Assessing species-specific neonicotinoid toxicity using cross-species chimeric nicotinic acetylcholine receptors in a Drosophila model
Anna Lassota, James J. L. Hodge, Matthias Soller

TL;DR
Researchers used fruit flies to study how changes in a specific brain receptor affect sensitivity to insecticides, finding that swapping parts of the receptor reduced toxicity but impaired movement.
Contribution
A novel cross-species chimeric nicotinic acetylcholine receptor model was created to assess neonicotinoid toxicity differences between species.
Findings
Flies with the α8/β2 chimeric receptor showed impaired motor functions in climbing and flight assays.
Flies with the chimeric receptor or β2 knock-out had increased survival after neonicotinoid exposure compared to wild-type flies.
Combinatorial insecticide exposure did not reveal significant differences in survival.
Abstract
Nicotinic acetylcholine receptors (nAChRs) are ligand-gated ion channels and the main mediators of synaptic neurotransmission in the insect brain. In insects, nAChRs are pivotal for sensory processing, cognition and motor control, and are the primary target of neonicotinoid insecticides. Neonicotinoids are potent neurotoxins, and pollinators such as honey bees are more sensitive and affected by extremely low sub-lethal doses. nAChR subtypes exist as homomers of α-subunits or heteromers composed of α and β subunits. The honey bee nAChRα8 subunit is orthologous to nAChRβ2 in Drosophila, raising the question of whether this α to β change makes flies less sensitive to neonicotinoids. To investigate species-specific aspects of neonicotinoid toxicity, we CRISPR-Cas9 engineered a cross-species chimeric nAChR subunit by swapping the ligand-binding domain in Drosophila of nAChRβ2 with honey bee…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
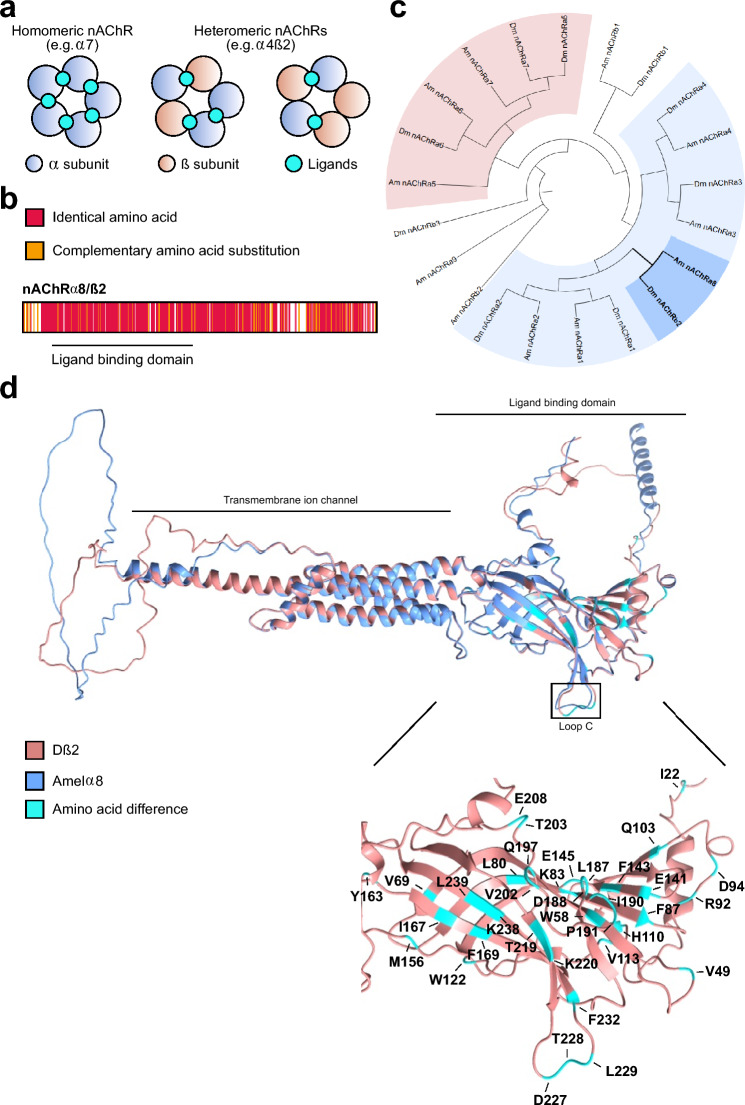 Figure 1
Figure 1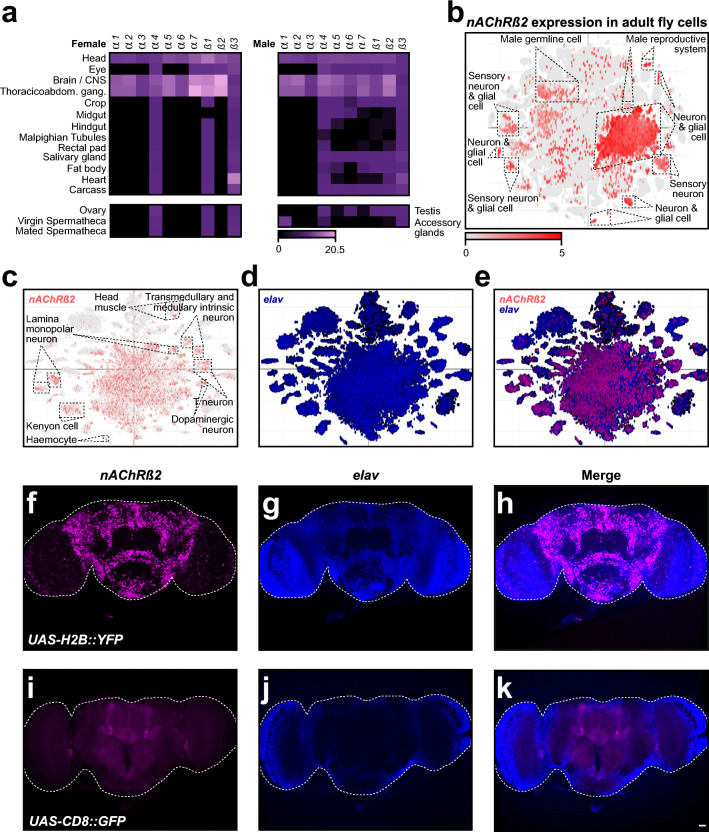 Figure 2
Figure 2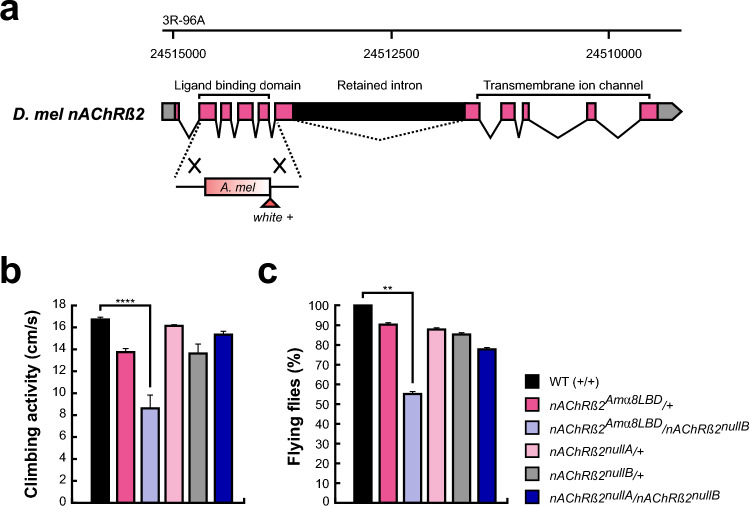 Figure 3
