Adjuvant potential of Satureja hortensis metabolites with antibiotics against Gram-positive and Gram-negative bacterial strains
Marah Alburqan, Annamária Kincses, Melinda Paizs, Anita Barta, Katalin Veres, Antal Csámpai, Morteza Yazdani, Judit Hohmann

TL;DR
This study explores how compounds from Satureja hortensis can boost the effectiveness of antibiotics against drug-resistant bacteria.
Contribution
The study identifies specific plant metabolites that act as antibiotic adjuvants, reducing required drug dosages.
Findings
Naringenin, rosmarinic acid, and its methyl ester significantly reduced gentamicin MICs against drug-resistant E. coli.
Combinations of plant compounds with antibiotics like ciprofloxacin and ampicillin showed reduced MICs against K. pneumoniae and MRSA.
The study demonstrates that these combinations can lower antibiotic dosages while maintaining efficacy.
Abstract
The global rise in bacterial resistance to existing antibiotics has led to increased mortality rates and escalating healthcare costs, underscoring the urgent need for new classes of antibacterial agents that can act synergistically with current antimicrobials. This study aimed to isolate bioactive compounds from Satureja hortensis L. (Lamiaceae) and evaluate their effects in combination with ciprofloxacin (CIP), ampicillin (AMP), gentamicin (GEN), and tetracycline (TET). The adjuvant potential of naringenin (N), 5,6-dihydroxy-7,3′,4′-trimethoxyflavone (TMF), cilicione-a (Cil), rosmarinic acid (Rs), rosmarinic acid methyl ester (MRs), and fulgidic acid was evaluated by determining the minimum inhibitory concentrations (MICs) of selected antibiotics in presence of sub-inhibitory concentrations of these compounds. The assays were performed against ten bacterial strains using the twofold…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
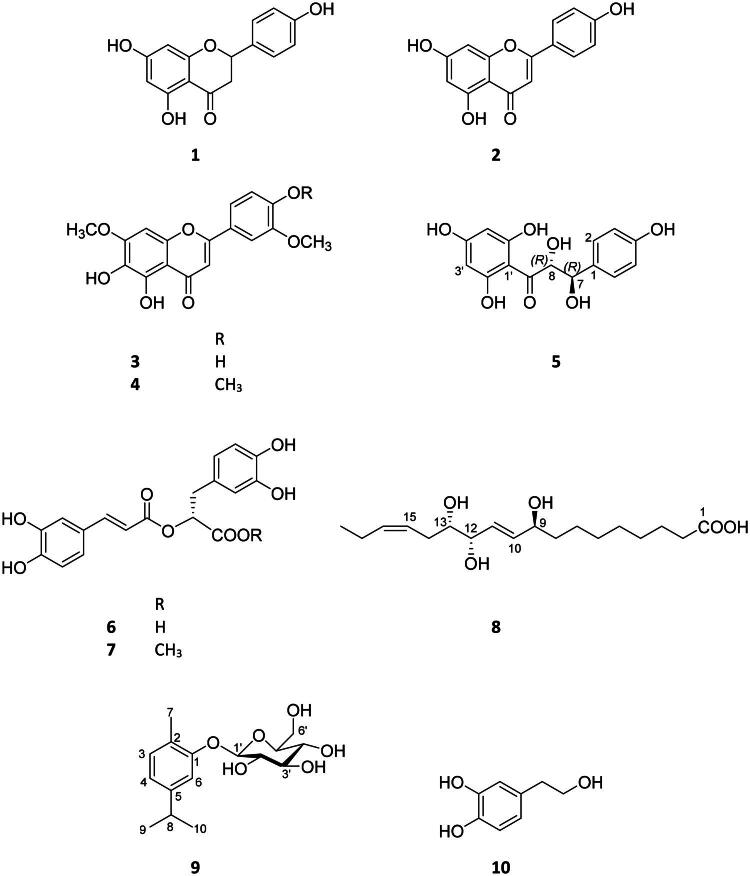 Figure 1
Figure 1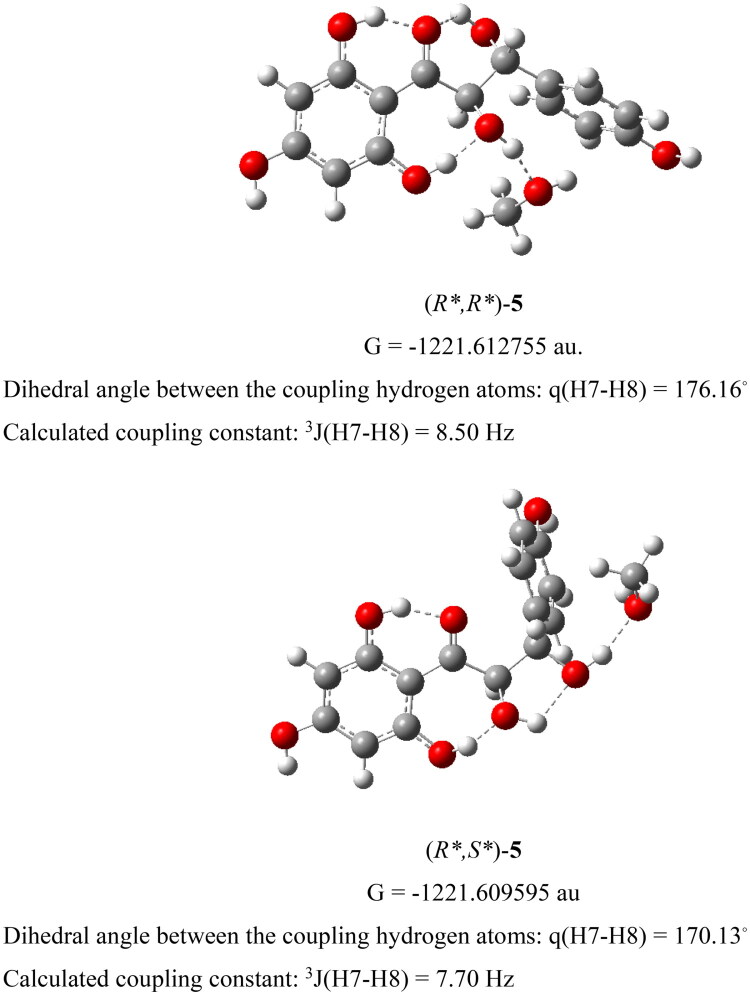 Figure 2
Figure 2| Bacterial strains | MIC (µM) | MIC (v/v %) | |||||
|---|---|---|---|---|---|---|---|
| 1, 4–8 | CIP | AMP | GEN | TET | DMSO | ||
| Gram-positive bacteria | >200 | 1.25 | 5 | 1.25 | 2.5 | >1 | |
| >200 | 1.25 | 50 | 100 | 2.5 | >1 | ||
| >200 | 0.39 | 3.125 | 0.0625 | >100 | >1 | ||
| >200 | 1.56 | 6.25 | 25 | 100 | >1 | ||
| >200 | 0.125 | 0.5 | 0.125 | 0.625 | >1 | ||
| Gram-negative bacteria | >200 | 0.039 | >100 | 1.25 | 6.25 | >1 | |
| >200 | 0.049 | 25 | 0.78 | 6.25 | >1 | ||
| >200 | 0.0156 | 6.25 | 0.78 | 6.25 | >1 | ||
| >200 | 3.125 | >100 | 12.5 | >100 | >1 | ||
| >200 | 0.625 | >100 | 1.25 | >100 | >1 | ||
| Bacterial strains | MIC (µg/mL) | ||||
|---|---|---|---|---|---|
| CIP | AMP | GEN | TET | ||
| Gram-positive bacteria | 0.414 | 1.857 | 1.861 | 1.202 | |
| 0.414 | 18.570 | 148.88 | 1.202 | ||
| 0.129 | 1.161 | 0.093 | >48.09 | ||
| 0.517 | 2.321 | 37.22 | 48.09 | ||
| 0.041 | 0.186 | 0.186 | 0.301 | ||
| Gram-negative bacteria | 0.012 | >37.139 | 1.861 | 3.006 | |
| 0.016 | 9.285 | 1.163 | 3.006 | ||
| 0.005 | 2.321 | 1.163 | 3.006 | ||
| 1.035 | >37.139 | 18.61 | >48.09 | ||
| 0.207 | >37.139 | 18.61 | >48.09 | ||
| Bacterial strains | MIC (µM) | |||||||
|---|---|---|---|---|---|---|---|---|
| CIP | CIP + | |||||||
| 1 | 4 | 5 | 6 | 7 | 8 | |||
| Gram-positive bacteria | 1.25 |
| None | None | None | None | None | |
| 1.25 |
| None | None |
|
| None | ||
| 0.39 | None | None | None | None | None | None | ||
| 1.56 | None | None | ND | None |
| None | ||
| 0.125 |
|
|
|
|
|
| ||
| Gram-negative bacteria | 0.039 | 2-fold increase | None | ND | None | None | None | |
| 0.049 | None | None | ND | None | 2-fold increase | None | ||
| 0.0156 | 2-fold increase | 2-fold increase | None | 2-fold increase | 2-fold increase | 2-fold increase | ||
| 3.125 |
| 2-fold increase |
