Postbiotic Supplementation Increases Amino Acid Absorption from Plant-Based Meal: A Placebo-Controlled, Randomized, Double-Blind, Crossover Study
Christine M. Florez, Javier Zaragoza, Jessica Prather, Mandy Parra, Jaci Davis, Amie Vargas, Audrey Ross, Ralf Jäger, Martin Purpura, Simone Guglielmetti, Grant M. Tinsley, Lem Taylor

TL;DR
This study found that postbiotic supplementation improves amino acid absorption from plant-based meals compared to a placebo.
Contribution
This is the first study to show that postbiotic supplementation enhances amino acid absorption from mixed macronutrient meals.
Findings
Postbiotic supplementation significantly increased absorption of several amino acids and total amino acids compared to placebo.
Improvements were observed for alanine, asparagine, citrulline, cystine, glycine, methionine, and proline.
Abstract
Supplementation of probiotic strains can enhance the absorption of amino acids from protein in the gut. The purpose of this study was to assess if supplementation of a multi-strain probiotic or a postbiotic, consisting of the same strains, would alter the absorption of individual and total amino acids following ingestion of a plant-based meal. Sixteen male participants consumed either probiotic (PRO) or postbiotic (cells inactivated by γ-irradiation; POST), both consisting of L. paracasei LP-DG® (CNCM I-1572) plus L. paracasei LPC-S01 (DSM 26760), or a placebo (PLA) for 2 weeks in a randomized, double-blind, crossover design study separated by a 4-week washout period. During the testing session, blood samples were taken at baseline, 30-, 60-, 120-, and 180-min post-ingestion of a plant-based vegan burger patty. Plasma amino acid levels were analyzed, and percent changes from baseline…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
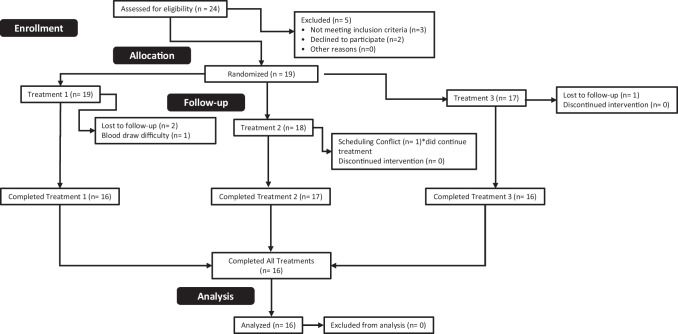 Figure 1
Figure 1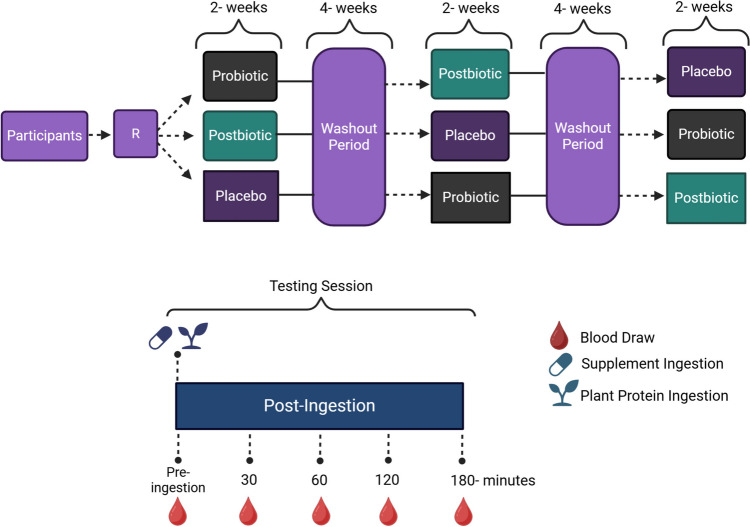 Figure 2
Figure 2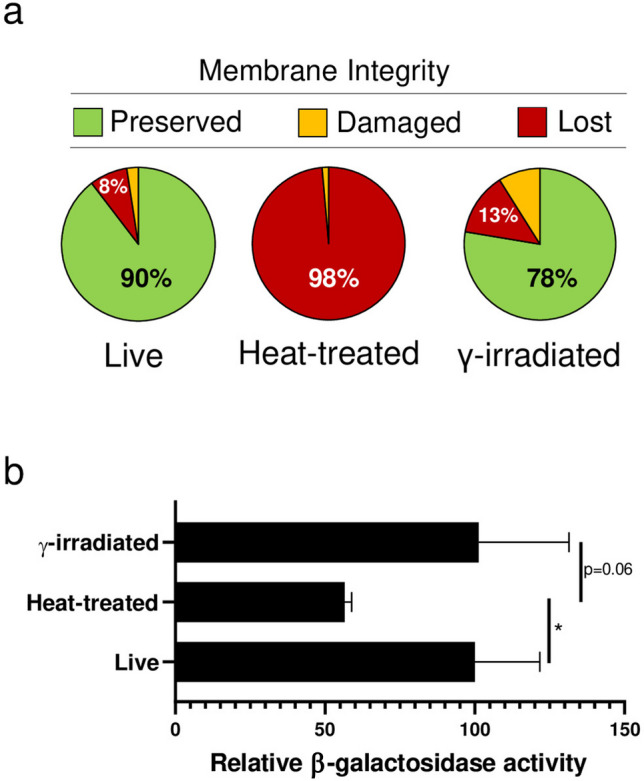 Figure 3
Figure 3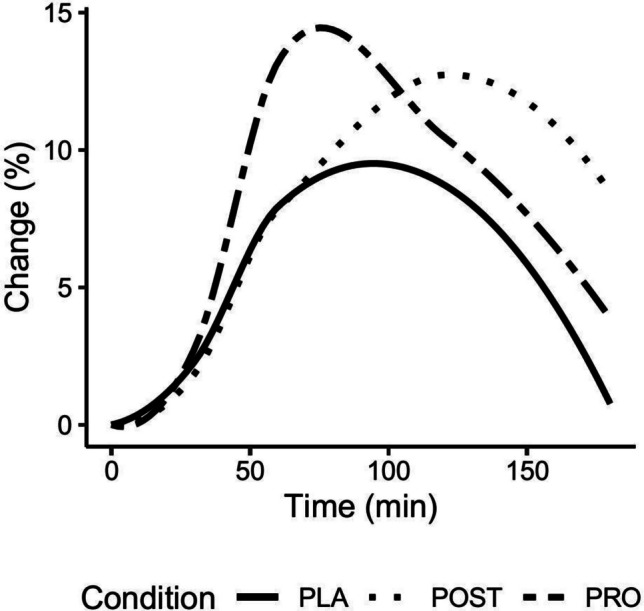 Figure 4
Figure 4- —Sofar S.p.A., Italy
- —University of Mary Hardin-Baylor
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAgriculture Sustainability and Environmental Impact · Consumer Attitudes and Food Labeling · Probiotics and Fermented Foods
Introduction
Probiotics are living microorganisms that can confer health benefits when administered in adequate amounts [1, 2]. Probiotic supplementation has been shown to elicit a myriad of benefits to the host; however, for a probiotic to impact health, it must survive the GI environment, and the magnitude of its effects is strain- and dose-dependent [3]. Moreover, many live microorganisms used as probiotics are sensitive to heat and/or oxygen, leading to challenges in sustaining their shelf-life. In fact, to offset the expected decrease in viable cells at the end of shelf-life, manufacturers often include more than the stated quantity [4]. Fermented foods such as kimchi, sauerkraut, yogurt, and kombucha also possess health-promoting bacteria; however, the number of bacteria in food products is not standardized. Furthermore, the bacteria in these foods, like probiotics, have limited shelf-life. For this reason, alternative methods of extending probiotic shelf-life have been explored, leading to the development of postbiotics [5, 6].