Figure 3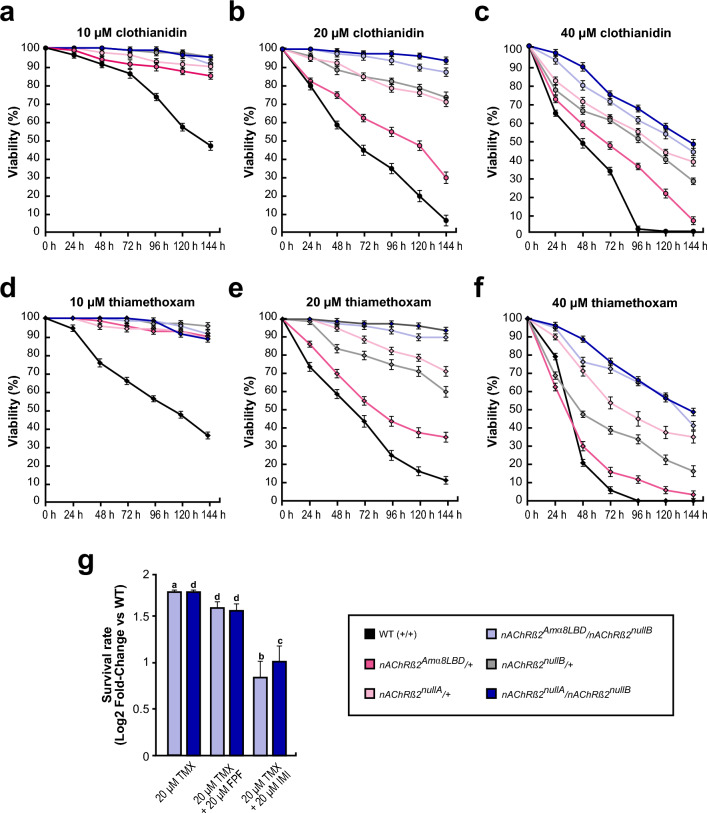 Figure 4
Figure 4- —https://doi.org/10.13039/501100000268Biotechnology and Biological Sciences Research Council
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect and Pesticide Research · Insect-Plant Interactions and Control · Plant and animal studies
Introduction
Pesticides are indispensable for protecting global agricultural yields by reducing crop losses caused by pests^1^. Among these, neonicotinoids are highly effective due to their broad-spectrum efficacy and systemic activity, which enable integration into plant tissues for targeted pest control, and versatility in application methods, such as seed coating, to reduce environmental contamination^2,3^. Neonicotinoids exhibit high toxicity to insects due to their selective action on insect nAChRs. While originally developed as safer alternatives for vertebrates, accumulating evidence has raised concerns about their effects on mammals, including humans^4–8^. Furthermore, neonicotinoids do not distinguish between pests and other insects, and harm beneficial pollinators like honey bees, which are critical for both wild flora and agricultural crops^9–11^. The widespread decline of pollinators, partially attributed to pesticide exposure, poses significant ecological and economic challenges, highlighting the need for pest-control strategies with reduced non-target effects.
Honey bees, wild bees and other pollinators are adversely affected by sub-lethal doses of neonicotinoids (2 ng/ml compared to the LC50 dose of 4.28 µg/ml) impairing foraging behaviour, cognition, navigation, weakening of the immune system, and reducing reproductive success and genetic diversity within a colony^9,12–23^. Neonicotinoids, which target cholinergic neurotransmission, inhibit mushroom body Kenyon cell activity^24^ and disrupt memory and olfactory sensory neuron activity in pollinators, including bumblebees and honey bees (Apis mellifera), as well as fruit flies (Drosophila melanogaster)^24–27^.