|
|
| 2-fold increase | ||
| 0.625 |
| 2-fold increase |
|
|
| 2-fold increase | ||
| Bacterial strains | MIC (µM) | |||||||
|---|---|---|---|---|---|---|---|---|
| GEN | GEN + | |||||||
| 1 | 4 | 5 | 6 | 7 | 8 | |||
| Gram-positive bacteria | 1.25 |
|
| None | None | None |
| |
| 100 |
| None |
|
|
| None | ||
| 0.0625 | None | None | ND | None | None | None | ||
| 25 |
| None | ND |
|
| None | ||
| 0.125 |
|
|
|
|
|
| ||
| Gram-negative bacteria | 1.25 | 2-fold increase | None | ND | None | 2-fold increase | None | |
| 0.78 |
|
| ND |
|
| None | ||
| 0.78 | 2-fold increase | None | 2-fold increase | 2-fold increase | 2-fold increase | None | ||
| 12.5 | None | None | None | 2-fold increase | 2-fold increase |
| ||
| 1.25 | None | None | None | None | None | None | ||
| Bacterial strains | MIC (µM) | |||||||
|---|---|---|---|---|---|---|---|---|
| AMP | AMP + | |||||||
| 1 | 4 | 5 | 6 | 7 | 8 | |||
| Gram-positive bacteria | 5 |
| None | None | None | None |
| |
| 50 |
| None |
|
|
| None | ||
| 3.125 |
| None | ND |
|
| None | ||
| 6.25 |
| None | ND |
|
| None | ||
| 0.5 |
| None |
|
| None | None | ||
| Gram-negative bacteria | >100 | None | None | None | None | None | None | |
| 25 |
| None | ND |
|
| None | ||
| 6.25 | None | None |
| None |
| None | ||
| >100 | None | None | None | None | None | None | ||
| >100 | None | None | None | None | None | None | ||
| Bacterial strains | MIC (µM) | |||||||
|---|---|---|---|---|---|---|---|---|
| TET | TET + | |||||||
| 1 | 4 | 5 | 6 | 7 | 8 | |||
| Gram-positive bacteria | 2.5 | None | None | None | None | None | None | |
| 2.5 |
| None |
|
|
| None | ||
| >100 | None | None | ND | None | None | None | ||
| 100 |
| None | ND |
|
| None | ||
| 0.625 | None | None | None | None | None | None | ||
| Gram-negative bacteria | 6.25 | 2-fold increase | None | ND | None | None | None | |
| 6.25 | None | None | ND | None | None | None | ||
| 6.25 | None | None | None | None | None | None | ||
| >100 | None | None | None | None | None | None | ||
| >100 | None | None | None | None | None | None | ||
- —Ministry of Innovation and Technology of Hungary
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEssential Oils and Antimicrobial Activity · Phytochemistry and Biological Activities · Medicinal Plant Research
Introduction
The rise and global dissemination of multidrug-resistant (MDR) bacteria pose a major public health challenge, significantly compromising the efficacy of current treatments for infectious diseases. According to the World Health Organization (WHO), the progressive reduction in therapeutic options—primarily driven by bacterial resistance—has led to increased morbidity and mortality associated with bacterial infections (Matsunaga and Hayakawa 2018; Getahun et al. 2022). In response, intensified research aimed at identifying novel antimicrobial agents, optimizing existing therapeutic combinations, and elucidating synergistic interactions is essential for developing effective antibacterial strategies.
Synergistic combinations with antibiotics exert effects greater than the sum of their individual actions, enhancing efficacy while reducing required doses and minimizing side effects. The diminished effectiveness of antibiotics is influenced not only by microbial resistance but also by host-related factors such as immunodeficiency, advanced age, transplantation, and physiological stress. Recent studies have highlighted the potential of synergistic interactions between antibiotics and herbal compounds or phytochemicals. Co-administration of phytochemicals with antibiotics has been shown to enhance antibacterial efficacy and suppress the emergence of resistance (Ayaz et al. 2019). Additionally, antibiotic adjuvants can potentiate antibacterial activity by directly targeting bacterial mechanisms, bypassing intrinsic resistance, or improving antibiotic performance within the host (Dhanda et al. 2023).
