Validation of phiC31-mediated expression and functional knockout of Opn3 in the Opn3-phiC31o knock-in mouse
Rachel Kam Yan Kwok, Hikaru Ikuta, Chisato Iba, Yuka Nakano, Ziyan Ma, Yahan Chuai, Yuichi Hiraoka, Taichi Sayanagi, Toshihide Kurihara, Satoru Moritoh, Kenji F. Tanaka

TL;DR
Researchers created a mouse model to study Opn3 function by inserting a gene recombination system, confirming Opn3 knockout and its effects on body temperature and vision.
Contribution
A novel Opn3-phiC31o knock-in mouse model was developed for targeted gene recombination and functional knockout of Opn3.
Findings
Opn3-phiC31o mice showed a larger decrease in body temperature under cold exposure compared to wild-type controls.
The Opn3-phiC31o model confirmed functional knockout of Opn3 without disrupting Chml expression in homozygous mice.
Recombination efficiency in the cerebellum of double transgenic mice was 30% to 44%.
Abstract
Opn3 is a non-visual blue light-sensitive opsin that has recently been reported to have an expansive repertoire of biological functions. To investigate the function of Opn3-expressing cells, we aimed to generate a system in which Opn3-expressing cells can be targeted by site-specific gene recombination. Opn3-phiC31o knock-in (KI) mice were generated using the CRISPR-Cas9 method. The phiC31o-poly(A) cassette was inserted into the translation start site in exon 1. Opn3 mRNA and phiC31o mRNA were visualized by in situ hybridization (ISH). 