The sensitivity of insects to neonicotinoids varies considerably between species. For example, the 24-h 50% lethal concentration (LC50) of thiamethoxam (TMX) is 3.13 µg/ml for adult Drosophila^28^, whereas for honey bees it is 4.28 µg/ml^23^. Variation arises from differences in the repertoire of cytochrome P450 detoxification genes. Drosophila encodes 85 of these neonicotinoid-metabolising enzymes, whereas honey bees have only 46^29,30^. Additionally, honey bees have fewer antioxidant gene paralogs than Drosophila, which has an expanded antioxidant defence system. Since neonicotinoids can induce oxidative stress, weaker antioxidant capacity may contribute to the greater sensitivity^31^.
Another reason contributing to this species-specific susceptibility could be the composition of nicotinic acetylcholine receptors (nAChRs). nAChRs are essential for neurotransmission, synaptic plasticity, and neuronal development, making them prominent targets for neonicotinoid pesticides^32–35^. These pentameric cys-loop ligand-gated cation channels consist of α and β subunits forming homo- or hetero-pentamers. Ligand binding pockets, located at α-α or α-β interfaces (Fig. 1a), consist of loops A, B, and C from an α subunit and loops D, E, and F from the adjacent α or β subunit^36–38^. Importantly, β subunits lack the ability to form functional homo-pentamers due to the absence of loops A, B, and C^36,38^. Consequently, nAChR subunit composition affects their properties and determines susceptibility to agonists, such as acetylcholine or neonicotinoids. Although homo-pentamers potentially can bind five ligands, binding of one ligand in human α7 pentamer is sufficient to induce the full response^39^.Fig. 1. Highly conserved Drosophila β2 subunit originated from an α subunit in other insects. (a) Conceptualised schematic of insect nAChR receptors illustrating possible examples of α–α and α–β homo- and heteropentameric assemblies (α: blue, β: pink). Cyan coloured circles indicate potential ligand-binding sites, which form at α–α or α–β interfaces depending on subunit arrangement. (b) Amino acid alignment of Dβ2 and honey bee Amelα8 subunits. Identical amino acids are marked in red, complementary amino acid substitutions are labelled in orange, and non-complementary amino acid substitutions are unmarked. The ligand binding domain is underlined. (c) Molecular phylogenetic analysis of all nAChR subunits present in Drosophila and honey bee with two major α subunits clades highlighted with light blue and pink (subunits highly conserved with mammalian α7). Dβ2 and Amelα8 (bold) clade is highlighted in darker blue. Species codes are Dm: D. melanogaster, Amel: A. mellifera. (d) Structural alignment of Drosophila Dβ2 (blue) and honey bee Amelα8 (pink) with all amino acid differences in D. melanogaster ligand binding domain indicated (cyan).
Compared to vertebrates, the majority of insects have smaller nAChR gene families^40–42^. D. melanogaster encodes 10 subunits (Dα1-Dα7 and Dβ1-Dβ3), the honey bee encodes 11 (Amelα1-Amelα9 and Amelβ1-Amelβ2)^43^, and the cockroach Periplaneta americana has 19^44^. These subunits are classified as α or β based on the presence or absence of two adjacent cysteines in loop C of the ligand binding domain (LBD) which are crucial for ligand interaction^45^.
Insect nAChR genes include both highly conserved and more divergent members^40,41^. Among these conserved subunits is the nAChRα8 present in many insects but notably absent in certain Dipterans, including D. melanogaster^41^. Instead, Drosophila encodes the Dβ2 subunit, which exhibits 75% overall sequence identity to the Amelα8 subunit, with striking 92% identity within the LBD (Fig. 1b and Supplementary Fig. 1). Notably, the absence of loop C in Dβ2 which is essential for ion channel gating, highlights a structural divergence that may contribute to species-specific functional properties and differences in neonicotinoid susceptibility^46,47^. Further, specific amino acid substitutions in nAChR subunits are known to impact insecticide susceptibility. For instance, Drosophila mutants lacking the Dβ1 gene or carrying the variant R81T in the Dβ1 subunit originally found in aphids exhibit about 10 and 100-fold increase in resistance to clothianidin and thiamethoxam, respectively^48–50^.
Here, we examine the phylogenetic relationship and functional implications of nAChR subunits in Drosophila melanogaster and Apis mellifera. Phylogenetic analysis confirms that Dβ2 and Amelα8 are orthologous, suggesting a conserved ancestral origin despite their classification as β and α subunits, respectively (Fig. 1c). To investigate whether this α-to-β subunit divergence contributes to neonicotinoids sensitivity, we employed CRISPR-Cas9 gene-editing technology to generate chimeric Drosophila flies expressing a chimeric nAChR receptor. Specifically, we replaced the LBD of the Dβ2 subunit with that of the Amelα8 subunit, providing a model to explore cross-species receptor functionality. Behavioural and survival assays revealed significant impairments in motor functions and altered sensitivity to insecticides, highlighting how subtle structural differences within the ligand-binding domain of a single nAChR subunit can influence pesticide response. Our results provide insights into the evolutionary trajectories of nAChR subunits and their roles in mediating neurotoxic compound interactions, establishing a valuable Drosophila model for investigating cross-species pesticide toxicity.
Results
Species-specific structural differences in the ligand-binding domains of Drosophila β2 and honey bee α8 nAChR subunits
To investigate whether Dβ2 represents a divergent lineage from other β subunits or shares a closer evolutionary relationship with α subunits, we used Alphafold structural modelling. Our analysis confirms overall structural conservation between Dβ2 and Amelα8, particularly in the LBD, with the exception of the loop C, which is absent in Dβ2 (Fig. 1d)^51^.