The genus Satureja L. (savory) comprises approximately 200 aromatic herbs and shrubs with global distribution. Species within this genus are used in traditional and modern medicine, as well as in culinary and agricultural applications. Traditionally, Satureja species have been employed to treat respiratory and gastrointestinal disorders and are recognized for their antifungal and antibacterial properties. Satureja hortensis L. (summer savory) (Lamiaceae), an annual species native to southeastern Europe and western Asia, is widely cultivated as a culinary herb. Its extracts have demonstrated notable antioxidant, cytotoxic, and antibacterial activities in various bioassays (Ejaz et al. 2023).
This study aimed to identify the bioactive constituents of S. hortensis and evaluate their adjuvant potential in combination with various antibiotics. The antibacterial activities of ten compounds (1–10), isolated from the methanolic extract of the aerial parts, were assessed individually and in combination with conventional antibiotics against clinically relevant Gram-positive and Gram-negative bacterial strains.
Materials and methods
Experimental procedures of compound isolation and structure determination
Flash chromatography (FC) was performed on a CombiFlash Rf + Lumen instrument with integrated UV, UV–Vis, and ELS detection using a reversed-phase (RP) flash column (RediSep C_18_ Bulk 950, Teledyne Isco, Lincoln, NE, USA, Product No. Lab-Ex Kft. 692203336). RP high-performance liquid chromatography (RP-HPLC) and normal-phase HPLC (NP-HPLC) separations were performed on a Shimadzu LC-10 A S HPLC instrument equipped with a UV–Vis detector (Shimadzu, Co., Ltd., Kyoto, Japan) using RP-HPLC (LiChrospher RP-18, 5 μm, 250 × 4 mm, Merck, Darmstadt, Germany, Product No. Lab-Ex Kft. 00 G-3050-D0) and NP-HPLC (LiChrospher Si 60, 5 μm, 250 × 4 mm, Merck, Product No. Gen-Lab Kft., 00 G-3047-D0) columns, respectively. Sephadex LH-20 (25–100 μm, Cytiva, Sweden AB, Uppsala, Sweden, Product No. Merck GE17-0090-10) was used for gel filtration. Vacuum liquid chromatography (VLC) was performed on silica gel (GF_254_, 15 µm, Product No. Merck, 1.07730). Thin-layer chromatography (TLC) was performed using silica plates (20 × 20 cm silica gel 60 F_254_, Product No. Merck 1055540001). Analytical grade solvents were supplied by Molar Chemicals Kft. (Halásztelek, Hungary): methanol (Product No. 05730-006-410), n-hexane (Product No. 06850-101-411), cyclohexane (Product No. 01960-101-340), chloroform (Product No. 04730-101-411), ethyl acetate (Product No. 02930-101-411), etanol (Product No. 02911-101-340), and acetonitril (Product No. 06850-101-411). Solvents for HPLC were purchased from VWR International Kft. (Debrecen, Hungary): ethyl acetate (Product No. 83621.320), cyclohexane (Product No. 83629.320), methanol (Product No. 20864.320), and acetonitril (Product No. 20060.320). The water was supplied by the Milli-Q Direct 3 UV pump (Merck, Darmstadt, Germany). The structures were determined by one-dimensional (1D) (^1^H NMR,^13^C NMR JMOD) and two-dimensional (2D) NMR (^1^H-^1^H correlation spectroscopy [COSY], nuclear overhauser effect spectroscopy, heteronuclear single quantum coherence [HSQC], and heteronuclear multiple bond correlation [HMBC]) spectroscopy. NMR spectra were recorded on a Bruker Avance DRX 500 spectrometer (Bruker, Billerica, MA, USA) at 500 MHz (^1^H) and 125 MHz (^13^C). The signals of the deuterated solvents were taken as references. 2D NMR measurements were performed with standard Bruker software. In the ^1^H-^1^H COSY, HSQC, and HMBC experiments, gradient-enhanced versions were applied.