5' rapid amplification of cDNA end (5' RACE) analysis was performed using RNAs from wild-type mouse cerebral cortex and cerebellum to identify the transcription start site of Chml, predicted to be shared with the transcription start site of Opn3. Cold-induced decrease in body temperature was monitored with a telemetric probe to confirm the phenotype of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
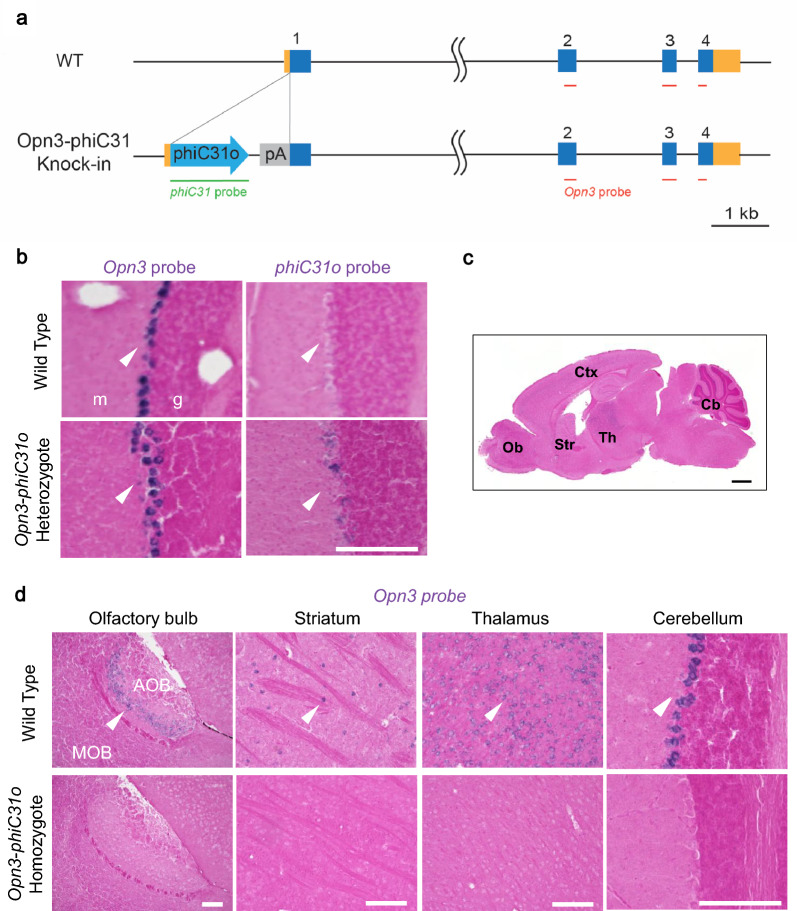 Figure 1
Figure 1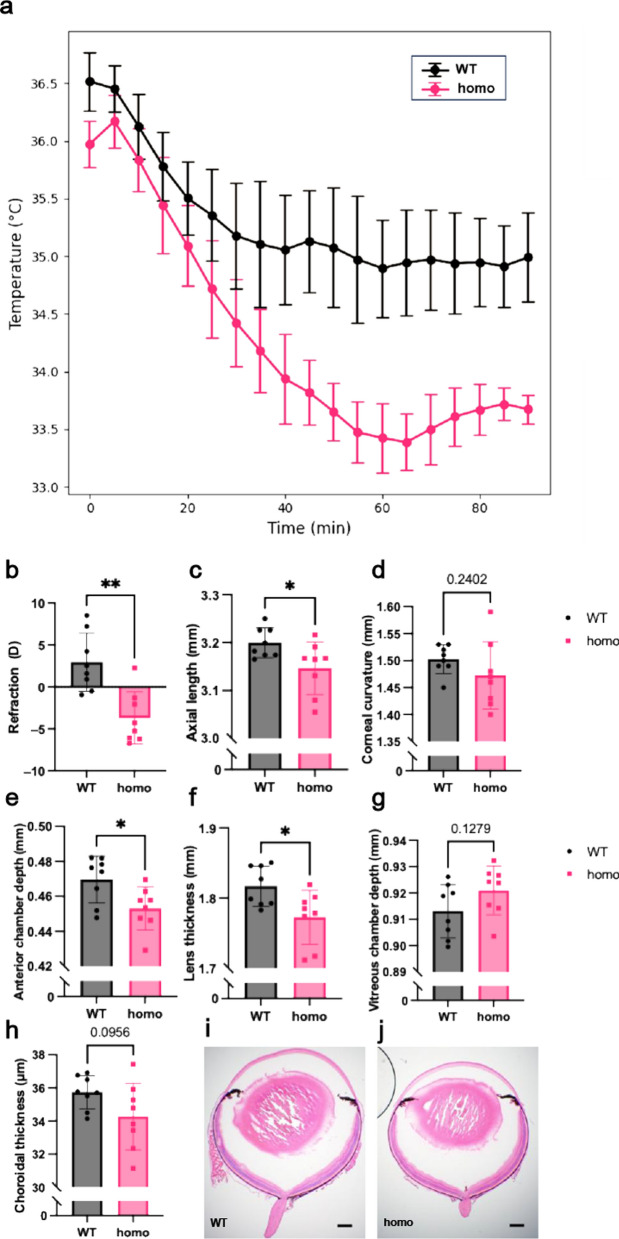 Figure 2
Figure 2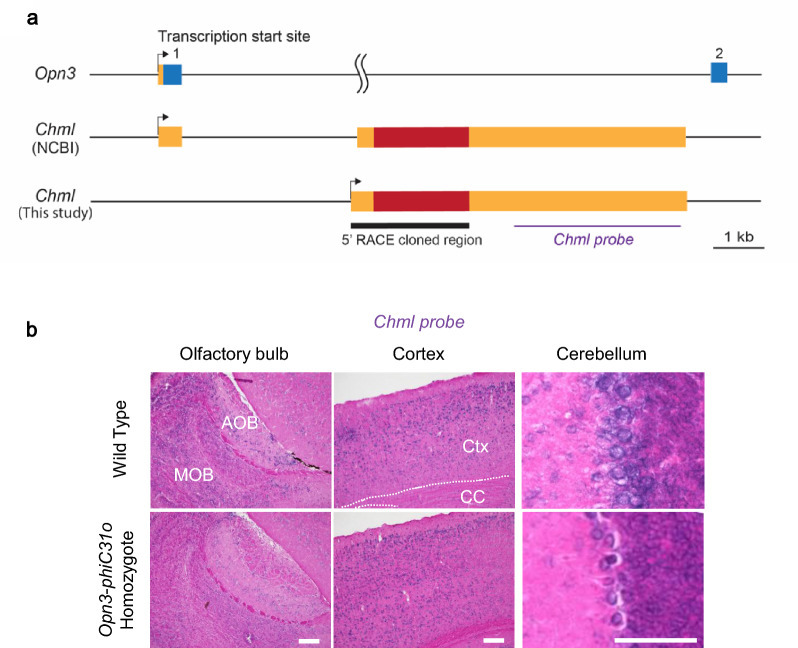 Figure 3
Figure 3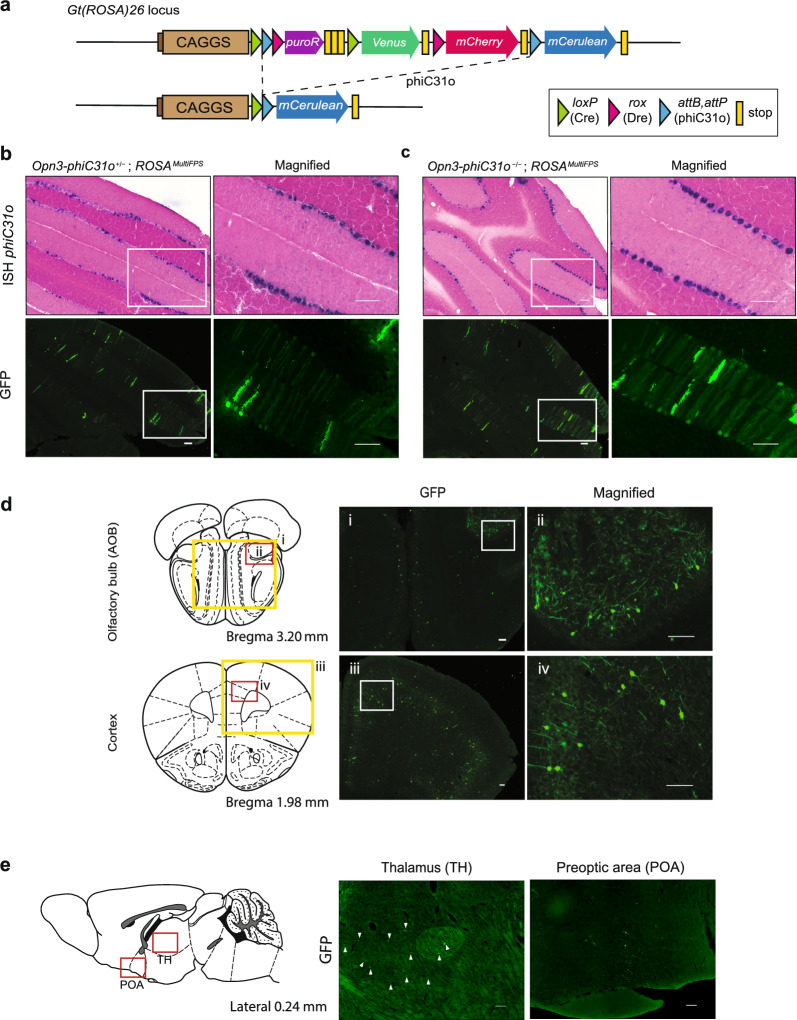 Figure 4
Figure 4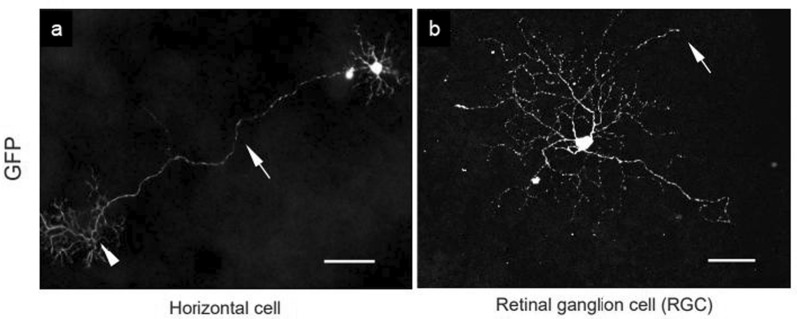 Figure 5
Figure 5- —AMED-CREST
- —JSPS KAKENHI
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhotoreceptor and optogenetics research · Circadian rhythm and melatonin · Light effects on plants
Background
Opn3 is a non-canonical opsin [1] expressed outside the eye and was the first mammalian extraocular opsin to be discovered [2]. Initially identified in the mouse brain, it was originally named encephalopsin [2], and later renamed panopsin after the discovery of its widespread expression in peripheral organs [3]. Opn3 expression was first identified by in situ hybridization (ISH) in the cerebral cortex, striatum, preoptic areas, lateral thalamus, paraventricular nucleus of the hypothalamus, spinal cord, and cerebellar Purkinje cells [2]. Protein expression of Opn3 has also been reported in some parts of the adult mouse brain and retina [4]. The expression of Opn3 in the adult brain and developing nervous system was later examined using Opn3-mCherry (Opn3-mCh) mice [5] and an Opn3 promoter-driven eGFP (Opn3-eGFP) bacterial artificial chromosome (BAC) transgenic mouse line [6], respectively.