nAChR subunits exhibit developmental dynamics and cell-specific distribution
To investigate the expression patterns of nAChR subunits, we analysed tissue specific expression data in adults, taking into account enrichment values providing the gene abundance measure in a tissue relative to that in the whole fly^52^. The majority of nAChR subunits exhibits higher expression levels in females than in males (Fig. 2a). In both sexes, nAChR subunits are predominantly expressed in the central nervous system (CNS) and thoraco-abdominal ganglion, with Dβ2 being the most highly expressed subunit in the brain. In other tissues, however, the levels of expression differ between sexes, with Dβ2 and Dα7 being the most expressed subunits in the thoraco-abdominal ganglion of males and females, respectively. Furthermore, only two subunits, Dα4 and Dβ3, are expressed across all tissues, with Dβ3 showing notable expression in the heart. Specific differences were observed between sexes. For instance, in the male salivary glands, carcass, and crop, only Dα1, Dα2, and Dα3 are expressed, while Dα4, Dβ1, and Dβ3 are restricted to females.Fig. 2Drosophila nAChRβ2 is highly expressed in adult Drosophila CNS but not in every cell. (a) Expression levels of all Drosophila nAChR subunits across various adult tissues in males (left) and females (right) obtained from FlyAtlas 2. Receptor subunits are indicated on top and tissues on the left. The colour scale bar at the bottom indicates transcript abundance in fragments per kilobase of transcripts per million mapped reads (FPKM). (b) Single-cell RNA-seq visualisation of nAChRβ2 gene expression from ASAP in whole adult fly showing in red expression of Dβ2 on top of all cells shown in grey. Selected annotated cell clusters are indicated. The colour scale bar at the bottom indicates transcript abundance in fragments per kilobase of transcripts per million mapped reads (FPKM). (c–e) Single-cell RNA-seq visualisation maps showing qualitative expression of nAChRβ2 (red, c) on top of all cells (grey, c), the panneuronal marker elav (blue, d) or both (e) on top of all cells (grey) in the adult fly heads from ASAP. Selected annotated cell clusters are indicated. (f–k) Expression of nAChRβ2 visualised with a GAL4 inserted in the endogenous locus in adult Drosophila brains using UAS nuclear localised YFP reporter (f–h) or using UAS cell-membrane associated GFP reporter (i-k). Dβ2 expression (f, i) overlaps with the neuronal marker elav (g, j), as shown on merged pictures (h, k), confirming Dβ2 expression in neuronal populations. Scale bar is 20 µm.
Further, we investigated whether the expression of nAChR subunit changes during development. Larval tissue analysis revealed that Dα3 is not expressed in any tissue (Supplementary Fig. 2a), suggesting it is dispensable during development. In contrast, Dβ3 is expressed in all larval tissues, with the garland cells showing the highest expression. The larval CNS is the only structure expressing all nAChR subunits, with Dβ3 showing the highest levels. Overall, nAChR subunit expression is relatively low in most tissues in both larvae and adults (Fig. 2a and Supplementary Fig. 2a).
To refine the spatial expression of β2 in Drosophila cells in more detail, we analysed expression at single-cell resolution^53^. In whole adult flies, Dβ2 was found to be expressed in various cell types, including neurons, sensory neurons, glial cells, as well as male germline and reproductive system, however, its expression is not ubiquitous (Fig. 2b). Notably, a prominent cluster of neurons classified as adult fly head CNS cells strongly expressed Dβ2, suggesting it may have a significant role in neuronal circuits^26,54^. Since Dβ2 displayed the highest expression levels in the CNS, we further analysed single cell expression data for the larval brain and whole adult fly head. In the adult brain, Dβ2 expression was observed in distinct neuronal populations, including Kenyon cells, dopaminergic neurons or haemocytes, contrasting with the broader expression of the neuronal marker elav (Fig. 2c–e)^55^. In contrast, larval Dβ2 was colocalised with elav in the majority of CNS neurons (Supplementary Fig. 2b–d)^56^.
To validate the expression patterns, we used nAChRβ2^[2A-GAL4]^ allele which has a T2A-GAL4 sequence fused at the 3′ end of the gene, resulting in GAL4 expression as a separate protein under the control of nAChRβ2 regulation^57^. Therefore, we analysed both larval and adult brains using nuclear UAS-Histone2B::YFP or membrane-bound UAS-mCD8::GFP reporters, co-staining neurons with anti-elav antibody. Consistent with our observations from the single cell expression analyses, Dβ2 expression was restricted to specific neuronal subtypes, confirming its non-ubiquitous distribution (Fig. 2f–k and Supplementary Fig. 2e–j). Furthermore, we noticed enhanced Dβ2 expression in Kenyon cells of the mushroom body and the ellipsoid body of the central complex, particularly when a membrane-bound GFP reporter was used (Fig. 2i–k)^54^.
Together, these findings reveal that nAChR subunit expression is dynamically regulated during development and exhibits sex- and cell-specific specialisation, highlighting the functional diversity of these receptors in neural circuits and other tissues. Furthermore, the observed developmental shift in Dβ2 expression, from widespread larval CNS expression to more specialised adult neuronal populations, reflects the transition from simpler larval neural circuits to the more specialised and complex adult CNS.
Chimeric Amelα8/Dβ2 ligand-binding domain flies are viable with impaired motor functions
Since the two adjacent cysteines in the loop C are essential for ligand binding in nAChRs, we hypothesised that this difference may influence neonicotinoid susceptibility, making honey bees more vulnerable. Therefore, we generated chimeric nAChRβ2/α8 (nAChRβ2^Amα8LBD^) flies, where the β2 LBD coding region in Drosophila was swapped with the Amelα8 LBD coding sequence using CRISPR-Cas9 genome engineering (Fig. 3a and Supplementary Fig. 3). Successful recombinant flies were marked with white + selection marker, disrupting the LBD, therefore resulting in a null allele (nAChRβ2^nullA^). The marker was then excised by PiggyBac transposase to restore the open reading frame and generate flies expressing the chimeric nAChRβ2/α8 subunit (nAChRβ2^Amα8LBD^).Fig. 3. Chimeric nAChRα8β2 flies exhibit locomotor defects. (a) Schematic of the endogenous Dβ2 locus showing CRISPR-mediated replacement of the native ligand binding domain with the corresponding region from A. mellifera nAChRα8 and the white + (w +) selection marker. Coding exons are marked in pink. The w + marker and bee LBD sequence were inserted through homology-directed repair, disrupting the reading frame and generating a nAChRβ2 null allele. The reading frame can be restored by excising the PiggyBac transposon. (b) Climbing activity was assessed by negative geotaxis and is shown as means with the standard error from four biologically independent groups of 20 flies. Statistically significant differences are indicated by asterisks (***p = 7.510^–7^). (c) Flies of the indicated genotypes were tested for their flight ability shown as means with the standard error from four biologically independent groups of 10 flies. Statistically significant differences are indicated by asterisks (**p = 0.0066).