A broad consensus on the definition of postbiotics has not yet been established [7]. However, postbiotics must contain intact cells or cell fragments and are produced from killing bacteria through a deliberate killing process. They are “preparations of inanimate microorganisms and/or their components that confer a health benefit on the host.” [8]. These inactivated cells may offer an attractive alternative relative to safety [9] and stability, when compared to their probiotic counterparts [5]. The process of inactivation can be achieved in several ways, including thermal sterilization, pasteurization, or irradiation [5]. Regardless of their preparation method, postbiotics are thought to be potentially as effective as probiotic supplementation [10]. Of the purported benefits, certain strains of probiotics have been observed to support digestive health [11], positively modulate immune system function [12, 13], and aid in gastrointestinal protein absorption [14]. Research in probiotic and postbiotic supplementation continues to gain interest with an emerging focus on its potential to improve protein absorption efficiency.
Dietary protein intake is essential to basic human physiology and influences tissue-building, body composition, and immune function [15]. Plant proteins are typically considered to be low-quality when compared to animal sources due to their amino acid profiles [16]. However, despite this classification, the increased consumption of plant-based proteins is cited as a nutritional strategy to improve overall health [17]. Additionally, many individuals have opted to reduce meat consumption or avoid it altogether. A worldwide survey in 2019 revealed that 40% of those surveyed admitted to intentionally reducing meat consumption in favor of alternative protein sources [18]. In the USA, the plant-based meat market is anticipated to grow by several billion dollars [19]. Due to increased interest in plant-based diets, research to improve the bioavailability of plant-based proteins is currently being conducted [20]. One such strategy is the co-ingestion of bacterial cells alongside plant-based proteins to improve amino acid bioavailability.
Lacticaseibacillus paracasei is a species of bacteria frequently included in commercially available supplements [21]. Strains from this species have been previously reported to aid in improving the gut microbiota environment [22–24]. Additionally, we have previously demonstrated that L. paracasei DG and L. paracasei LPC-S01 increased amino acid absorption from pea protein powder when compared to a placebo [20]. Therefore, the purpose of this study was to assess if supplementation of a multi-strain probiotic or a postbiotic consisting of inactivated cells of the same strains would alter the absorption of individual and total amino acids following ingestion of plant-based meals. It was hypothesized that both probiotic and postbiotic supplementation would enhance amino acid appearance relative to placebo.
Methods
Probiotic and Postbiotic Preparations
Three microbial preparations were evaluated, each comprising a blend of industrially lyophilized biomasses from two Lacticaseibacillus paracasei strains: DG (L. casei DG®, CNCM I-1572) and LPC-S01 (DSM 26760). One preparation contained viable bacterial cells, another included bacteria inactivated by heat treatment (80 °C for 30 min), and the third consisted of bacteria inactivated by γ-irradiation (10 kGy, in accordance with ISO 11137). Only the viable (probiotic) and γ-irradiated (postbiotic) preparations were used in the human trial. The postbiotic, probiotic, and corresponding placebo (maltodextrin) were provided in sachets by Sofar S.p.A., Italy. The probiotic sachets contained 10 billion colony-forming units (CFU), while the postbiotic sachets contained 10 billion active fluorescent units (AFU), 5 billion of each individual strain, respectively.
Evaluation of Membrane Integrity
The membrane integrity of bacteria was evaluated following labeling with SYTO™ 24 and propidium iodide using flow cytometry (BD Accuri™ C6 flow cytometer, BD Biosciences, Milan, Italy). One gram of lyophilizate was resuspended in PBS buffer to obtain a 1:10 dilution. Following homogenization, the cell suspension was subjected to serial decimal dilutions in filtered PBS buffer, using 1.5-mL Eppendorf tubes. These dilutions were used to determine vitality by setting the following instrumental parameters: sample acquisition volume: 50 µL and threshold: FSC: 4000 and SSC: 1000. The flow cytometric analysis involved an initial acquisition of the unlabeled sample to identify the appropriate decimal dilution for subsequent staining, ensuring that the number of events/microliter was within the range of 1000 to 3000. After determining the correct dilution, cell labeling was performed by adding 50 µL of the sample and 10 µL of the SYTO™ 24 marker to 440 µL of diluent (filtered PBS) (absorption (nm): DNA (490), RNA (ND)) along with propidium iodide (PI). The total number of viable cells was then determined by double labeling the samples in accordance with protocol B of the ISO 19344:2015 method (IDF 232).