Opn3 is a type of blue light-sensitive G-protein-coupled receptor. Molecular and biochemical properties, such as light sensitivity, retinal binding, G-protein activation, intracellular responses, and absorption spectra, of several Opn3 homologs have been studied [7–11]. Recently, one of the Opn3 homologs, MosOpn3, has been used for optogenetic manipulation in mammals [12], Caenorhabditis elegans [13], and zebrafish [14].
Opn3 has been reported to perform a variety of functions, including influencing the intrinsic retinal rhythmic amplitude [15], melanin production in human primary melanocytes [16, 17] and melanocyte survival [18], photorelaxation of smooth muscle in the airway [19] and colon [20], hair growth in the human hair follicle [21], expression in the iris/ciliary body complex [22], and early stages of eye development [23]. In addition, Opn3 is required for the restoration of the barrier function [24], regulation of cancer cell functions [25, 26], and is sensitive to osmotic stimuli and light [27]. Opn3 knockout mice exhibit a lower body temperature than wild-type mice when challenged with cold conditions [28]. This is because Opn3 is required for blue light enhanced activation of the lipolysis pathway. Other reported phenotypes of Opn3 knockout include refractive myopia [29], attenuated acoustic startle reflex [30], and susceptibility to diet-induced obesity [31].
Site-specific recombinases (SSRs), such as Cre, are powerful tools for analyzing gene function and tracing the specific progeny of cells that once expressed a gene of interest in vivo [32]. Cre/loxP is the most widely used system for genome manipulation in mice. Recently, several additional SSR systems have been developed, such as Flp/FRT, Dre/rox, and phiC31/att [33, 34]. phiC31 integrases mediate unidirectional recombination between attB and attP sites and produce the resulting recombination sites, attL, and attR [35]. Using transgenic and reporter mice, phiC31 can induce recombination by deleting a stop cassette flanked by attB and attP [36].
The combination of these SSR systems enables simultaneous gene targeting at different gene loci. For a deeper understanding of non-visual opsins in mice, the combination of SSR systems in each opsin gene would be beneficial. While Cre driver lines of Opn3, Opn4, and Opn5 exist [28, 37–39], drivers using SSRs other than Cre have not been developed. The aim of this study was to develop Opn3-phiC31o knock-in (KI) mice using CRISPR-Cas9 gene editing technology and to validate phiC31 integrase-mediated recombination in heterozygous mice and knockout phenotype in homozygous mice.
Methods
Animals
All animal procedures in this study were performed in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals and were approved by the Animal Research Committee of Keio University School of Medicine (protocol number: A2023-023). *ROSA26 *^MultiFPsΔPuro^ mice [40, 41] were provided by the RIKEN BioResource Research Center (ID: RBRC10966). Both male and female mice were used, as it has been reported that there are no significant differences in Opn3 expression between the sexes [5]. All mice were maintained on a 12/12 h light/dark cycle.
Generation of Opn3-phiC31o KI mice
Opn3-phiC31o KI mice were generated using the previously described cloning-free CRISPR-Cas9 system [42]. CRISPR RNA (crRNA) was designed to introduce the double-strand break 7 bp upstream of the endogenous translation start site of Opn3. Both crRNA (5'-CCGAGUACAUGGCCCGACGCguuuuagagcuaugcuguuuug-3') and trans-activating CRISPR RNA (tracrRNA, 5'-AAACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCU-3') were chemically synthesized by the manufacturer (Fasmac, Kanagawa, Japan). KI targeting vector with 1.5 kb each of 5' arm and 3' arms as homologous regions and phiC31o (codon optimized phiC31)-bovine growth hormone poly(A) sequence was synthesized by Twist Bioscience (South San Francisco, CA, USA). All CRISPR-Cas9 components, including the Cas9 protein (New England Biolabs, Ipswich, MA, USA), cr/tracr RNA, and KI targeting vector, were injected into C57BL/6J (CLEA Japan, Inc., Shizuoka, Japan) fertilized egg at the pronuclei stage. Mice were screened by PCR-based genotyping followed by Sanger sequencing. Founder generation mice were backcrossed to wild-type, and the progenies were used in the experiments.