To normalise the genetic background, we used trans-heterozygotic flies for the nAChRβ2^attP^ allele (nAChRβ2^nullB^) in which the coding region was replaced with an attP site, 3xP3-RFP and a loxP site^57^, together with the nAChRβ2^nullA^ allele. The chimeric nAChRβ2^Amα8LBD^ mutants, as well as nAChRβ2^nullA^ and nAChRβ2^nullB^ flies are fully viable when crossed with a chromosomal deficiency (n = 453, 275, 421, respectively).
To assess motor abilities, we performed negative geotaxis assays (Fig. 3b) and evaluated flight ability (Fig. 3c). In negative geotaxis assays the climbing ability of LBD swap mutant (nAChRβ2^Amα8LBD^/nAChRβ2^nullB^) was greatly reduced (n = 4 from 20 flies each, p = 7.5*10^–7^) (Fig. 3b) compared to wild type (WT). We did not detect significant differences in the Dβ2 null mutant (nAChRβ2^nullA^/nAChRβ2^nullB^) nor the heterozygous mutant controls.
In flying ability assays, we observed the same, namely the LBD swap mutants (nAChRβ2^Amα8LBD^/nAChRβ2^nullB^) were impaired (n = 4 from 10 flies each, p = 0.0066) (Fig. 3c) compared to control flies, but not the β2 null mutant (nAChRβ2^nullA^/nAChRβ2^nullB^) or heterozygote mutant controls.
Chimeric Amelα8/Dβ2 ligand-binding domain flies display resistance to insecticides
To investigate the impact of substituting the Dβ2 LBD with the honey bee Amelα8 LBD on neonicotinoid susceptibility, we selected two well-characterised neonicotinoids thiamethoxam (TMX) and clothianidin (CLO) known for their high toxicity to pollinators^7,50^.
TMX is a prodrug that requires metabolic conversion to CLO to exert its full insecticidal effect^58,59^. CLO exhibits strong agonistic activity at insect nAChRs and slightly weaker toxicity than TMX (LC50 of 30.25 µM for TMX and 39.37 µM for CLO, Supplementary Fig. 4)^58,60^.
Next, we measured toxicity over time at different TMX and CLO concentrations. At 10 μM, wild type flies showed 43.75 and 47.5% survival at 144 h for CLO and TMX, respectively. In contrast, nAChRβ2^Amα8LBD^/ + , nAChRβ2^nullA^/ + , nAChRβ2^nullB^/ + , nAChRβ2^nullA^/nAChRβ2^nullB^, and nAChRβ2^Amα8LBD^/nAChRβ2^nullB^ flies remained viable (Fig. 4a,d).Fig. 4. The α8 ligand binding domain does not sensitise flies to neonicotinoids. (a–f) Viability of flies exposed to 10 µM (a, d), 20 µM (b, e) and 40 µM (c, f) clothianidin (a-c) or thiamethoxam (d-f) measured every 24 h for 6 days is shown as mean with standard error from four biologically independent groups starting with 20 flies. (g) Survival rate of nAChRβ2^Amα8LBD^/nAChRβ2^nullA^ and nAChRβ2^nullA^/nAChRβ2^nullB^ mutants compared to WT flies subjected to different insecticides combinations. The log2 fold-change shown as mean with standard error from four biologically independent groups starting with 20 flies represents the viability of flies after six days of treatment with TMX (p = 0.0429 for nAChRβ2^Amα8LBD^/nAChRβ2^nullB^ and* p* = 4.210^–5^ for nAChRβ2^nullA^/nAChRβ2^nullB^) or TMX mixed with flupyradifurone (FPF; p = 2.810^–5^ for nAChRβ2^Amα8LBD^/nAChRβ2^nullB^ and p = 1.3*10^–5^ for nAChRβ2^nullA^/nAChRβ2^nullB^) or imidacloprid (IMI; p = 0.0121 for nAChRβ2^Amα8LBD^/nAChRβ2^nullB^ and p = 0.0021 for nAChRβ2^nullA^/nAChRβ2^nullB^) in equimolar ratios. Statistically significant differences are indicated with letters (a = *p ≤ 0.5, b = **p ≤ 0.01, c = ***p ≤ 0.001, d = ****p ≤ 0.0001).
At 20 μM TMX and CLO, survival declined progressively across all genotypes. nAChRβ2^nullA^/nAChRβ2^nullB^ mutants showed the highest viability, followed by nAChRβ2^Amα8LBD^/nAChRβ2^nullB^ flies, which retained approximately 90% viability by day 6. In contrast, nAChRβ2^nullA^/ + (71.25% and 72.15%), nAChRβ2^nullB^/ + (60 and 73.75%), nAChRβ2^Amα8LBD^/ + (35% and 30%), and WT (11.25% and 7.75%) exhibited markedly lower survival (Fig. 4b,e).
