundaries, the flow of information may depend on what exactly is happening in the stimulus. This is an extension of Baldassano (2017)’s proposal in which all brain areas across the cortex segment incoming information, and mostly included upward propagation of information. Our results suggest that a neural state boundary in a high-level area can trigger a top-down flow of information, strengthening information separation in lower-level areas.
In relation to the cognitive skill of event segmentation, previous studies have related event boundaries to group-level neural state boundaries in high-level areas (such as the lateral and medial prefrontal cortex) while participants were watching a movie (Baldassano 2017; Geerligs 2022; Oetringer et al. 2025), or listening to music (Williams 2022), as well as to individual-level neural state boundaries (Sava-Segal et al. 2023). Such event boundaries could then perhaps trigger a top-down updating process in lower-level areas, possibly leading to a change in representation of lower-level features, or a prediction of a change. This would be in line with global event updating: the representation of an event as a whole is updated, even when some features (such as characters or spatial locations) remain stable, akin to Event Segmentation Theory (Zacks et al. 2007). Others have proposed that events could instead be updated incrementally (Zwaan et al. 1995; Huff et al. 2014), which would be more congruent with a bottom-up propagation of information. Given that we found evidence for a top-down flow of information but only at particular points in the stimulus rather than continuously, the global and incremental updating perspectives could be combined, with incremental updating happening in general, and global updating at particular moments that fulfill certain criteria. This would correspond to behavioral findings indeed supporting both global and incremental updating (Kurby and Zacks 2012; Bailey and Zacks 2015).
The concept of neural states generalizes from fMRI to electrophysiology
In the context of event segmentation, neural states have previously mostly been studied in fMRI, and strong support has been found for the hypothesis that all brain areas across the brain segment incoming information. To investigate whether neural states can be studied in ECoG data as well, we first studied whether the neural state boundaries in 1 brain area were aligned with the neural state boundaries in another. We indeed found within-subject alignments between these areas, with all subjects showing a match above chance-level. This is in line with Baldassano (2017) and Geerligs (2022), who both showed that neural state boundaries are often shared across brain areas.
We additionally demonstrated that our 2 language areas both showed an alignment with speech in the external environment, and in particular with the onsets and offsets of clauses. This corresponds with the notion that each brain area along the cortex segments information in a way that is at least partially stimulus-driven. This has also been discussed and supported in our previous work (Oetringer et al. 2025), showing that location-sensitive areas tend to have a neural state boundary when there is a change in location in the movie stimulus. We did not find such an alignment with words in the language ROIs, which may be due to the overall size of these regions being too large to capture such low-level characteristics.
One phenomenon that is consistently observed in fMRI, but that we could not replicate with ECoG data, is the hierarchy of neural state durations (Baldassano 2017; Geerligs 2022; Sava-Segal et al. 2023). Such a hierarchy would be in line with the previously reported intrinsic temporal hierarchies (eg Hasson et al. 2008; Honey et al. 2012; Stephens et al. 2013; Raut et al. 2020). In such hierarchies, various areas across the cortex have been shown to be sensitive to different timescales, and in particular such that low-level sensory areas operate over the shortest timescales, while high-level multimodal regions operate on the longest timescales. Perhaps we were not able to find this neural state hierarchy because of how we defined our regions of interest, which were relatively large compared to fMRI studies due to the nature of ECoG data. Taking multiple areas together, which each have their own intrinsic neural timescale (see Wolff 2022,for a review), could make the duration hierarchy less robust. We also encountered some challenges with defining the optimal number of neural states, compared to previous fMRI studies. In particular, we found larger intra-subject variability than would be expected, potentially due to the signal-to-noise ratio being lower than in group-averaged fMRI data on which GSBS has been developed and tested (Geerligs et al. 2021). Group-averaging is unfortunately not possible in ECoG due to the subject-specific electrode locations. Although some adjustments were necessary in determining the optimal number of neural states (see Materials and methods), we were still able to extract neural states in a fully data-driven approach.