Evaluation of β-Galactosidase Activity
The β-galactosidase activity was assessed using a colorimetric test on the protein extract obtained from cell lysis of the freeze-dried final product. Specifically, 1 g of freeze-dried product was resuspended, washed twice in 20 mL of 0.1 M phosphate buffer (pH 7), and then resuspended in a final volume of 5 mL. Following cell disruption via bead beating (4 cycles of 20 s at 6300 RPM), the suspension was centrifuged at 13,000 RPM for 45 min at 5 °C. After protein quantification using the Bradford assay, a 100 μL aliquot of the protein extract was added to 130 μL of phosphate buffer and 100 μL of o-nitrophenyl-β-galactosidase (ONPG) (3 mg/mL) and incubated for 2 h at 37 °C, monitoring the change in absorbance at 420 nm. The enzymatic activity was expressed as a relative unit, calculated by comparing the enzymatic units of the sample with those of the “non-inactivated” sample of the respective product.
Overview of Human Trial
A randomized, double-blind, crossover study was performed to assess the amino acid concentration in the blood after the administration of a plant-based burger containing 20 g of protein from pea protein isolates, rice protein, and mung bean protein with co-administration of probiotics or postbiotics vs. placebo. The subjects were asked to consume one sachet in the morning with 240 mL of water. The University of Mary Hardin-Baylor Institutional Review Board approved this study (IRB number: #114 on March 9, 2022). Participants were informed of the rationale and purpose of this study and their right to refuse or discontinue participation at any given time throughout the study. All participants provided written informed consent prior to participation.
Participants
Sixteen normal-weight recreationally active males ([mean ± SD] age: 23.1 ± 3.2 years; height: 180.3 ± 7.7 cm; weight 86.6 ± 13.8 kg; body fat 17.8 ± 6.3%) were recruited for the present study. Subjects did not consume any nutritional or ergogenic supplement known to affect measures of the current study for 6 weeks prior to participation, including probiotics, prebiotics, and digestive enzymes. Exclusion criteria included any individual who was being treated for or diagnosed with a gastrointestinal, cardiac, respiratory, circulatory, musculoskeletal, metabolic, immune, autoimmune, psychiatric, hematological, neurological, or endocrinological disorder. Additionally, participants who were determined to not be weight stable, defined as measured body mass deviating by 2% or more between trials, and participants who were not willing to abstain from alcohol, nicotine, and caffeine for 12 h prior to each visit were excluded (Fig. 1).Fig. 1. Consort chart
Study Design
Three supplementation periods that each spanned 2 weeks were completed and separated by a washout period of 4 weeks (Fig. 2). For each study visit, all participants reported to the laboratory between 06:00 and 9:00 h after an 8- to 10-h fast. Their diet was recorded, and subjects were asked to repeat the same diet for the 2 weeks leading up to the second and third experimental testing sessions.Fig. 2. Study scheme for the intervention trial with the probiotic (PRO), postbiotic (POST), and placebo (PLA) preparations
Participants were randomly assigned to ingest either a placebo (PLA), a postbiotic (POST), or a probiotic (PRO) product for 2 weeks. On testing days, biotics were ingested fasted with 240 mL of water (or whatever the standard procedure was), while on non-testing days, the biotics were ingested upon waking with breakfast with 240 mL of water. Upon arrival for each study visit, participants had their resting heart rate, blood pressure, body mass, height, and body composition (InBody 770, InBody, Seoul, South Korea) measured.
Testing Session
Subjects rested semi-supine for placement of a Teflon catheter into an antecubital vein for multiple blood sampling. The catheter was kept patent by flushing with 2–3 mL of 0.9% sodium chloride. Following baseline sampling, participants ingested their respective treatments and then consumed a vegan burger (Beyond Burger, Beyond Meat, El Segundo, CA). Thereafter, blood samples were collected at 30-, 60-, 120-, and 180-min post-ingestion (Fig. 2). Subsequently, a 4-week washout period was implemented, followed by the opposite condition.
Vegan Burger Preparation
A single vegan patty (20 g protein, 14 g fat, and 7 g carbohydrate) was prepared in the lab on an indoor grill. The patty was cooked until the internal temperature reached 165° Fahrenheit as measured by a meat thermometer and served to participants plain.
Outcome Variables
Dependent variables for the amino acid appearance portion of the trial included total amino acids, essential amino acids (EAAs), branched-chain amino acids (BCAAs), and individual amino acids (arginine, glutamine, citrulline, serine, asparagine, glycine, threonine, alanine, ornithine, methionine, proline, lysine, aspartic acid, histidine, valine, glutamic acid, tryptophan, leucine, phenylalanine, isoleucine, cysteine, and tyrosine) in peripheral blood. The maximum observed concentration (Cmax) and corresponding time (Tmax) and incremental area under the curve (iAUC) were calculated for each amino acid. Safety was assessed with a complete blood count (CBC) and comprehensive metabolic panel (CMP) at all pre- and post-time points.
Amino Acid Analysis
Amino acid analysis was performed by Heartland Assays, Iowa State University Research Park, Ames, IA, USA. EZ:faast® amino acid analysis kits (Phenomenex, Torrance, CA) were used for liquid chromatographic analysis of amino acids using tandem-mass spectrometry (LC/MS/MS) and electrospray ionization (ESI). The procedure consisted of solid phase extraction of 25 µL of plasma with internal standards by a sorbent tip attached to a syringe with an eluting solvent (a 3:2 mixture of sodium hydroxide with 77% n-propanol and 23% 3-picoline). The free amino acids were then derivatized by adding a mixture of 17.4% propyl chloroformate, 11% isooctane, and 71.6% chloroform. The resulting mixture was vortexed and allowed to sit at room temperature for 1 min, followed by liquid–liquid extraction with isooctane. Fifty microliters of the organic layer was removed, dried under nitrogen gas, and suspended in the HPLC run solvents before being injected into the LC/MS/MS. Chromatographic separation of the derivatized amino acids was conducted on an EZ:faast amino acid analysis-mass spectrometry column (250 × 2.0 mm i.d., 4 µm) using an Agilent 6460 triple quadrupole LC/MS/MS system (Santa Clara, CA). Ten millimolar of ammonium formate in water with 0.2% formic acid (mobile phase A) and 10 mM ammonium formate in methanol with 0.2% formic acid (mobile phase B) were used as a solvent system with gradient conditions of 68% B at 0 min to 83% B over 13 min with a flow rate of 0.25 mL/min. Amino acids and internal standard data were collected using the Dynamic Multiple Reaction Monitoring mode using Mass Hunter acquisition software (Agilent, Santa Clara, CA). Mass Hunter Quantitation software was used to quantitate the unknown plasma samples based on best-fit standard curves [20].