To detect phiC31o-mediated recombination in adult brain and retina, Opn3-phiC31o KI mice were crossed with ROSA26 ^MultiFPsΔPuro^ reporter mice [40]. This reporter strain carries an attB-multiple reporter-attP cassette between the CAG promoter and the mCerulean gene. In this study, Opn3-phiC31o^+/+^ (homozygous); ROSA26 ^MultiFPs^ mice and Opn3-phiC31o^+/−^ (heterozygous); ROSA26^MultiFPs^ mice were used to analyze the phiC31o-mediated recombination.
Genotyping
The following PCR primer sets were used for mouse genotyping: Opn3 IDA F v2 (5'-AGAGGTTCTAAGAAGGCGGA-3') and Opn3 IDA R (5'-GGTGACTCCGAACAGGGATA-3') for the detection of the wild-type allele (742 bp) and the KI allele (2841 bp) using Tks Gflex DNA Polymerase (TAKARA Bio, Shiga, Japan). In some cases, the Opn3 IDA F v2 and Opn3 IDA R primers were used to identify the wild-type allele (742 bp), while the phiC31-F (5'-GAAGCACTTCAGAAAGCAGCAG-3') and Opn3 IDA R primers were used to detect the KI allele (916 bp). These procedures were performed using the KAPA Taq Extra PCR Kit (Kapa Biosystems, Inc., Wilmington, MA, USA).
Tissue collection
Mice were anesthetized intraperitoneally with a mixture of ketamine (100 mg/kg)-xylazine (10 mg/kg), and then transcardially perfused with 4% paraformaldehyde (PFA)/phosphate buffer solution. The brains were collected and postfixed with 4% PFA overnight at room temperature (RT). After postfixation, brains were cryoprotected in 20% sucrose/phosphate-buffered saline (PBS) until they sank to the bottom of the conical tubes. Brains were snap frozen for approximately 10 s in 2-methylbutane precooled with liquid nitrogen. Coronal sections were cut at 25 µm on a cryostat (Leica CM3050 S, Leica Biosystems, Nussloch, Germany) and mounted on silane-coated glass slides (Matsunami Glass Ind., Ltd., Osaka, Japan). Mouse eyes were also collected after perfusion and postfixed with 4% PFA for 2 h at RT. The eyes were then washed 2–3 times with PBS and the retina was removed from the eye cup for immunohistochemical staining.
Immunohistochemistry
Brain sections and retinal flatmounts were incubated overnight at RT in a goat polyclonal anti-GFP antibody (1:250, Rockland Immunochemicals Inc., Pottstown, PA, USA). They were then washed and incubated for 2 h at RT in an anti-goat secondary antibody conjugated to Alexa Fluor 488 (1:1000 dilution; Invitrogen, Carlsbad, CA, USA) or Alexa Fluor 555 (1:1000 dilution, Invitrogen). Cerulean is a derivative of GFP and can be detected by the anti-GFP antibody. Fluorescence images were captured using an inverted microscope (BZ-X710, Keyence, Kyoto, Japan) or a confocal microscope (LSM 710, Carl Zeiss AG, BW, Germany). The maximum intensity projection image of a confocal z-stack was obtained using Fiji/ImageJ (National Institutes of Health, Bethesda, MD, USA).
ISH
ISH was performed as described previously [43]. Briefly, digoxigenin-labeled phiC31o, Opn3, and Chml antisense RNA probes were hybridized to sections, NBT/BCIP (Roche, Basel, Switzerland) was used for color development, and nuclear fast red (Sigma Aldrich, St. Louis, MO, USA) was used for counterstaining. To generate the Opn3 probe, a plasmid with the same sequence as the Opn3 probe used in the Allen Brain Atlas (Opn3-RP 050331_04_B02) was synthesized at GenScript Biotech (Piscataway, NJ, USA). To make the phiC31o probe, we cloned the phiC31o coding sequence (1815 bp from pPGKPhiC31obpA (addgene plasmid number 13795) into 5' BamHI and 3' EcoRI of pBlueScript II SK ( −) (Agilent Technologies, Inc., Santa Clara, CA, USA). The sense and antisense probe for Opn3 cover nt 632–1286 (NM_010098; 1763 bp; coding region is 97–1299 bp). The sense and antisense probe for Chml probe cover nt 3552–6316 (NM_021350; 6907 bp; coding region is 791–2656 bp).
Calculation of the recombination efficiency in mice
Multiple areas of the fluorescence images of the cerebellum in which mCerulean expressions were amplified with GFP antibody were selected, and the number of somas with mCerulean expression in the Purkinje cell layer was counted using the cell counter in Fiji/ImageJ (National Institutes of Health). The same regions of the cerebellum in consecutive sections that had undergone ISH with phiC31o probe visualization were counted using the same method. The average of the counted areas was taken, and the recombination efficiency in mice was calculated using the following equation:
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\frac{\text{number of mCerulean }-\text{expressing soma}}{\text{number of }phiC31o-\text{expressing cells}}\times 100\%$$\end{document}Measurement of body temperature using telemetric probes
Telemetric probes (Anipill, BodyCAP, Hérouville Saint-Clair, France) were surgically implanted intraperitoneally in mice under ketamine/xylazine anesthesia. Mice were allowed to recover for at least 5 days after surgery. Mice were then placed in a 4 °C chamber (BR-33FL, TAITEC, Saitama, Japan) and body temperature was recorded every 5 min for 90 min. Male and female mice aged 2- to 7-month-old were used. Food and water were provided ad libitum during the cold treatments.
Measurement of ocular biometric parameters
Prior to measurements, 0.5% tropicamide and 0.5% phenylephrine eye drops (Santen Pharmaceutical Co., Ltd., Osaka, Japan) were administered to induce mydriasis. Then, mice were anaesthetized via intraperitoneal injection of a mixture containing midazolam, medetomidine, and butorphanol tartrate (0.01 mL/g bodyweight).