Even though our 2 ECoG ROIs were larger and much more sparsely sampled than voxels usually studied with fMRI, we were still able to detect neural states, similar to Henderson et al. (2025), Mariola et al. (2022), and Silva et al. (2019), who defined neural states in EEG data. This is not surprising, as there is a large body of literature around neural states at almost all spatiotemporal scales. If we consider neural states to be temporarily stable neural activity patterns that abruptly shift into new patterns regardless of their spatial scale, then neural states have actually been studied with a multitude of imaging methods at a wide range of spatial scales. These include local populations with multivariate single cell activity (Abeles 1995), the whole brain as different configurations of global functional connectivity networks (Yamashita et al. 2021; Song et al. 2023), as well as multivariate whole-brain voxel activity states with fMRI (Zhang et al. 2021), and as microstates with EEG (Michel and Koenig 2018) and MEG (Tait and Zhang 2022). In fact, evidence also suggests a direct correspondence between electrophysiological microstates and fMRI functional connectivity states (Britz et al. 2010; Michalopoulos and Bourbakis 2015). Such “neural states” have been associated with a lot of behavioral and cognitive processes, such as attention, memory, exploration, and more (Panagiotides et al. 2011; Yamashita et al. 2021; Suhail et al. 2022; Artoni 2023; Song et al. 2023). This background makes us confident that our sparsely sampled ECoG neural states are still methodologically sound and behaviorally meaningful.
Although studying neural states in ECoG comes with extra challenges, we were still able to replicate multiple fMRI findings, indicating that neural states are present and relevant in ECoG data as well despite the difference in temporal resolution. In particular, this is the first study to our knowledge that shows a neural state boundary alignment between multiple brain areas in electrophysiological data. Showing this alignment in ECoG is of particular importance, as it cannot be studied in MEG and EEG due to their lower spatial specificity. This study therefore provides a stronger foundation for studying neural states in existing and future electrophysiological studies, including more accessible methods such as EEG and MEG.
Conclusion
By investigating neural states in ECoG data while subjects were watching a movie, we were able to show that neural state boundaries can be propagated in a top-down manner. In particular, the top-down processes in general language areas take place at onsets and offsets of clauses. We did not find a similar effect for words rather than clauses, possibly because of the electrode placement and subsequent ROI definitions. We additionally showed that fMRI findings generalize to electrophysiology, giving stronger incentive to (further) investigate neural states in electrophysiological data, which includes ECoG, but also EEG and MEG.
Supplementary Material
Supplementaries_bhaf284
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abeles M et al. 1995. Cortical activity flips among quasi-stationary states. Proc Natl Acad Sci. 92:8616–8620. 10.1073/pnas.92.19.8616.7567985 PMC 41017 · doi ↗ · pubmed ↗
- 2Altmann GT, Mirković J. 2009. Incrementality and prediction in human sentence processing. Cogn Sci. 33:583–609. 10.1111/j.1551-6709.2009.01022.x.20396405 PMC 2854821 · doi ↗ · pubmed ↗
- 3Arai M, Keller F. 2013. The use of verb-specific information for prediction in sentence processing. Lang Cogn Process. 28:525–560. 10.1080/01690965.2012.658072. · doi ↗
- 4Artoni F et al. 2023. Microsynt: exploring the syntax of eeg microstates. Neuro Image. 277:120196. 10.1016/j.neuroimage.2023.120196.37286153 · doi ↗ · pubmed ↗
- 5Bailey HR, Zacks JM. 2015. Situation model updating in young and older adults: global versus incremental mechanisms. Psychol Aging. 30:232–244. 10.1037/a 0039081.25938248 PMC 4451381 · doi ↗ · pubmed ↗
- 6Baldassano C et al. 2017. Discovering event structure in continuous narrative perception and memory. Neuron. 95:709–721.e 5 8. 10.1016/j.neuron.2017.06.041.28772125 PMC 5558154 · doi ↗ · pubmed ↗
- 7Berezutskaya J et al. 2022, 2022 ISSN. Open multimodal i EEG-f MRI dataset from naturalistic stimulation with a short audiovisual film. Scientific Data. 9, 9:1–13. 10.1038/s 41597-022-01173-0. URL https://www.nature.com/articles/s 41597-022-01173-0.35314718 PMC 8938409 · doi ↗ · pubmed ↗
- 8Bird CM . 2020. How do we remember events? Curr Opin Behav Sci. 32:120–125 4. 10.1016/J.COBEHA.2020.01.020. · doi ↗