Plant material
Aerial parts of S. hortensis L. (Lamiaceae) were collected in the flowering stage by the grower Ferenc Okvátovity in Bátya, Hungary, in August 2022. A voucher specimen (Voucher No. 948) was deposited at the Herbarium of Department of Pharmacognosy, University of Szeged, Hungary. The plant material was evenly spread in a thin layer and air-dried at room temperature under protected conditions.
Extraction and isolation
The dried aerial parts of S. hortensis (1.9 kg) were exhaustively percolated with methanol (12 L) at room temperature. The resulting extract was concentrated under reduced pressure, dissolved in 50% aqueous methanol, and subjected to solvent–solvent partitioning with n-hexane (10 × 600 mL), chloroform (10 × 600 mL), and ethyl acetate (10 × 600 mL).
The chloroform-soluble fraction (8.27 g) was subjected to open-column chromatography on polyamide (150 g) and eluted with a stepwise methanol–water gradient (1:4, 2:3, 3:2, 4:1, v/v) followed by 100% methanol, yielding fractions A–E corresponding to 20%, 40%, 60%, 80%, and 100% methanol elutions, respectively. Fraction B (1.28 g), eluted with 40% methanol, was further purified by reversed-phase FC (RP-FC) on RP-silica gel (24 g) using a linear gradient of water–methanol (0% to 100%) over 75 min. Fractions with similar TLC profiles (B1–B22) were combined. Crystallization of fraction B15 yielded pure fulgidic acid (8) (115.6 mg). Fraction C (1.92 g), eluted with 60% methanol, was subjected to RP-FC under the same conditions (RP-silica gel, 24 g; water–methanol gradient, 0% to 100%) over 70 min, affording fractions C1–C16 based on TLC monitoring. From fraction C6, compound 4—identified as 5,6-dihydroxy-7,3′,4′-trimethoxyflavone (4) (113.5 mg)—was crystallized using methanol as the solvent. Fraction D (1.06 g), eluted with 80% methanol, was separated by RP-FC on RP-silica gel (24 g) using a linear water–methanol gradient (20% to 100%) over 70 min, yielding subfractions D1–D10. Subfraction D2 was further purified by Sephadex LH-20 gel filtration with methanol as the eluent, affording five subfractions (D2/1–D2/5). Subfraction D2/4 was further purified by preparative TLC on silica gel plates using chloroform–methanol (97:3, v/v) as the developing solvent, followed by NP-HPLC using an isocratic ethyl acetate–cyclohexane (70:30, v/v) solvent system, yielding naringenin (1) (1.62 mg). From subfraction D2/5, a crystalline material identified as 5,6,4′-trihydroxy-7,3′-dimethoxyflavone (3) (1.3 mg) was obtained.