Statistical Analysis
Seventeen individuals participated in the study. One participant was missing all data from all time points within one condition and was therefore excluded from the analysis. For the remaining 16 participants, the only missing data point was the 180-min time point for one participant within the placebo condition. These missing data were imputed using the last observation carried forward from the 120-min time point. Therefore, 16 individuals were included in the statistical analysis.
Changes in raw amino acid concentrations and percent changes from baseline were analyzed using linear mixed-effects models (nlme package [25], v. 3.1–157) with a random intercept for the participant. A first-order autoregressive (AR1) variance–covariance matrix was employed, using the correlation form of time | participant/condition. These models were fit by maximizing the restricted log-likelihood (REML). In all models, the reference condition was placebo (PLA). Model assumptions were examined through graphical methods (i.e., residuals vs. fitted plots and quantile–quantile plots). Model coefficients were examined using the SjPlot package [26] (v. 2.8.14) to determine the effects of each condition, time, and condition-by-time interactions.
In addition to linear mixed-effects models, pharmacokinetic calculations were performed. The incremental area under the concentration vs. time curve (iAUC) was calculated using the method of Brouns et al. [27]. The PKNCA package [28] (v. 0.10.0) was used to establish the maximum observed concentration (Cmax) and time of maximum observed concentration (Tmax). iAUC, Cmax, and Tmax values were analyzed via the Friedman rank sum test, using the rstatix package [29] (v. 0.7.0). When a Friedman test was significant, post hoc tests were performed using Wilcoxon signed-rank tests. For all tests, statistical significance was accepted at p < 0.05. Data were analyzed in R (version 4.2.1).
Results
Selection of the Postbiotic Formulation
The industrial bacterial biomass of the strain L. paracasei LPC-S01 was used to determine the most suitable method for producing the postbiotic for the subsequent human study. To this end, we compared the live, heat-inactivated, and γ-irradiated preparations in terms of cell membrane permeability to propidium iodide and intracellular enzymatic activity (β-galactosidase). Both heat treatment and γ-radiation completely inactivated the L. paracasei LPC-S01 biomass (CFU per gram of powder below the detection limit). Nonetheless, we found that ionizing radiation (γ rays) inactivated bacterial cells while preserving membrane integrity to a much greater extent than heat treatment (Fig. 3a). Additionally, the β-galactosidase activity in γ-inactivated cells was not significantly lower than that in viable cells, whereas in heat-inactivated bacteria, it was significantly reduced (Fig. 3b). Based on these findings, bacterial cells inactivated by γ-irradiation were selected as the postbiotic for the human trial, to be compared with the same live probiotic strains.Fig. 3. Comparison of lyophilized biomasses of Lacticaseibacillus paracasei LPC-S01 before (live) and after inactivation by heat treatment or γ-irradiation. a Cell membrane integrity assessed by flow cytometry following SYTO-9/propidium iodide staining, with data presented as the percentage of fluorescent units in the 2D pie chart. b β-Galactosidase activity assessed through the ONPG (ortho-nitrophenyl-β-galactoside) hydrolysis assay; statistical analysis was performed using an unpaired Student’s t-test (n = 3; *p < 0.05)
Amino Acid Concentrations
Relative to the reference model (i.e., PLA condition), there were statistically significant POST condition-by-time interactions for percent changes in alanine (0.07%/min [0.01–0.13]; p = 0.02), asparagine (0.08%/min [0.01–0.15]; p = 0.03), citrulline (0.06%/min [0.01–0.12]; p = 0.02), cystine (0.04%/min [0.01–0.08]; p = 0.02), glycine (0.08%/min [0.02–0.14]; p = 0.01), methionine (0.05%/min [0.01–0.10]; p = 0.02), proline (0.05%/min [0.0–0.10]; p = 0.04), and total amino acids (0.05%/min [0.0–0.09]; p = 0.04; Fig. 4, Table 1). Additionally, there was a statistically significant PRO condition-by-time interactions for cystine (0.05%/min [0.01–0.08]; p = 0.02). In addition to these condition-by-time interactions, significant effects of time were observed for alanine (p = 0.02), arginine (p = 0.02), BCAAs (p < 0.001), citrulline (p = 0.01), cystine (p < 0.001), EAA (p = 0.03), isoleucine (p < 0.001), leucine (p < 0.001), methionine (p < 0.001), ornithine (p < 0.001), and valine (p = 0.002). Percent changes in amino acid concentrations over time are displayed in Fig. 4 and Table 1. Raw amino acid concentrations and coefficients and p-values for percent change linear mixed-effects models are displayed in the supplementary materials.Fig. 4. Percent change in total amino acids. Percent changes in total amino acids following postbiotic (POST), probiotic (PRO), or placebo (PLA) are displayed. In the linear mixed-effects model, a statistically