Refractive status was assessed using an infrared photorefractor (Steinbeis Transfer Center, Graz, Austria). Axial length was measured with a spectral-domain optical coherence tomography (SD-OCT) system (Envisu R4310; Leica, Wetzlar, Germany) and was defined as the distance between the corneal vertex and the retinal pigment epithelium layer near the optic nerve head. Anterior chamber depth, lens thickness, and vitreous chamber depth were calculated based on the proportional lengths of each region measured on OCT scans using ImageJ (NIH), normalized to the total axial length. For choroidal analysis, an OCT image was acquired from a retinochoroidal region located 500 µm from the optic nerve head. The area between the retinal pigment epithelium and the posterior choroidal boundary was quantified using ImageJ and the choroidal thickness (ChT) was calculated as the ratio of this area to the corresponding boundary length, as previously described [44]. Corneal radius of curvature was measured using a separate infrared photorefractor (Steinbeis Transfer Center, Germany) according to a previously described method [45]. After general anesthesia and mydriasis, eight infrared LEDs arranged in a circular pattern illuminated the mouse cornea, and reflected light was captured by an infrared camera. The diameter of the reflected light ring on the corneal surface was used to determine corneal curvature.
5' Rapid amplification of cDNA end (5' RACE) analysis
Experiments were performed using the GeneRacer kit according to the manufacturer's instructions (ThermoFisher Scientific, Waltham, MA, USA). Total RNA was isolated from the cerebral cortex and cerebellum of a 3-month-old male C57BL/6J mouse using Ambion TRIzol Reagent (Life Technologies, Carlsbad, CA, USA). Five micrograms of total RNA were reverse transcribed using the GeneRacer kit with SuperScript III RT and GeneRacer Oligo dT Primer. Amplification of 5' cDNA ends was performed using Tks Gflex DNA Polymerase (TAKARA Bio) with the GeneRacer 5' Primer and a Chml gene-specific primer (5'-TGCTTCACCACAGGTTCCAAGTCTTC-3') in the first round of PCR. Nested PCR was performed using the GeneRacer 5' Nested Primer and a Chml gene-specific nested primer (5'-CCATGACCCGAACAGAAGTGGTTCC-3'). Gel electrophoresis of the 5' nested PCR products yielded a single band of approximately 2 kb size in both the cerebral cortex and cerebellum. These PCR fragments were cloned using the Zero Blunt TOPO PCR Cloning Kit for Sequencing (Invitrogen) and sequenced (Eurofins Genomics, Tokyo, Japan). The cloned sequence was submitted to the EMBL/GenBank/DDBJ database (accession number LC816633).
Statistical analysis
Statistical analyses were performed using Python 3.9. The graph showing the decrease in body temperature in the cold exposure experiment was analyzed by one-way repeated measures ANOVA. The level of significance was defined as a P value of less than 0.05. Data are presented as mean ± standard error.
Results
Generation and validation of Opn3-phiC31o KI mice
We generated KI mice in which phiC31o-poly(A) was inserted into the translation start site of Opn3 using CRISPR-Cas9 genome editing (Fig. 1a). To confirm whether phiC31o is expressed under the control of the Opn3 promoter, and whether the phiC31o mRNA expression pattern reflects the endogenous Opn3 mRNA expression, ISH with Opn3 and phiC31o probes was performed in serial sections of Opn3-phiC31o heterozygous mouse cerebellum. The cerebellum was selected for comparison due to its well established high Opn3 expression in the brain [5]. In two consecutive brain sections, endogenous Opn3 and exogenous phiC31o signals were similarly localized in the Purkinje cell layer of the cerebellum (Fig. 1b), with phiC31o reflecting approximately 97% of Opn3 expression. This suggests that the expression of phiC31o in the Opn3-phiC31o mouse line corresponds to the endogenous Opn3 expression. Opn3 signal levels were not observed in Opn3-phiC31o homozygous mouse samples (Fig. 1c, 1d), indicating the successful knockout of Opn3.Fig. 1. Generation of Opn3-phiC31o KI mice using CRISPR-Cas9. a Schematic representation of the wild-type allele of the Opn3 gene according to the NCBI genome annotation (NM_010098.4) and the KI allele, Opn3-phiC31o. b Validation of Opn3-phiC31o expression using the heterozygous Opn3-phiC31o mouse. ISH results for Opn3 mRNA and phiC31o mRNA are shown in representative coronal sections of the cerebellum of the heterozygous Opn3-phiC31o KI mouse. g, granule layer; m, molecular layer. Scale bar: 150 μm. c A schematic image of the sagittal brain highlighting regions examined in this paper. Ob, olfactory bulb; Ctx, cortex; Str, striatum; Th, thalamus; Cb, cerebellum. Scale bar: 1 mm. d Validation of Opn3 knockout in homozygous Opn3-phiC31o mice. Representative sagittal sections of the olfactory bulb, striatum, thalamus and cerebellum from wild-type and homozygous mice were subjected to ISH using the Opn3 probe. Opn3 mRNA expression in the Opn3-phiC31o homozygous mouse was then compared with that in the wild-type mouse. AOB, accessory olfactory bulb; MOB, main olfactory bulb. Scale bars: 150 µm
Opn3 was functionally knocked out in our Opn3-phiC31o mice
Previous studies have reported that Opn3 knockout mice exhibit a significantly larger decrease in body temperature than wild-type mice in response to cold exposure [28]. We recreated these cold exposure experiments to functionally validate the Opn3-phiC31o allele. Opn3-phiC31o homozygous mice (n = 5) and wild-type mice (n = 5) were both surgically implanted with a telemetric probe under general anesthesia and subsequently placed in a 4 °C chamber. The body temperatures of both groups were recorded every 5 min over a 90 min period. Analysis using one-way repeated measures ANOVA revealed that the body temperature of the Opn3 knockout mice was significantly lower than that of the wild-type mice (P = 0.044, Fig. 2a). This result is consistent with previous findings.Fig. 2Opn3 knockout functional experiment in homozygous Opn3-phiC31o mice. a Decrease in body temperatures of wild-type (n = 5) and Opn3 knockout mice (n = 5) after cold exposure (P = 0.044; One-way repeated measures ANOVA). Body temperatures were recorded every 5 min for 90 min. Data is presented as mean ± standard error. WT, wild-type mice; homo, Opn3-phiC31o homozygous mice. b Refraction; c Axial length; d Corneal radius of curvature; e Anterior chamber depth; f Lens thickness; g Vitreous chamber depth; h Choroidal thickness. All parameters were measured in wild-type littermates (n = 8) and homozygous Opn3-phiC31o mice (n = 8). Data are presented as mean ± standard deviation. Statistical comparisons were performed using a two-tailed unpaired t-test with Welch’s correction. *P < 0.05; **P < 0.01. i Representative frozen sections of wild-type eyeballs (H&E staining). Scale bar: 200 µm. j Representative frozen sections of homo eyeballs (H&E staining). Scale bar: 200 µm
To further confirm the functional knockout of Opn3, we assessed refractive measurements and ocular biometry in Opn3-phiC31o homozygous mice. Refractive assessments revealed that Opn3-phiC31o homozygous mice (n = 8) exhibited a myopic phenotype compared to wild-type controls (n = 8, Fig. 2b).