The ethyl acetate-soluble fraction (20.5 g) was subjected to normal-phase VLC (NP-VLC) on silica gel using a gradient system of cyclohexane–ethyl acetate–ethanol (60:15:0, 60:20:0.5, 60:30:1, 60:30:3, 60:30:5, 60:30:10, 60:30:20, and 50:50:30, v/v/v), followed by ethyl acetate–methanol (8:2, v/v). Fractions were combined based on TLC monitoring to yield 13 pooled fractions (F1–F13). Fraction F3 (159.31 mg) was purified by RP-FC on RP-silica gel (40 g) using a linear water–acetonitrile gradient (0% to 100%) over 60 min, affording subfractions F3/1–F3/15. Crystallization of subfraction F3/13 from methanol yielded a crystalline compound identified as cilicione-a (5) (3.88 mg). Compound 2 (2.21 mg), identified as apigenin, was isolated from subfraction F3/15 by repeated RP-FC. Fraction F4 (799.21 mg) was further separated by RP-FC on RP-silica gel (40 g) using a linear water–methanol gradient (0% to 100%) over 70 min, yielding subfractions F4/1–F4/15. Subfraction F4/3 (67.37 mg) was further purified by gel filtration on Sephadex LH-20 using methanol as the eluent, followed by RP-HPLC with an isocratic water–methanol system (70:30, v/v), yielding 3-hydroxytyrosol (10) (3.69 mg). Fraction F6 (3.075 g) was purified by RP-FC on RP-silica gel (40 g) using a linear water–acetonitrile gradient (0% to 100%) over 75 min, affording subfractions F6/1–F6/21. Subfraction F6/15 (273.47 mg) was further separated by RP-FC using a similar gradient (0% to 100% acetonitrile in water over 70 min), yielding subfractions F6/15.1–F6/15.13. Subfraction F6/15.7 was purified by RP-HPLC with an isocratic acetonitrile–water system (1:1, v/v), affording carvacrol-β-O-glucoside (9) (1.46 mg). Subfraction F6/15.9 (133 mg) was subjected to RP-VLC on C_18_ silica gel using a water–acetonitrile gradient (10% to 50%), yielding rosmarinic acid methyl ester (7) (6.65 mg). Furthermore, subfraction F6/18 (38.91 mg) was purified by RP-VLC using a water–acetonitrile gradient (0% to 50%) to yield rosmarinic acid (6) (7.33 mg).
5,6,4′-Trihydroxy-7,3′-dimethoxyflavone (3): pale yellow crystals; Mp. 268 °C–271 °C; ^1^H NMR (500 MHz, CD_3_OD, δ ppm): 7.53 (1H, dd, J = 8.2, 1.7 Hz, H-6′), 7.50 (1H, d, J = 1.8 Hz, H-2′), 6.92 (1H, d, J = 8.3, H-5′), 6.84 (1H, s, H-8), 6.64 (1H, s, H-3), 3.99 (3H, s, 7-OCH_3_), and 3.97 (3H, s, 3′-OCH_3_); ^13^C NMR (125 MHz, CD_3_OD, δ ppm): 184.3 (C-4), 166.5 (C-2), 155.8 (C-7), 152.8 (C-4′), 152.0 (C-9), 149.7 (C-3′), 147.2 (C-5), 131.5 (C-6), 123.4 (C-1′), 121.8 (C-6′), 117.0 (C-5′), 110.6 (C-2′), 106.6 (C-10), 103.7 (C-3), 91.9 (C-8), 57.0 (7-OCH_3_), and 56.7 (3′-OCH_3_).
Cilicione-a (5): yellow solid; ^1^H NMR (500 MHz, CD_3_OD, δ ppm): 7.35 (2H, d, J = 8.2 Hz, H-2, H-6), 6.83 (2H, J = 8.2 Hz, H-3, H-5), 5.93 (1H, s, H-8), 5.89 (1H, s, H-3′), 4.98 (1H, d, J = 11.6 Hz, H-7), 4.54 (1H, d, J = 11.6 Hz); ^13^C NMR (125 MHz, CD_3_OD, δ ppm): 198.5 (C-9), 168.9 (C-4′), 165.3 (C-6′), 164.6 (C-2′), 159.2 (C-4), 130.4 (C-2, C-6), 129.3 (C-1), 116.2 (C-3, C-5), 101.8 (C-1′), 97.4 (C-5′), 96.4 (C-3′), 85.0 (C-7), 73.7 (C-8).
Fulgidic acid (8): white amorphous solid; ^1^H NMR (500 MHz, DMSO-d6, δ ppm): 5.58 (2H, m, H-10, H-11), 5.40 (1H, dt, J = 6.8, 10.9 Hz, H-15), 5.36 (1H, dt, J = 6.8, 10.9 Hz, H-16), 3.90 (1H, brs, H-9), 3.82 (1H, m, H-12), 3.28 (1H, m, H-13), 2.19 (1H, m, H-14a), 2.17 (2H, t, J = 7.4 Hz, H-2), 1.97 (2H, t, J = 7.5 Hz, H-17), 1.91 (1H, m, H-14b), 1.47 (2H, m, H-3), 1.36 (2H, m, H-8), 1.24 (8H, m, H-4–H-7), and 0.97 (3H, t, J = 7.5 Hz, H-18); ^13^C NMR (125 MHz, DMSO-d6, δ ppm): 174.5 (C-1), 134.6 (C-10), 131.9 (C-16), 129.3 (C-11), 126.6 (C-15), 74.0 (C-13), 73.6 (C-12), 70.5 (C-9), 37.4 (C-8), 33.7 (C-2), 30.0 (C-14), 29.0, 28.8, 28.5, 24.9 (C-4–C-7), 24.5 (C-3), 20.2 (C-17), and 14.1 (C-18).
Carvacrol-β-O-glucoside (9): colorless oil; ^1^H NMR (500 MHz, CD_3_OD, δ ppm): 7.02 (1H, d, J = 8.0 Hz, H-3), 7.01 (1H, brs, H-6), 6.78 (1H, brd, J = 7.7 Hz, H-4), 4.86 (1H, H-2′), d, J = 7.1 Hz, H-1′), 3.89 (1H, d, J = 11.9 Hz, H-6’a), 3.70 (1H, dd, J = 5.1, 11.9 Hz, H-6’b), 3.48 (1H, m, H-2′), 3.47 (1H, m, H-3′), 3.41 (1H, m, H-5′), 3.40 (1H, m, H-4′), 2.84 (1H, sept, J = 6.6 Hz), 2.22 (3H, s, H-7), and 1.22 (6H, d, J = 6.6 Hz, H-9, H-10); ^13^C NMR (125 MHz, CD_3_OD, δ ppm): 157.2 (C-1), 149.1 (C-5), 131.4 (C-3), 126.2 (C-2), 121.2 (4), 114.6 (C-6), 102.9 (C-1′), 78.3 (C-3′), 78.2 (C-5′), 75.1 (C-2′), 71.6 (C-4′), 62.7 (C-6′), 35.2 (C-8), 24.5 (C-10), 24.4 (C-9), and 16.1 (C-7). NMR data are in agreement with the literature values (Mastelić et al., 2004).