significant group by time interaction term was present for POST vs. PLA (p = 0.04), but not PRO vs. PLA (p = 0.56)Table 1. Changes in amino acid concentrationsAmino acidConditionMean percent changes (%)Condition by time interaction vs. PLA0–30 min0–60 min0–120 min0–180 minp-valueAlaninePLA − 2.11.21.5 − 7.7–POST − 3.02.99.33.70.02PRO1.611.55.1 − 3.90.79ArgininePLA21.337.442.118.4–POST29.143.446.227.50.60PRO27.748.841.422.50.92AsparaginePLA15.319.816.19.5–POST4.217.023.416.60.03PRO11.127.623.810.70.48Aspartic acidPLA18.628.728.818.4–POST30.718.832.045.10.67PRO27.6102.811.114.40.61BcaaPLA3.313.419.610.7–POST2.510.618.616.10.29PRO4.617.821.613.20.73CitrullinePLA4.11.1 − 5.3 − 5.3–POST3.70.4 − 0.35.60.02PRO2.02.91.6 − 1.50.18CystinePLA − 1.4 − 6.6 − 10.5 − 12.5–POST − 5.6 − 6.4 − 2.0 − 8.90.02PRO − 4.4 − 5.8 − 7.7 − 5.40.02EaaPLA3.812.114.66.0–POST2.910.617.012.10.13PRO5.017.717.98.50.65Glutamic acidPLA12.014.88.6 − 1.2–POST17.924.322.125.80.07PRO7.519.311.9 − 3.00.96GlutaminePLA − 3.11.02.1 − 2.7–POST − 1.22.26.04.00.16PRO − 2.74.42.72.00.42GlycinePLA1.93.52.1 − 4.2–POST − 0.42.99.07.70.01PRO2.212.55.30.20.51HistidinePLA1.12.33.50.9–POST − 1.43.810.66.00.08PRO1.49.16.91.70.88IsoleucinePLA8.224.533.819.2–POST6.020.134.729.20.17PRO9.429.339.025.50.42LeucinePLA3.417.223.312.7–POST5.616.421.421.10.33PRO6.823.826.715.80.81LysinePLA9.622.521.19.9–POST8.920.328.817.10.15PRO13.031.429.913.70.58MethioninePLA − 2.9 − 0.6 − 5.8 − 14.0–POST − 2.30.61.5 − 5.70.02PRO − 1.65.40.6 − 7.50.12OrnithinePLA21.738.248.737.3–POST15.531.248.047.40.21PRO24.343.545.538.50.84PhenylalaninePLA3.410.814.02.7–POST3.812.216.510.80.11PRO6.018.816.97.30.57ProlinePLA1.95.95.0 − 5.3–POST4.18.17.25.70.04PRO3.312.56.2 − 0.50.49SerinePLA11.612.111.01.6–POST3.78.013.011.20.07PRO10.319.413.210.30.28ThreoninePLA3.57.86.40.4–POST1.55.39.26.30.10PRO3.515.011.74.50.33Total AAPLA2.37.98.70.8–POST1.97.812.78.50.04PRO2.813.210.53.90.56TryptophanPLA1.13.92.6 − 7.7–POST0.79.75.81.50.12PRO − 1.94.9 − 1.2 − 7.40.99TyrosinePLA1.015.111.61.2–POST − 1.116.112.97.70.33PRO1.713.714.06.30.46ValinePLA1.88.013.26.9–POST0.04.812.69.80.37PRO2.011.413.88.30.83PLA* placebo, POST postbiotic, PRO probiotic*Denotes significance
When examining raw concentrations without accounting for baseline differences, the only statistically significant condition-by-time effects was for higher citrulline (0.02 μmol/L/min [0.0–0.04]; p = 0.04) and cystine (0.03 μmol/L/min [0.0–0.06]; p = 0.04) over time in the POST condition relative to the reference model. In addition to these condition-by-time interactions, significant effects of time were observed for arginine (p = 0.04), BCAA (p = 0.002), citrulline (p = 0.03), cystine (p < 0.001), EAA (p = 0.048), isoleucine (p < 0.001), leucine (p = 0.001), methionine (p = 0.01), ornithine (p < 0.001), and valine (p = 0.03). Coefficients and p-values for raw concentration linear mixed-effects models are displayed in the supplementary material.
Pharmacokinetic Analysis
Arginine iAUC values significantly differed by condition (p = 0.0498), with post hoc tests indicating higher iAUC in the POST condition as compared to PLA (p = 0.04; Table 2). Additionally, trends (0.05 < p < 0.1) were present for alanine (p = 0.07) and phenylalanine (p = 0.099) iAUC values. For Cmax, a significant Friedman test result was observed only for alanine (p = 0.047). However, post hoc tests did not reveal any significant differences between individual conditions (Table 3). For Tmax, significant effects of condition were observed for isoleucine (p = 0.03), methionine (p = 0.04), ornithine (p = 0.03), and total amino acids (p = 0.04). For isoleucine, post hoc tests indicated a significant difference between POST and PLA (p = 0.01), such that Tmax tended to occur later in POST as compared to PLA (Table 4). For methionine, post hoc tests did not reveal any significant differences between individual conditions. For ornithine, post hoc tests indicated a significant difference between POST and PRO (p = 0.02), such that Tmax tended to occur later in POST as compared to PRO. For total amino acids, post hoc tests revealed a significant difference between POST and PRO (p = 0.03), such that Tmax tended to occur later in POST as compared to PRO. Additionally, trends for significant Friedman tests were observed for phenylalanine (p = 0.08), proline (p = 0.06), and threonine (p = 0.09). Table 2. Incremental area under the curve (iAUC)Amino acidPostbioticProbioticPlacebop (Friedman test)Mean percent differencesMedian percent differencesMeanSDMedianIQRMeanSDMedianIQRMeanSDMedianIQRPOST vs. PLAPRO vs. PLAPOST vs. PLAPRO vs. PLAAlanine21293504883222024742722141623941418156990722370.0740.154.3 − 2.743.8Arginine27969422745.51313265410032482147723231003234516520.049818.513.315.75.7Asparagine6834066976097993117644726913806323270.65 − 1.314.59.718.9Aspartic acid1351756212919537078101104109711480.6525.760.9 − 14.49.5Bcaa5193262550562861592335494902535354413293540639490.83 − 4.78.5 − 6.7 − 9.8Citrulline171212513161511671002581281411052140.3628.816.9 − 69.8 − 