Ocular biometry was performed using a SD-OCT system. Axial length (Fig. 2c), anterior chamber depth (Fig. 2e), lens thickness (Fig. 2f), vitreous chamber depth (Fig. 2g), and choroidal thickness (Fig. 2h) was measured. In Opn3-phiC31o homozygous mice, axial length was significantly reduced (Fig. 2c), along with decreased anterior chamber depth (Fig. 2e) and lens thickness (Fig. 2f), compared to wild-type mice. No significant differences were observed in vitreous chamber depth (Fig. 2g) or choroidal thickness (Fig. 2h). In addition, corneal curvature did not differ significantly between groups (Fig. 2d). These results are consistent with previous reports that Opn3 knockout mice exhibit a myopia-like phenotype [29], and further support the successful functional knockout of Opn3 in our mouse model.
The Opn3-phiC31o homozygotes did not disrupt Chml gene transcription
Chml, also known as choroideremia-like, is a gene associated with diseases such as choroideremia [46]. According to the National Center for Biotechnology Information (NCBI) genome annotation, the first exon of Chml overlaps with the first exon of Opn3 (Fig. 3a**)**. Therefore, knocking in of phiC31o-poly(A) at the first exon of Opn3 is predicted to disrupt both Opn3 and Chml genes. However, ISH analysis of Chml expression in the olfactory bulb, cortex, and cerebellum in both wild-type and Opn3-phiC31o homozygous mice still showed Chml signals in the latter (Fig. 3b), demonstrating that Chml is not disrupted in the Opn3-phiC31o homozygous mouse. To determine the exact transcription start site of Chml, RNA was isolated from the cerebral cortex and cerebellum and analyzed by the 5' RACE technique (see Methods). Sequence analysis of 11 clones from the cerebral cortex and 6 clones from the cerebellum by 5' RACE showed that they all have identical sequences (EMBL/GenBank/DDBJ accession number LC816633) and that, at least in the cerebral cortex and cerebellum, the transcribed region of Chml did not initiate at the predicted first exon but starts transcription at 112 bp upstream of the predicted second exon (Fig. 3a), confirming that our mouse line is a single knockout model of Opn3.Fig. 3. The transcription start site of Chml was different from the NCBI genome annotation. a Schematic diagram of the Opn3 gene (NM_010098.4) and the Chml gene (NM_021350.3) according to the NCBI genome annotation, as well as the Chml gene structure cloned by 5' RACE analysis using RNA from the cerebral cortex and cerebellum. b Representative sagittal sections of the olfactory bulb, cortex, and cerebellum showing Chml expression in wild-type and Opn3-phiC31o homozygous mice. AOB, accessory olfactory bulb; MOB, main olfactory bulb; Ctx, cortex; CC, corpus callosum. Scale bars: 150 µm
Recombination efficiency of the Opn3-phiC31o mice is low
To detect phiC31o-mediated recombination in adult brains, we crossed Opn3-phiC31o KI mice with ROSA26 ^MultiFPsΔPuro^ reporter mice [40]. This reporter strain carries an attB-multiple reporter-attP cassette between the CAG promoter and the mCerulean gene (Fig. 4a)**. Upon recombination mediated by phiC31o, the mCerulean cyan fluorescent protein will be expressed. To amplify the mCerulean reporter signal, brain sections were immunostained with an anti-GFP antibody. We then evaluated the recombination efficiency of phiC31o using serial sections of phiC31o ISH probe and GFP antibody. By counting the phiC31o-positive signals and GFP-positive signals in the consecutive section, we estimated that the phiC31o-mediated recombination in the cerebellum of Opn3-phiC31o^+/−^; ROSA26 ^MultiFPs^ mice occurred at 30.3% ± 6.36% (n = 3), while that of Opn3-phiC31o^+/+^; ROSA26 ^MultiFPs^ mice was 44.4% ± 11.8% (n = 3). These data demonstrated that the expression of mCerulean reporter in the cerebellum of the mice heterozygous for the Opn3-phiC31o KI allele, Opn3-phiC31o^+/−^; ROSA26 ^MultiFPs^ (Fig. 4b), was lower than that in the cerebellum of the mice homozygous for Opn3-phiC31o KI allele, Opn3-phiC31o^+/+^; *ROSA26 *^MultiFPs^ (Fig. 4c).Fig. 4. Recombination efficiency and Opn3 expression in Opn3-phiC31o KI mice brain. a Schematic representation of the ROSA26 ^MultiFPs^ allele [40]. Multiple fluorescent proteins are inserted into the allele at the Gt(ROSA)26 locus under the control of the ubiquitous CAGGS promoter. mCerulean is activated by phiC31o-specific recombinase in response to the Opn3-phiC31o gene expression. b Representative coronal sections of the cerebellum for the calculation of recombination efficiency in the Opn3-phiC31o^+/−^; ROSA26 ^MultiFPs^ mouse. Magnified images of the cerebellum with visualization of phiC31o probe and GFP expression are shown. Scale bars: 100 μm. c Representative coronal sections of the cerebellum showing the recombination efficiency in Opn3-phiC31o^+/+^; ROSA26 ^MultiFPs^ mouse. Magnified images of the cerebellum with visualization of the phiC31o probe and GFP expression are shown. Scale bars: 100 μm. d Representative coronal sections of phiC31o-mediated mCerulean expression amplified with GFP antibody in the olfactory bulb (i) and the cerebral cortex (iii). Magnified images of the boxed regions are shown (ii, iv). Scale bars: 100 µm. e Representative sagittal sections of the brain of the Opn3-phiC31o^+/+^; *ROSA26 *^MultiFPs^ mouse showing phiC31o-mediated mCerulean expression amplified with GFP antibody in the thalamus (TH) (labelled with arrowheads) and preoptic area (POA). Scale bars: 100 µm