Comparative density functional theory (DFT) modeling study of cilicione-a (5)
Calculations were carried out with M062X global functional (Zhao and Truhlar 2008) using 6–31 G(d,p) basis set (Hehre et al. 1986) and supplemented by the integral equation formalism polarizable continuum solvent model (Tomasi et al. 1999). The diagnostic theoretical coupling constants were calculated using the online generalized ^3^JHH calculation program elaborated by Haasnoot et al. (1980).
Bacterial strains
In this study, the following Gram-positive bacterial strains were examined: Staphylococcus aureus ATCC 29213; methicillin- and oxacillin-resistant S. aureus (MRSA) ATCC 43300; Staphylococcus epidermidis ATCC 12228; Enterococcus faecalis ATCC 29212; and Bacillus subtilis ATCC 6633. The Gram-negative bacterial strains tested included Escherichia coli ATCC 35218; E. coli K-12 AG100, which expresses the AcrAB-TolC efflux pump at basal levels; Salmonella enterica serovar Typhimurium SL1344 (SEO1); Klebsiella pneumoniae ATCC 700603; and Pseudomonas aeruginosa ATCC 27853. The Salmonella strain was kindly provided by Dr. Jessica M. A. Blair (University of Birmingham, UK).
Determination of minimum inhibitory concentrations (MICs)
The MICs of all tested compounds and antibiotics were determined according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI 2018). MICs were established using twofold serial dilutions of the compounds (200–0.0195 µM) in Mueller–Hinton Broth (MHB, Biolab Zrt., Product No. MHC20500), with a standardized bacterial inoculum of 5 × 10^5^ CFU/mL. The turbidity of the bacterial suspension was adjusted using a McFarland densitometer (Biosan, Riga, Latvia). Assays were performed in 96-well microtiter plates and incubated at 37 °C for 18 h. MICs were determined visually at the end of the incubation period. Dimethyl sulfoxide (DMSO, Molar Chemicals Kft., Product No. 02610-526-340) served as the solvent control, with its final concentration (2% v/v) verified to have no antibacterial activity. Results are presented as mean values from three replicates, each conducted in three independent experiments. The reference antibiotics—ciprofloxacin (CIP) (Product No. Merck 17850-5 G-F**)**, ampicillin sodium crystalline (AMP) (Sigma Aldrich, Product No. A9518-5G), gentamicin sulfate (GEN) (Sigma Aldrich, Product No. G1264-250), and tetracycline hydrochloride (TET) (Sigma Aldrich, Product No. T7660-5G)—were used.
Enhancement of the activity of antibiotics
The chemosensitizing potential of compounds 1 and 4–8 was evaluated by determining the MIC values of selected antibiotics in the presence of fixed sub-inhibitory concentrations (100 µM) of each compound against Gram-positive and Gram-negative bacterial strains. MICs were determined using the standard twofold broth microdilution method in 96-well microtiter plates, with serial dilutions of CIP, AMP, GEN, and TET. In each assay, the first four rows of the plate contained twofold serial dilutions of the antibiotics alone, while the last four rows assessed the antibiotic–compound combinations. A 10^−4^ dilution of overnight bacterial cultures in 50 µL of MHB was added to each well, except for medium control wells. Plates were incubated at 37 °C for 18 h, and MICs were determined visually at the end of the incubation period. All results are reported as mean values from three biological replicates, each conducted in three independent experiments.
Results
In this study, the aerial parts of S. hortensis were subjected to detailed phytochemical investigation. Ten compounds (1–10) were isolated from the chloroform- and ethyl acetate-soluble phases of the methanolic extract using a multistep chromatographic purification procedure. The structures of the isolated compounds were elucidated by spectroscopic methods and identified as naringenin (1), apigenin (2), 5,6,4′-trihydroxy-7,3′-dimethoxyflavone (3), 5,6-dihydroxy-7,3′,4′-trimethoxyflavone (4), cilicione-a (5), rosmarinic acid (6), rosmarinic acid methyl ester (7), fulgidic acid (8) [= (9S,12S,13S)-9,12,13-trihydroxy-10E,15Z-octadecadienoate], carvacrol-β-O-glucoside (9), and 3-hydroxytyrosol (10) (Figure 1).
Structure of the investigated compounds: naringenin (1), apigenin (2), 5,6,4′-trihydroxy-7,3′-dimethoxyflavone (3), 5,6-dihydroxy-7,3′,4′-trimethoxyflavone (4), cilicione-a (5), rosmarinic acid (6), rosmarinic acid methyl ester (7), fulgidic acid (8), carvacrol-β-O-glucoside (9), and 3-hydroxytyrosol (10).