5.4Cystine207784014621300353692040.98141.053.6NANAEaa10,197577811,059889511,15867118664979998135425927183460.783.812.817.6 − 6.8Glutamic acid6554985526175775234727794495193168210.2837.525.054.439.5Glutamine2920234829333044295929622350474820521981168535620.6134.936.254.033.0Glycine1325157278514511653201694318759217469496040.6536.056.9 − 18.9 − 0.7Histidine5384085566644834464202673473722074410.1443.332.891.567.9Isoleucine14687431390514165387614421436151586214638840.44 − 3.28.7 − 5.1 − 1.5Leucine2030100920071443219411971795157119671147184916980.943.110.98.2 − 2.9Lysine3118212730462098321315562685226630012508234919770.373.86.825.913.4Methionine1051156818810511384169679028990.8243.844.083.399.3Ornithine1437653150976414006771221902144260514697530.94 − 0.4 − 3.02.7 − 18.5Phenylalanine6413706394906834295376885553434513800.1014.420.734.617.5Proline1124928944121011551033951153991987162111310.7420.122.841.342.1Serine861670803637111998891213638265247447051.004.230.27.520.3Threonine8636487511051109584591313897825676674150.379.833.311.831.2Total AA22,22814,11820,38119,56621,38414,89914,49220,62818,64712,11615,14814,4310.4717.513.729.5 − 4.4Tryptophan3814332895091972401402323225011144350.5916.9 − 48.487.220.9Tyrosine6077355174786274556046036727574874940.83 − 10.2 − 7.06.021.5Valine1791108416361425210415301603244120361303209916230.72 − 12.83.3 − 24.8 − 26.8POST postbiotic, PRO probiotic, PLA placeboDenotes significanceTable 3Maximal observed concentrations (Cmax)Amino acidPostbioticProbioticPlacebop* (Friedman test)Mean percent differencesMedian percent differencesMeanSDMedianIQRMeanSDMedianIQRMeanSDMedianIQRPOST vs. PLAPRO vs. PLAPOST vs. PLAPRO vs. PLAAlanine375.998.5335.4125.6372.777.6368.2138.3361.390.4362.5146.00.0474.03.1 − 7.81.6Arginine122.024.9121.333.2124.519.1122.623.5125.424.5124.634.20.65 − 2.7 − 0.7 − 2.7 − 1.6Asparagine58.111.559.615.759.88.159.58.359.011.055.416.20.47 − 1.61.27.37.2Aspartic acid7.93.27.52.910.710.97.32.87.23.76.73.80.449.639.410.27.7Bcaa548.396.9537.466.4563.9115.5523.4170.0575.2124.4545.997.90.83 − 4.8 − 2.0 − 1.6 − 4.2Citrulline33.97.134.49.832.96.731.75.732.16.231.96.80.175.52.37.6 − 0.6Cystine72.326.261.827.372.916.871.428.971.018.966.720.40.301.72.6 − 7.76.8Eaa1157.6225.51111.7237.31168.6205.81132.6283.51190.0271.81151.5207.50.83 − 2.8 − 1.8 − 3.5 − 1.7Glutamic acid48.115.649.422.451.618.847.233.345.719.738.224.90.275.112.025.621.1Glutamine619.488.6631.0133.7646.6162.6614.570.1618.784.5607.8102.10.570.14.43.81.1Glycine244.168.8245.455.5252.753.6246.272.3242.364.7236.571.70.270.74.23.74.0Histidine88.412.986.822.587.112.484.615.991.119.385.921.60.94 − 3.0 − 4.51.0 − 1.5Isoleucine99.418.698.520.7102.718.699.419.8105.624.2102.527.00.65 − 6.0 − 2.8 − 4.0 − 3.1Leucine172.134.1166.428.0176.435.5164.557.3181.340.3175.044.50.30 − 5.2 − 2.7 − 5.0 − 6.2Lysine219.377.1195.943.0218.349.4211.349.5229.286.4209.044.90.37 − 4.4 − 4.9 − 6.51.1Methionine31.07.530.09.930.45.930.56.931.77.731.39.60.57 − 2.4 − 4.5 − 4.2 − 2.6Ornithine69.416.561.821.168.412.563.721.270.219.061.529.60.83 − 1.2 − 2.60.53.5Phenylalanine71.610.969.313.871.610.573.413.675.513.274.518.50.47 − 5.4 − 5.3 − 7.2 − 1.5Proline201.055.1203.949.5198.255.9199.350.5203.777.5181.435.90.65 − 1.3 − 2.711.79.4Serine104.626.397.638.9110.321.1112.028.7105.526.3101.729.20.30 − 0.84.5 − 4.19.6Threonine138.535.5133.559.2139.528.4133.328.0136.140.0131.145.10.831.82.51.81.7Total AA3127.4555.23141.5695.13162.9440.93189.4697.13162.8636.63087.1511.50.47 − 1.10.01.73.3Tryptophan66.612.661.920.263.711.262.011.366.018.959.98.70.830.9 − 3.53.23.3Tyrosine80.824.573.527.378.113.875.619.084.233.779.724.50.57 − 4.1 − 7.5 − 8.1 − 5.4Valine278.946.8273.437.5288.269.4265.877.6291.266.7279.848.90.78 − 4.3 − 1.0 − 2.3 − 5.1POST* postbiotic, PRO probiotic, PLA placeboDenotes significanceTable 4Time of maximal observed concentration (Tmax)Amino acidPostbioticProbioticPlacebop* (Friedman test)Median percent differencesMedianIQRMedianIQRMedianIQRPOST vs. PROPRO vs. PLAAlanine120606067.5451200.1490.928.6Arginine1206012060120600.890.00.0Asparagine12060606060600.3766.70.0Aspartic Acid15015045906067.50.4185.7 − 28.6BCAA1203012060120150.580.00.0Citrulline4597.56052.530600.2140.066.7Cystine0750600300.38NANAEAA1207512060120600.700.00.0Glutamic Acid6010560904597.50.5028.628.6Glutamine9067.56097.56097.50.4940.00.0Glycine12075609060900.1966.70.0Histidine12060606060900.2366.70.0Isoleucine1206012015120600.030.00.0Leucine1207512015120600.960.00.0Lysine1207512060120600.130.00.0Methionine6097.5607530600.0466.766.7Ornithine1206012060120150.030.00.0Phenylalanine12060906090600.0828.60.0Proline9060606060450.0640.00.0Serine1201206067.59067.50.4128.6 − 40.0Threonine12075906060600.0966.740.0Total AA120309060120600.040.0 − 28.6Tryptophan6097.56037.530750.2566.766.7Tyrosine120601206090600.6328.628.6Valine120159060120150.480.0 − 28.6POST postbiotic, PRO probiotic, PLA placebo*Denotes significance
Safety Analysis
All safety blood values for CBC and CMP were within normal limits for all participants throughout the study duration. Safety outcomes are displayed in the supplementary material.