Opn3-phiC31-mediated mCerulean expression was examined in various regions of the mouse brain (Table 1). We observed mCerulean expression in the olfactory bulbs (Fig. 4d), cerebral cortex (Fig. 4d), Purkinje cell layer of the cerebellum (Fig. 4b, c), and slightly in the thalamus (Fig. 4e) and preoptic area (Fig. 4e). These reporter protein expression patterns are included in previously published reporter lines [5, 6]. Table 1phiC31-mediated gene expression in *Opn3-phiC31o; ROSA26 *^MultiFPs^ miceBrain regionExpression-positive structuresCortexACA, AI, AUD, MO, PTLp, RSP, ORB, SS, VISOlfactory areasAOB, MOBStriatumNot observedNucleus accumbensNot observedHippocampal formationNot observedThalamusSubregions not identifiedHypothalamusPOACerebellumDN, FN, IP, PurkinjeMidbrainNot observedPonsNot observedMedullaNot observedRetinaHC, RGCphiC31-mediated gene expressing CNS regions were shown in bold. The brain region list was created based on previous literature [5]. Cortex: ACA = anterior cingulate area; AI = agranular insular area; AUD = auditory area; MO = somatomotor area; PTLp = posterior parietal association area; RSP = retrosplenial area; ORB = orbital area; SS = somatosensory area; VIS = visual area; Olfactory areas: AOB = accessory olfactory bulb; MOB = main olfactory bulb. Hypothalamus: POA = preoptic area. Cerebellum: DN = dentate nucleus; FN = fastigial nucleus; IP = interposed nucleus. Retina: HC = horizontal cell; RGC = retinal ganglion cell
Low recombination efficiency in the retina
In addition to the phiC31o-mediated recombination in the brain, we examined recombination in the retina. Retinas were stained with GFP antibody to amplify the mCerulean signals and observed under a fluorescence microscope. However, recombination in the retina was inconsistent, showing extremely low or no occurrence. Among the 12 retinas (n = 6 mice) that we observed, only 3 of the retinas contained 5 mCerulean-expressing cells [1 horizontal cell (HC) from 1 retina (Fig. 5a) or 4 retinal ganglion cells (RGCs) from 2 retinas (Fig. 5b)].Fig. 5mCerulean-expressing cells in Opn3-phiC31o KI mice retina despite the low recombination efficiency. phiC31o-mediated mCerulean expression labelled with GFP antibody in the retina of Opn3-phiC31o^+/+^; *ROSA26 *^MultiFPs^ mice. A small number of (a) horizontal cells (HCs) and (b) retinal ganglion cells (RGCs) showed mCerulean expression. The HC and RGC were imaged by confocal and fluorescence microscope, respectively. a Arrow and arrowhead indicate an axon and axon terminal arborization of the HC, respectively. b Arrow indicates an axon of the RGC. Scale bars: 50 µm.
Discussion
Since the functional role of Opn3 is the least studied among the non-visual opsins [1], its expression in mammals has been extensively studied in recent years. It has been reported that Opn3 mRNA expression is found not only in the central nervous system (CNS), but also in peripheral tissues such as adipose tissue [28], skin [5], smooth muscles [19], and blood vessels [47]. Since there are currently no reliable antibodies for murine Opn3, several mouse reporter lines have been developed to assess Opn3 promoter activity, including Opn3-mCh KI [5] and Opn3-eGFP BAC transgenic mice [6]. To further investigate Opn3 expression in the CNS, we generated a new KI mouse model, Opn3-phiC31o.
Codon-optimization of the phiC31 gene into phiC31o has been reported to significantly improve recombination activity in embryonic stem (ES) cells, to a level similar to that of Cre recombinase [34]. However, the recombination efficiency of phiC31o was later reported to be lower than that of Cre- or Flpe-expressing transgenic mice [36]. For example, the phiC31o-mediated recombination efficiency of TH-PhiC31o mice crossed with the R26attR reporter has been reported to range from 4.7% to 48.2% in various brain regions [36]. The recombination efficiency of phiC31o was also observed to be relatively low compared with Cre and Dre in a recent study, which was around 76% [40]. Consistent with these findings, our Opn3-phiC31o^+/−^; *ROSA26 *^MultiFPs^ showed ~ 30% recombination efficiency in the Purkinje cell layer of the cerebellum.
Previous research using Opn3-mCh mice revealed additional brain regions with Opn-mCh expression such as the mitral cell layer, the posterior hypothalamic nucleus, and the superior olivary complex in the pons [5]. However, in our Opn3-phiC31o mice, we observed only limited signals in the brain, mainly in the olfactory bulb, cerebral cortex, and the Purkinje cell layer of the cerebellum. It has also been reported that Opn3-mCh expression is abundant in hypothalamic formation, but in our Opn3-phiC31o mice, we observed only sparse signals in the preoptic area within sagittal brain sections. Moreover, although Opn3-mCh was reported to be most highly expressed in the thalamus, our model showed limited signals in the thalamus in sagittal brain sections. The advantage of the Opn3-mCh reporter mouse over other models is that the Opn3-mCherry fusion protein is expressed under the native promoter, thereby preserving endogenous levels of Opn3 with native expression throughout development [5]. This is important for further characterization of Opn3, as Opn3 is developmentally regulated, at least in the brain [2, 6, 48].