Compounds 1 and 4–8, in sufficient amounts, in combination with antibiotics were investigated for their possible adjuvant effects. In this study, antibiotics representing different structural classes were selected: CIP (fluoroquinolone class), GEN (aminoglycoside), AMP (β-lactam), and TET (tetracyclic derivative). These antibiotics exhibit distinct mechanisms of action, targeting the bacterial cell wall (AMP), nucleic acids (CIP), or protein biosynthesis (GEN and TET). First, the MICs were determined for 1 and 4–8 (Table 1(A)) and for the antibiotics (Table 1(B)) against five Gram-positive and five Gram-negative bacterial strains. The MICs of all compounds were >200 µM, whereas most of the bacterial strains used in the study were sensitive to antibiotics, with some exceptions (MIC >100 µM) presented in Table 1(A).
The MICs of antibiotics were subsequently determined in the presence of a sub-inhibitory concentration (100 µM) of each compound in both Gram-positive and Gram-negative bacterial strains to evaluate their chemosensitizing potential. The potentiating effects of the antibiotics and S. hortensis metabolites were assessed using a twofold broth microdilution method in 96-well plates. The results are presented in [Tables 2–5](#t0003 t0004 t0005 t0006).
The combination of naringenin (1), cilicione-a (5), rosmarinic acid (6), and rosmarinic acid methyl ester (7) with CIP increased the potency of CIP in two- to eightfold in some bacteria (Table 2). K. pneumoniae ATCC 700603 exhibited the highest susceptibility to naringenin (1), cilicione-a (5), rosmarinic acid (6), and rosmarinic acid methyl ester (7) combined with CIP, reducing the MIC of CIP from 3.125 to 0.39 µM (eightfold) by 1 and 7 and to 0.78 µM (fourfold) by 5 and 6. 5,6-Dihydroxy-7,3′,4′- trimethoxyflavone (4) and fulgidic acid (8) decreased the MIC of CIP twofold only against B. subtilis; however, they reduced the effectiveness of CIP by half against S. Typhimurium SEO1, K. pneumoniae ATCC 700603, and P. aeruginosa ATCC 27853. S. Typhimurium SEO1 was the only strain in which all compounds, except for 5, caused a twofold increase in the MIC of CIP.
Combination of compounds with GEN resulted in the highest MIC reduction effect of naringenin (1), rosmarinic acid (6), and rosmarinic acid methyl ester (7) (Table 3). In particular, 1 potentiated the effect of GEN in S. aureus ATCC 29213, S. aureus MRSA ATCC-43300, E. faecalis ATCC 29212, B. subtilis ATCC 6633, and E. coli AG100. Against the drug-resistant E. coli AG100 strain, combinations of GEN with naringenin (1), rosmarinic acid (6), and rosmarinic acid methyl ester (7) substantially modulated the effect of the antibiotic resulting in eightfold reductions in MICs (from 0.78 µM to 0.097 µM). In combinations with GEN and CIP, 1, 5, 6, and 7 demonstrated the ability to enhance antibiotic activity against B. subtilis ATCC 6633.
Combinations of naringenin (1), rosmarinic acid (6), and rosmarinic acid methyl ester (7) with AMP improved the antibiotic effect against most of the tested bacteria by two- or fourfold (Table 4). Naringenin (1) enhanced the activity of AMP against all Gram-positive bacteria and against E. coli AG100. Notably, against the drug-resistant S. aureus MRSA ATCC 43300 strain naringenin (1), cilicione-a (5), rosmarinic acid (6), and rosmarinic acid methyl ester (7) were found to potentiate the activity of AMP, resulting in a reduction of the MIC from 50 to 12.5 µM.
The summer savory metabolites naringenin (1), cilicione-a (5), rosmarinic acid (6), and rosmarinic acid methyl ester (7) showed the smallest reductions in MICs when combined with TET (Table 5). These compounds enhanced the antibiotic effect only against S. aureus MRSA ATCC 43300 and E. faecalis ATCC 29212, reducing the MICs by half. In contrast, 5,6-dihydroxy-7,3′,4′- trimethoxyflavone (4) and fulgidic acid (8) markedly enhanced the activity of TET, resulting in significantly lower MIC values.
Discussion
In traditional medicine, summer savory has been used to treat various respiratory and gastrointestinal disorders, many of which are associated with microbial infections. In recent years, numerous studies have demonstrated the antibacterial and antibiofilm activities of S. hortensis extracts and essential oils against a broad range of bacterial strains (Lesjak et al. 2016; Ramezani et al. 2016; Harmati et al. 2017; Masoum et al. 2018; Sharifi et al. 2018; Huwaimel et al. 2023). However, to date, no studies have investigated the interactions between the secondary metabolites of summer savory and conventional antibiotics.