Discussion
The increasing worldwide interest in adopting a more plant-based, whole-food diet is often met with concerns about whether these diets allow for the ingestion of adequate protein, both in terms of quantity and quality. A lifestyle implementing a balanced plant-based and protein-rich diet can offer health benefits for individuals [30]; however, the distribution of amino acids within plant protein versus animal protein differs and may subsequently impact the gut—even when protein consumption meets the recommended dietary allowance (RDA) [31]. Probiotics as a whole have demonstrated the ability to aid human physiology in multiple ways including the improvement of gut health [11], mood [32], stress and depression [33], immune system function [34], and allergy response [35]. Various strains of probiotics have been observed to improve the absorption of nutrients, namely, protein, through their influence in upregulating digestive enzymes, which has led some researchers to believe that probiotics may also positively influence body composition by facilitating muscle protein synthesis [20, 36, 37].
Postbiotics are a more recent conception and are thought to positively influence health as effectively as probiotics with differences only in their preparation, though research is warranted to fully elucidate the mechanisms by which postbiotics affect the gut [8]. Additionally, the potential benefits of postbiotic supplementation could extend beyond just the host. For example, postbiotics are not as sensitive as their “live” counterparts and are therefore less susceptible to degradation with changes in the environment. This characteristic may allow for a longer shelf-life and facilitate transport and storage without deterioration. Postbiotics may also possess the ability to better withstand the harsh environment of the digestive tract [8].
In a previous study, we demonstrated that the ingestion of plant protein with 2 weeks of probiotic supplementation containing L. paracasei DG and L. paracasei LPC-S01 increased circulating levels of amino acids when compared to a placebo. Thus, the purpose of the present study was to examine whether supplementation of a multi-strain probiotic consisting of 5 billion CFU L. paracasei DG plus 5 billion CFU L. paracasei LPC-S01 or a postbiotic consisting of the same strains would alter the absorption of individual and total amino acids following ingestion of a plant-based meal. The primary findings of the present study were observed in the postbiotic supplementation group whereby the observed increase in circulating levels of alanine, asparagine, citrulline, cystine, glycine, methionine, proline, and total amino acids demonstrates: (1) confirmation that the administration of the strains DG and LPC-S01 may aid in protein absorption and (2) that postbiotic supplementation L. paracasei DG and LPC-S01 may potentially be more effective than probiotic supplementation. Moreover, incremental area under the curve (iAUC) values were higher for arginine in the postbiotic condition when compared to the placebo condition. Additionally, time to concentration max (Tmax) in the postbiotic condition occurred later than in both the placebo and probiotic conditions for isoleucine, methionine, ornithine, and total amino acids. This may reflect additional unique effects of postbiotics on intestinal physiology.
Prior to the present investigation, there were, to our knowledge, no studies that examined postbiotic supplementation and its co-ingestion of a plant protein–based meal. That said, the current body of knowledge regarding postbiotic supplementation and its positive impact on health cannot be ignored. Postbiotics have been reported in animal models to modulate the gut environment leading to subsequent benefits similar to those obtained through probiotic supplementation [38–40]. Investigations analyzing postbiotics in humans have reached similar conclusions relative to the efficacy of postbiotics. Lee and colleagues [41] assessed probiotic versus postbiotic supplementation compared to a placebo following 6 weeks of treatment and measured biomarkers indicative of muscle damage. They observed that the postbiotic was just as effective in reducing muscle damage as probiotics when compared to the placebo. In fact, there are several studies that observe the positive influence of postbiotics whether it is immune function, recovery, inflammation, and now, plant protein absorption [41–44].
In our study, we selected a postbiotic prepared through γ-irradiation over the more commonly used heat inactivation. Recent studies indicate that gamma irradiation removes lactobacilli’s ability to replicate while preserving their metabolic activity [45]. Similarly, in our study, tests on the industrial biomass of L. paracasei LPC-S01 demonstrated that γ-irradiation not only effectively inactivates cells’ reproductive capacity but also preserves both membrane integrity, potentially leading to better retention of the probiotic properties of the original live microorganism. The preservation of membrane integrity is essential for bacterial cells because it maintains the proper internal environment for cellular functions, including enzyme activity. If the membrane is compromised, it can disrupt enzyme function.
Unexpectedly, we observed that irradiated bacterial cells were even more effective than live cells in facilitating the release of free amino acids. This surprising and apparently counterintuitive result could have several speculative explanations. One possibility is related to L. paracasei’s auxotrophy for various amino acids, which, as in many lactobacilli, is due to evolutionary adaptation to nutrient-rich substrates like milk or animal intestines, leading to efficient systems for the import and use of exogenous amino acids [46–48]. Unlike live bacteria, irradiated cells are unable to reproduce, likely resulting in reduced amino acid import and utilization, which in turn may leave a greater pool of free amino acids available for host absorption. It is also possible that γ-inactivation, despite preserving cytoplasmic membranes, induces the release of cellular components such as proteolytic enzymes, which could further promote protein hydrolysis. Finally, we cannot rule out that the postbiotic and the probiotic preparations may influence differently the host’s microbiota, immune responses, or gene expression in the small intestine mucosa, thereby impacting amino acid absorption. These hypotheses, however, remain speculative and require further investigation. Overall, the present study adds to the mounting evidence in support of postbiotic supplementation and offers the first observation of improved amino acid absorption following the co-ingestion of a postbiotic supplement versus a probiotic supplement with a plant protein–based, mixed meal.