At 40 μM TMX and CLO, all genotypes showed less than 50% viability by 144 h. nAChRβ2^nullA^/nAChRβ2^nullB^ mutants retained 48.75% (TMX) and 47.5% (CLO) survival, followed by nAChRβ2^Amα8LBD^/nAChRβ2^nullB^ (41.25% and 43.75%), nAChRβ2^nullA^/ + (35 and 37.5%), nAChRβ2^nullB^/ + (16.25% and 26.25%), nAChRβ2^Amα8LBD^/ + (3.33% and 7.5%), while WT flies exhibited complete lethality by day 4 (TMX) and 5 (CLO) (Fig. 4c,f). Interestingly, apart from the trans-heterozygous β^nullA^/β^nullB^ flies, all genotypes experienced a sharp drop in viability between the 1st and 2nd day of treatment with TMX, particularly nAChRβ2^Amα8LBD^/ + dropping from 62.5 to 30% and WT failing from 79 to 20%.
TMX and CLO primarily target nAChR subtypes containing Dβ1, and to a lesser extent, Dα1 and Dα3 subunits^50,61,62^. In contrast, imidacloprid (IMI) acts on receptor subtypes comprising Dα1, Dα2, Dβ1 and Dβ2 subunits, with lower affinity for those containing Dα4 and Dα7 subunits^50,61^. Flupyradifurone (FPF), a systemic butanolide compound chemically distinct from neonicotinoids representing a novel alternative to these compounds in pest management^63^. FPF acts on receptor subtypes having Dβ1 subunits and, to a lesser degree, to receptor subtypes having Dα3^50^ and exerts broad-spectrum activity and demonstrated toxicity to non-target pollinators, including bees^64,65^.
TMX, IMI and FPF act as nAChR agonists, bind to the same site and cause prolonged activation, they vary in the binding dynamics, receptor subtype preferences, and potential to develop resistance^66^. Given the different mechanisms of toxicity of TMX compared to IMI and FPF, we tested whether IMI and FPF would potentiate toxicity of TMX in combination. We exposed flies to a mixture of TMX (20 μM) with either FPF (20 μM) or IMI (20 μM). In nature, pollinators and insects are frequently exposed to multiple insecticides rather than a single compound, making it crucial to understand potential interactions and cross-resistance effects^67,68^. The TMX concentration was chosen based on prior survival analyses, where double mutant flies retained > 90% viability (Fig. 4e), while WT flies showed significant susceptibility. We tested chimeric nAChRβ2^Amα8LBD^/nAChRβ2^nullB^ flies and nAChRβ2^nullA^/nAChRβ2^nullB^ mutants to determine whether altered receptor composition influences responses to dual insecticide exposure. Flies were treated for six days, and viability on day six was analysed as a log-fold change relative to WT control and TMX-only treatment (Fig. 4g). Notably, exposure to TMX in combination with either FPF (p = 2.810^–5^ for nAChRβ2^Amα8LBD^/nAChRβ2^nullB^ and p = 1.310^–5^ for nAChRβ2^nullA^/nAChRβ2^nullB^) or IMI (p = 0.0121 for nAChRβ2^Amα8LBD^/nAChRβ2^nullB^ and p = 0.0021 for nAChRβ2^nullA^/nAChRβ2^nullB^) resulted in a slight increase in survival rates. However, the combined treatment did not greatly alter overall insecticide susceptibility. As expected, nAChRβ2^Amα8LBD^/nAChRβ2^nullB^ flies and nAChRβ2^nullA^/nAChRβ2^nullB^ mutants flies exhibited significantly higher resistance compared to WT across all conditions, including TMX-only treatment (p = 0.0429 for nAChRβ2^Amα8LBD^/nAChRβ2^nullB^ and p = 4.2*10^–5^ for nAChRβ2^nullA^/nAChRβ2^nullB^).
Taken together, our findings demonstrate that the β2^null^ (nAChRβ2^nullA^/nAChRβ2^nullB^) mutants, as well as the LBD swap (nAChRβ2^Amα8LBD^/nAChRβ2^nullB^) flies exhibit higher survival rates upon exposure to TMX, reinforcing the role of Dβ2 in neonicotinoid susceptibility. Further, exposure to combined neonicotinoid and butanolide treatments did not significantly alter overall lethality compared to TMX treatment on its own, indicating that sub-lethal doses of these nAChR-targeting insecticides do not act synergistically in this context. The observed resistance of chimeric and β2^null^ mutants further highlights the functional divergence of Amelα8 in modifying nAChR-mediated insecticide susceptibility.
Discussion
The agricultural industry relies on pesticides to protect crops and maintain a stable food supply for a growing population. Among these, neonicotinoids are the most widely used insecticides due to their high efficacy and systemic action, reducing the need for repeated applications. However, their persistence in plant tissues, including nectar and pollen, poses a serious threat to pollinators. Despite well-documented sublethal and lethal effects on bees and other beneficial insects, neonicotinoids remain the dominant insecticides worldwide^69^, highlighting the urgent need to mitigate their ecological impact.
Neonicotinoid toxicity varies widely across species. Drosophila melanogaster, for instance, is considerably more resistant than honey bees, despite homologous nAChR subunits^61^, raising the question of whether sequence identity between subunits is sufficient to explain these differences. To understand whether structural conservation confers insecticide resistance, we directly tested the contribution of the LBD to neonicotinoid sensitivity. Thus, we used CRISPR-Cas9 to generate a chimeric Drosophila mutant expressing a β2 subunit in which the LBD was replaced with that of honey bee Amelα8. Contrary to expectations, the resulting chimeric receptor did not sensitise fly viability to neonicotinoid exposure. However, the LBD swap mutant displayed motor function deficits, indicating that the chimeric receptor forms functional channels that affect motor behaviour. These findings demonstrate that sequence identity alone does not predict pharmacological response and is insufficient to recapitulate the function of homologous subunit.
Nevertheless, even minor structural changes in receptor subunits can profoundly impact pharmacological properties^36,38,49,50^. Specific amino acid substitutions in nAChR subunits are known to alter insecticide susceptibility as found for the R81T substitution in the aphid β1 subunit. Also Drosophila gene knockouts of individual subunits exhibit an increase in resistance to neonicotinoids and other insecticides^48–50,70^. More broadly, species-specific differences in sensitivity likely result from variation in receptor composition, metabolism, or detoxification pathways^29,31^, though the relative contribution of each remains unclear.