Opn3 expression in the developing nervous system has also been examined in previous studies using an Opn3 promoter-driven eGFP BAC transgenic mouse line [6]. It has been reported that Opn3-eGFP was detected in 25 new structures in the central and peripheral nervous system that were not previously reported to have Opn3 expression, including sensory ganglia, the olfactory epithelium, vestibular nuclei, and parafascicular nucleus [6]. Nevertheless, the regions of Opn3 expression reported in those studies were much broader than what we observed in Opn3-phiC31o; *ROSA26 *^MultiFPsΔPuro^ mice. Knowing that BAC-based GFP reporters have the possibility to be leaky and generate false positives [5], this discrepancy once again proves that the recombination efficiency in our newly generated mouse line was not satisfactory enough to thoroughly examine Opn3 expression in the CNS.
Opn3-eGFP has previously been reported to be expressed in RGCs [29], and thus we examined Opn3-phiC31o-mediated gene expression in the retina of our mouse line. We found extremely low recombination efficiency in the retina of the Opn3-phiC31o; *ROSA26 *^MultiFPsΔPuro^ mice. Although Opn3 promoter activity is observed throughout the ganglion cell layer at birth, the proportion of RGCs expressing Opn3 in the adult brain decreases and is restricted to a few cells, as reported in previous studies [29]. Since phiC31o-mediated recombination is therefore more likely to occur at birth, and mCerulean expression should be indelible, our results suggest that the recombination efficacy of phiC31o is insufficient to conduct recombination even at the neonatal stage. Previous studies have also shown that approximately 24% of Opn3-eGFP-positive RGCs co-express Opn4 [29]. Opn4-positive RGCs are categorized into M1 to M5 subtypes based on morphological characteristics, with M2 and M4 subtypes known to express Opn3-eGFP [49]. Therefore, sparse labelling through Opn3-phiC31o-mediated recombination may allow identification of RGCs co-expressing Opn4, especially M2 and M4 types. If a sufficient number of labeled RGCs are recovered for morphological analysis, this could potentially enable morphological characterization of these specific RGC populations.
Despite the low recombination efficiency of our mouse line, one of the positive achievements is the observation of cell morphological structure. Due to the low recombination efficiency, there are few mCerulean-expressing cells which allow us to easily observe the whole morphological structure of the RGCs and HCs. Mice have only one type of HC, called the B-type axon-bearing HC [50, 51], which shows the same morphological structure as the one stained in our slides. Although the number of HCs we discovered was severely low, which may raise doubt about the reliability of Opn3 expression in HCs, an Opn3 homolog, cTMT, has been reported to be expressed in chicken retinal HCs [8]. This provides evidence that Opn3 expression can be found in retinal HCs, and thus there is a high possibility that it is also expressed in other animals, such as mice in this case. Nevertheless, it should be noted that phiC31o-mediated recombination in the retina was extremely few and the number of labeled RGCs and HCs was limited. Further studies are needed to confirm the Opn3 expression pattern, as well as how these Opn3-expressing neurons respond to blue light and modulate intracellular signaling in an Opn3-dependent manner. However, it would be necessary to perform experiments using a system in which more RGCs and HCs can be labeled in the retina, rather than the Opn3-phiC31o; *ROSA26 *^MultiFPsΔPuro^ mice reported here.
With the generation of the Opn3-phiC31o mouse line, a future approach would be to generate an Opn5-Dre mouse line, and then to cross these two mouse lines with the existing Opn4-Cre mouse line [38, 39] and the *ROSA26 *^MultiFPsΔPuro^ reporter mice [40]. By crossing these 4 mouse lines, the expressions of the non-visual opsins, Opn3, Opn4, and Opn5, could be studied simultaneously within the same mouse individual, since phiC31o, Cre, and Dre drivers mediate the recombination and express different fluorescent proteins, mCerulean, Venus, and mCherry, respectively, as reported previously [40].
There are several limitations in this experiment that should be considered in future studies, which may arise from technical aspects of the procedures. First, in the analysis of the transcription start site of Chml gene using the 5' RACE method, we used clones derived from the cerebral cortex and cerebellum of a single 3-month-old male C57BL/6 J mouse. We did not confirm whether the same transcription start site of Chml could be consistently identified across multiple animals. Next, the recombination efficiency of phiC31o was extremely low, resulting in very sparse mCerulean labeling in Opn3-phiC31o^+/+^*; ROSA *^MultiFPS^ mice. Although we made every effort to analyze the brain as comprehensively as possible, it is possible that we failed to detect all the sparsely distributed recombination-positive cells. The absence of mCerulean-positive cells in the paraventricular nucleus may be attributable to this limitation. To more comprehensively visualize mCerulean-positive cells, the use of tissue clearing techniques may have been beneficial.
Conclusions
We have developed an Opn3-phiC31o KI mouse line. Although phiC31o exhibits low recombination efficiency in the brain and retina, this line serves as a valuable resource for the random and sparse labelling of individual cells, facilitating detailed morphological characterization. Additionally, it can function as an Opn3 null model without perturbing Chml, thereby expanding its utility for diverse experimental applications.
Supplementary Information
Supplementary Figure 1. In situ hybridization results using sense probe showed no signals in brain sections. Representative sagittal sections of the cerebellum and thalamus from wild-type and Opn3-phiC31o homozygous mice were subjected to ISH using the Opn3 sense probe, phiC31o sense probe, and Chml sense probe. Opn3 mRNA expression in the Opn3-phiC31o homozygous mouse brain was not detected. Scale bars: 300 µm.