Interestingly, in this trial, the probiotic condition exhibited increases in only cystine (+ 9%) when compared to the placebo. This finding differs from our previous experiment in which the appearance of amino acids following the co-ingestion of a probiotic supplement and plant protein was greater in multiple amino acids, including total amino acids, when compared to placebo [20]. Specifically, increases in leucine (+ 23.3%), isoleucine (+ 26.0%), valine (+ 21.5%), and total EAA (+ 16.0%) were observed in the probiotic condition as compared to the placebo condition [20]. A possible explanation for these differences could be due to the selected protein source and the amino acid composition of those sources. Despite protein sources from both studies containing similar amounts of pea protein (20 g), the vegan patty selected in the present investigation included an additional 14 g of fat and 7 g of carbohydrate. The mixed-meal aspect of this investigation may have altered absorption times in comparison to the protein shake absorption kinetics which was seemingly unaffected in the absence of carbohydrates and fat [20]. Yoshii and colleagues [49] observed that co-ingestion of leucine with a mixed meal containing fat, protein, and carbohydrates suppressed the appearance of leucine in plasma. Unfortunately, this was the only amino acid the investigators measured so comparisons to other individual amino acids, other BCAAs, EAAs, and total amino acids cannot be extrapolated.
Future research into postbiotic influence on nutrient absorption should continue and efforts to identify their mechanisms of action are necessary. Although probiotics can influence absorption through several different mechanisms, it is purported that increased digestive enzyme production is one of the primary drivers of amino acid absorption. The postbiotic used in this investigation was produced with membrane integrity intact and maintained enzyme activity; therefore, we speculate that amino acid absorption improved through increased digestive enzyme activity elicited by the postbiotic. This investigation presents promising and novel evidence that postbiotic supplementation enhances amino acid absorption in a plant-based, mixed meal. Additional research into varying forms of protein and amino acid profiles should also be conducted to fully understand the influence of postbiotics. For example, examination of the appearance of amino acids following the co-ingestion of a postbiotic with a whole food source of protein (plant versus animal), protein powder (plant versus whey versus soy), and mixed meals (animal versus plant-based) would aid in deepening our understanding. Finally, of interest to athletes and other individuals who choose to adopt a plant-based diet is the absorption of BCAAs and EAAs to optimize muscle repair and muscle protein synthesis. Evidence already exists demonstrating the ability of postbiotics to influence recovery; therefore, additional research into specific populations (e.g., vegans or vegetarians) should be conducted to determine if postbiotic supplements can similarly influence muscle growth and recovery.
A limitation of this research was the inability to analyze the intestinal microbiome to determine whether the interventions affected the abundance of microbial taxa or their metabolites. Although a follow-up study in women is expected, we recognize that the inclusion of only men is a limitation.
Conclusions
Two weeks of supplementation of postbiotic supplementation containing 5 billion AFU L. paracasei DG plus 5 billion AFU L. paracasei LPC-S01 resulted in significant improvements in amino acid absorption profiles for various amino acids and total amino acids compared to placebo and probiotic supplementation. This is the first data to report an improved absorption of amino acids in a mixed macronutrient meal and provides a rationale for probiotic and/or postbiotic supplementation as a support strategy to improve the amino acid response in the post-prandial state.
Supplementary Information
Below is the link to the electronic supplementary material.Supplementary file1 (PDF 457 kb)Supplementary file2 (XLSX 14 kb)Supplementary file3 (DOCX 50 kb)Supplementary file4 (DOCX 50 kb)Supplementary file5 (XLSX 17 kb)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Vinderola G, Sanders ME, Salminen S (2022) The concept of postbiotics. Foods 11(8). 10.3390/foods 1108107710.3390/foods 11081077 PMC 902742335454664 · doi ↗ · pubmed ↗
- 2Ferrari L, Panaite SA, Bertazzo A, Visioli F (2022) Animal- and plant-based protein sources: a scoping review of human health outcomes and environmental impact. Nutrients 14(23). 10.3390/nu 1423511510.3390/nu 14235115 PMC 974133436501146 · doi ↗ · pubmed ↗
- 3Alcorta A, Porta A, Tárrega A, Alvarez MD, Vaquero MP (2021) Foods for plant-based diets: challenges and innovations. Foods 10(2). 10.3390/foods 1002029310.3390/foods 10020293 PMC 791282633535684 · doi ↗ · pubmed ↗
- 4Mariotti F, Gardner CD (2019) Dietary protein and amino acids in vegetarian diets-a review. Nutrients 11(11). 10.3390/nu 1111266110.3390/nu 11112661 PMC 689353431690027 · doi ↗ · pubmed ↗
- 5Mazziotta C, Tognon M, Martini F, Torreggiani E, Rotondo JC (2023) Probiotics mechanism of action on immune cells and beneficial effects on human health. Cells 12(1). 10.3390/cells 1201018410.3390/cells 12010184 PMC 981892536611977 · doi ↗ · pubmed ↗
- 6Lee MC, Ho CS, Hsu YJ, Huang CC (2022) Live and heat-killed probiotic Lactobacillus paracasei PS 23 accelerated the improvement and recovery of strength and damage biomarkers after exercise-induced muscle damage. Nutrients 14(21). 10.3390/nu 1421456310.3390/nu 14214563 PMC 965858736364825 · doi ↗ · pubmed ↗
- 7Lee CC, Liao YC, Lee MC, Cheng YC, Chiou SY, Lin JS et al. (2022) Different impacts of heat-killed and viable Lactiplantibacillus plantarum TWK 10 on exercise performance, fatigue, body composition, and gut microbiota in humans. Microorganisms 10(11). 10.3390/microorganisms 1011218110.3390/microorganisms 10112181 PMC 969250836363775 · doi ↗ · pubmed ↗
- 8Kato K, Arai S, Sato S, Iwabuchi N, Takara T, Tanaka M (2024) Effects of heat-killed Lacticaseibacillus paracasei MCC 1849 on immune parameters in healthy adults-a randomized, double-blind, placebo-controlled, parallel-group study. Nutrients 16(2). 10.3390/nu 1602021610.3390/nu 16020216 PMC 1082148738257109 · doi ↗ · pubmed ↗