Functional diversity in nAChR subunit expression adds further complexity. In honey bees, α2, α8 and β1 are expressed in Kenyon cells, while α7 is restricted to antennal lobe neurons^71^. In Drosophila, α1 and α6 contribute to dendrite morphogenesis and synaptic transmission in larval visual circuits^72^. Developmental reprogramming of nAChR expression, such as reduced Dα3 in larvae and stage-specific expression profile shifts of subunits, may support behavioural role-dependent specialisations of different subunits, including reproduction^62,73^. Furthermore, plasticity is not restricted to development. Subchronic exposure to sub-lethal doses of imidacloprid in Periplaneta americana induces a significant decrease in α2 mRNA expression, and reduces sensitivity to the insecticide^74^, highlighting potential adaptive changes in nAChR subunit composition even in fully developed insects.
Furthermore, high Dβ2 expression in CNS-specific neuronal populations, including Kenyon cells, suggests its role in higher-order functions such as learning and memory^54^. Similarly, Amelα8 is enriched in honey bee mushroom bodies^43,75,76^, suggesting a conserved role across insect taxa, despite species-specific specialisations. Notably, neonicotinoid exposure disrupts cell activity in mushroom body and impairs learning and memory in insects, including Drosophila and honey bees^24–26,77^, further highlighting the critical role of nAChRs in cognitive functions.
Pharmacological and genetic studies have identified α1, α2, β1, and β2 as the primary mediators of neonicotinoid toxicity^40,61,78^, however, identifying the molecular determinants of binding and downstream signalling underlying species-specific effects require further investigation of receptor stoichiometry and subunit interfaces. Our finding that the α8β2 chimeric mutant did not exhibit increased sensitivity indicates that homology at the LBD level is insufficient to account for species-specific pharmacological differences. This challenges the assumption that sequence similarity alone is predictive of toxicological response. Instead, interactions at subunit interfaces, receptor assembly dynamics, and accessory proteins may be critical determinants of response. Clarifying these features is essential for understanding what governs neonicotinoid binding and efficacy across taxa.
In conclusion, our study establishes a framework for investigating molecular determinants of neonicotinoid sensitivity of insects and vertebrates nAChR using a cross-species chimeric nAChR Drosophila model. This model can contribute to the design of next generation of insecticides for enhanced species-specific responses with implications for pollinator conservation and pesticide safety for vertebrates including humans.
Materials and methods
Fly stocks, genetics, immunostaining of tissues and imaging
D. melanogaster CantonS and w^1118^ were used as the wild type control. nAChRß2^attP^ (BDSC 84545) and nAChRβ2^[2A-GAL4]^ (BDSC 84666) stocks were described previously^57^ and were together with the chromosomal deficiency (BDSC 24996) from Bloomington. Fly crosses were maintained at 25 °C in plastic vials containing 10 ml of a standard cornmeal/yeast-rich medium (1% agar, 2% yeast, 7% dextrose, 8% cornmeal w/v and 2% Nipagin from a 10% solution in ethanol) with a 12:12 h light–dark cycle.
Third instar wandering larvae and adult brains from the progeny of UAS-Histone2B::YFP/ + ;nAChRβ2^[2A-GAL4]^/ + and UAS-mCD8::GFP/ + ;nAChRβ2^[2A-GAL4]^/ + were dissected in phosphate buffered saline (PBS) and fixed in 4% paraformaldehyde in PBT (PBS with 0.1% TritonTM X-100 (Sigma-Aldrich, T8787)) for 30 min, followed by washes in PBT 3 × 15 min. Samples were incubated overnight at 4 °C with primary mouse anti-elav antibodies (MAb 7D, 1:20)^79^, followed by secondary antibodies conjugated with Alexa Fluor 546 again overnight at 4 °C. Samples were counterstained with DAPI (1:1000), mounted in Vectashield (Vector Labs), scanned with Leica SP8, and processed using FIJI.
Sequence analysis and single cell expression data visualisation
Amino acid sequences were aligned using ClustalW with Megalin (DNAstar) or with MAFFT^80^ and phylogenetic trees were generated in NGPhylogeny.fr^81–83^ and visualised using iTOL^84^.
Raw whole-genome sequencing reads were assessed for quality using FastQC, followed by adapter trimming and quality filtering with Trim Galore. High-quality reads were aligned to the Drosophila melanogaster (dm6) and Apis mellifera (Amel_HAv3.1) reference genomes using BBMap. Mapping quality and coverage were evaluated, and alignments were visualised using IGV (Integrative Genomics Viewer).
Single cell expression data was visualised as t-distributed stochastic neighbour embedding (tSNE) from the 10 × Stringent dataset in Scope and ASAP^85^.
Generation of chimeric nAChRα8β2 flies
Two single guide RNAs (sgRNAs) flanking the ligand binding domain Dβ2 were designed using PlatinumCRISPr^86^ (Supplementary Fig. 3a,b) and cloned into pUC-3GLA using the following primers: nAChRα8 sgRNA left F1 (AAGATATCCGGGTGAACTTCGCTTATTGGAGCTAGGAAAGGTTTTAGAGCTAGAAATAGC) and nAChRα8 sgRNA right R1 (GCTATTTCTAGCTCTAAAACCACATGGCACAATCAAATTCGACGTTAAATTGAAAATAGG) as described previously^86^. Chimeric nAChR subunits were constructed by combining sequences of nAChRα8 from A. mellifera and nAChRβ2 from D. melanogaster in three steps (Supplementary Fig. 3c–e). Fragments used for generating the chimeric nAChR subunit were synthesised using PCR with primers Beeα8 F2 (GGGAGTGGTCATTGCCATCTCAACCCTATATAAATTTG) and Beeα8 R2 (GGAATGTAATACCCACGCAAGGAATTATTAAG), followed by Beeα8 F1 (GCCAGTGAATTCGAGCTCGGTACCGCAGCTAGCGAAGCAAATCCTGACACAAAGAGACTTTATGATGAC) and Beeα8 R1 (TTTACGCAGACTATCTTTCTAGGGTTAACCGTATAGAATAATGTTTTTCTTCGCATTG) for Amelα8 ligand binding domain, Dmel nAChRβ2 left F1 (CGGCCAGTGAATTCGAGCTCGGTACCGATCCTTTTAGATAAAACATTTAGGAGCTATC) and nAChRsgmut left R1 (CTCTTTGTGTCAGGATTTGCTTCGAAACTCACTGGAGCCGTGAGAGGAGGGGTAATTCAGGGGAAAAACAGAGAAAAATGCC) for Dβ2 left homology arm, and nAChRsgmut right F1 (ATTTTACGCATGATTATCTTTAACGTACGTCACAATATGATTATCTTTCTAGGGTTAATCTGATTGTGCCCTGCGTAGCTTTAACATTCC) and Dmelα8 right R1 (GCATGCCTGCAGGTCGACTCTAGAGGATCCTGATAGTTGCTGCTGCGAATGCGGAGCTG) for Dβ2 right homology arm, ensuring sgRNA sites in both homology arms are mutated. Mutations were introduced based on conservation of the region between closely related Drosophila species analysed using the UCSC genome browser (https://genome.ucsc.edu) as described previously^87^. Fragments were then cloned stepwise into the pUC19 pBac w + [accession number: PV267745] containing white + marker, disrupting the LBD, therefore resulting in nAChRβ2^null^ flies. The selection marker was flanked with inverted terminal repeats inserted at a TTAA PiggyBac motifs for the later scarless excision by PiggyBac transposase and restoring the open reading frame resulting in a fly stock expressing the chimeric nAChRβ2/α8 subunit (nAChRβ2^Amα8LBD^). The pUC19 pBac w + Amelα8/Dβ2 plasmid was treated with the nicking endonuclease Nb.BtsI (NEB, R0707S) prior to injection. Transgenic lines were generated by injection into a GFP nosCas9 carrying flies (kindly provided by FlyORF). A total of three injections were done with supercoiled plasmids (0.4 µg/µl repair template and 0.1 µg/µl sgRNA plasmid) yielding 186, 176 and 141 survivors with a fertility rate of 67, 62 and 55%, but no transformants were found. From one injection with a nicked repair template, we obtained 135 survivors with a fertility rate of 65% and we got two G0 flies with transformants. Positive transformants were identified by red eye colour. Flies were validated by PCR, Sanger sequencing, and subsequently by whole genome sequencing.
For Drosophila whole genome sequencing, large fragment DNA was extracted with the Quick-DNA Tissue/Insect Miniprep Kit (Zymogene) according to the manufacturers’ instructions but replacing the BashingBead lysis step with cracking flies in liquid nitrogen followed by gentle homogenization with a pestle in the provided BashingBead Buffer. Illumina sequencing was done by Novogene and the sequence submitted to GEO [PRJNA1233772].
Locomotive assessment
Negative geotaxis experiments were done as previously described^88^. Briefly, two to five day-old flies of both sexes were collected with CO_2_ anaesthesia and grouped in sets of 20. Flies were allowed to recover for a day and then placed in two inverted fly vials (19 cm). Flies were tapped to the bottom, and their climbing behaviour was recorded on video. Every 5 s for a total of 30 s, the distance climbed by each fly was measured and recorded. The data was collected for four biological replicates.
For flying ability assessment, two to five day-old flies of both sexes were collected with CO_2_ anaesthesia and grouped in sets of 10. Flies were allowed to recover for a day before the flying test. Then, flies were tapped to a flat surface, and their ability to fly was recorded on video. Flies were observed over a 30 s period. At the end of 30 s, the number of flies that successfully flew away was recorded. The data was collected for four biological replicates.
Insecticide toxicity assay
Two to five day-old flies of both sexes, grouped in sets of 20, were exposed to various concentrations of clothianidin (Sigma-Aldrich, T21153), thiamethoxam (Sigma-Aldrich, 37,924), or a combination of 20 µM of thiamethoxam with 20 µM imidacloprid (Thermo Fisher, 466,752,500) or 20 µM flupyradifurone (MCE, HY-145295). The insecticides were dissolved in 0.01% acetone and added on top of the standard fly food, and flies were added to the vials after 24 h. A full dose–response survival assay was performed for the wild type genotype only. Flies were exposed to thiamethoxam and clothianidin concentrations ranging from 2.5 to 80 µM (Supplementary Fig. 4). A generalised linear model (GLM) was used to estimate the relationship between dose and mortality, yielding an LC50 of 30.25 µM for thiamethoxam and 39.37 µM for clothianidin (see Statistical analysis section). Mutant genotypes were tested at fixed concentrations based on the wild type response and ranged from 10 to 40 µM for all genotypes. Flies were exposed for 6 days, and survival was assessed every 24 h.
Statistical analysis
Statistical analyses were carried out using GraphPad Prism 9 and R (v4.4.1). One-way ANOVA followed by Tukey’s post hoc test was used for behavioural assays. For insecticide toxicity, GLM with a binomial error distribution and logit link were fitted to mortality data (cbind(dead, alive) ~ conc) using the glm() function in R. Model fit was assessed using residual deviance and AIC. Significance of the concentration effect was determined using Wald tests. To test for synergistic effects, survival under combined treatments (TMX + IMI or TMX + FPF) was compared to survival under TMX-only exposure using two-way ANOVA followed by Tukey’s test for pairwise comparisons. All data represent a minimum of three independent replicates conducted on separate days.
Supplementary Information
Supplementary Information.