Isolation and identification of compounds
Ten compounds were isolated from the aerial parts of S. hortensis; these compounds belong to the group of flavonoids [naringenin (1), apigenin (2), 5,6,4′-trihydroxy-7,3′-dimethoxyflavone (3), 5,6-dihydroxy-7,3′,4′- trimethoxyflavone (4), and cilicione-a (5)], cinnamic acid derivatives [rosmarinic acid (6) and rosmarinic acid methyl ester (7)], oxylipins [fulgidic acid (8)] and monoterpene glucosides [carvacrol-β-O-glucoside (9)] and phenylethyl alcohols [3-hydroxytyrosol (10)]. The structures were determined by NMR spectroscopy and comparison with published data. Naringenin (1), apigenin (2), rosmarinic acid (6), and rosmarinic acid methyl ester (7) are widely occurring in the plant kingdom, and their NMR assignments agreed well with data published by Zhang et al. (2006), Kim et al. (2006), Akoury (2019), and Sina Içen et al. (2021), respectively. 5,6,4′-Trihydroxy-7,3′-dimethoxyflavone (3) and 5,6-dihydroxy-7,3′,4′-trimethoxyflavone (4) were described previously from various Satureja species (Skoula et al. 2005; Maldonado and Ortega, 1997; Malmir et al. 2012; Davoodi et al. 2018; Nagao et al., 2002).
The dihydrochalcone derivative cilicione-a (5) has previously been isolated only from Thymus cilicicus (Ahmed and Al-Howiriny 2007), although the stereochemistry at C-7 and C-8 was not determined. To establish its relative configuration, the two possible diastereomers, (R,R**)-24 and (R*,S*)-24, were subjected to comparative DFT modeling. To better simulate the conditions of the NMR measurements, a methanol molecule was included in each model to complement the network of intramolecular hydrogen bonds, which are expected to restrict rotation around the C7–C8 bond. The calculations provided the relative thermodynamic stability of the two diastereomers, indicating that under NMR measurement conditions, (R,R**)-24 is more stable than (R,S**)-24 by 1.98 kcal/mol. Diagnostic theoretical vicinal coupling constants,^3^J(H7–H8), were estimated using the generalized ^3^JHH equation (Haasnoot et al. 1980), based on the dihedral angles obtained from the optimized structures. The calculated coupling constant for (R,R**)-24 (8.50 Hz) was closer to the experimental value (11.6 Hz) than that of (R,S**)-24 (7.70 Hz) (Figure 2). Taken together, the comparison of calculated and experimental coupling constants, along with the relative thermodynamic stability, supports the assignment of the compound as (R,R**)-24.
Optimized structure and gibbs free energy of measurements for cilicione-a (5) stereoisomers.
Fulgidic acid (8) was isolated here for the first time from the Lamiaceae family, and its NMR data agreed with data published by Zhang et al. (2024). Carvacrol-β-O-glucoside (9) was previously published from some Lamiaceae species: Monarda punctata (Yamada et al. 2010), Thymus vulgaris (Skopp and Hörster 1976), Lavandula multiflora (Sosa et al. 2005), and Origanum vulgare (Milos et al. 2000). However, this is the first report of its isolation from Satureja. The aglycon carvacrol is one of the main compounds of the essential oil of S. hortensis (Lesjak et al. 2016).
3-Hydroxytyrosol (10), a typical compound of olive leaves and fruits (Kalampaliki et al. 2019), was also identified from summer savory. To our knowledge, this is the first report of 10, 3, and 7 isolated from S. hortensis. Our 2D NMR studies allowed the assignment of NMR signals in solvents other than those previously reported for 3, 8, and 9 (Materials and methods).
Interactions of the isolated compounds with antibiotics
The interactions of conventional antibiotics—CIP, GEN, AMP, and TET—with six compounds (1, 4–8) isolated from S. hortensis were investigated. Among these, compounds 1 and 6 are widely distributed in the plant kingdom. Apigenin (2) was not included in the present study, as its interaction with antibiotics was previously evaluated under identical experimental conditions.
Prior to assessing the interactions, an initial antibacterial screening was performed against five Gram-positive and five Gram-negative bacterial strains to determine the MICs of the tested antibiotics and isolated compounds. None of the compounds exhibited antibacterial activity at concentrations up to 200 µM (MIC > 200 µM).
Among the tested combinations, B. subtilis ATCC 6633 exhibited the highest sensitivity, whereas S. enterica serovar Typhimurium SEO1 showed the lowest sensitivity to the compounds when combined with CIP, GEN, and AMP.
Naringenin (1) exhibited the strongest potentiating effect, enhancing the activities of CIP, GEN, and AMP. These combinations reduced the MICs by eightfold against K. pneumoniae (with CIP) and drug-resistant E. coli AG100 (with GEN), and by fourfold against S. aureus ATCC 29213 (with GEN), S. aureus MRSA ATCC 43300 (with GEN and AMP), and B. subtilis ATCC 6633 (with GEN). These results align with previous reports demonstrating the potentiating effects of naringenin (1). For example, Yi et al. (2024) reported an eightfold reduction in the MIC of amikacin against multidrug-resistant E. coli when combined with 1. Similarly, Mohammed et al. (2015) showed that 1 significantly decreased the MIC of CIP against a clinical MRSA isolate. Duda-Madej et al. (2022) also described a synergistic interaction between naringenin (1) and oxacillin against MRSA, while Ng’uni et al. (2015) observed synergism with AMP, TET, methicillin, and vancomycin against S. aureus and MRSA. Notably, the present study is the first to demonstrate the adjuvant potential of naringenin (1) in combination with antibiotics against E. faecalis ATCC 29212, B. subtilis ATCC 6633, K. pneumoniae ATCC 700603, and P. aeruginosa ATCC 27853.
Both rosmarinic acid (6) and its methyl ester (7) exhibited promising activity in combination assays, each reducing the MICs of CIP, GEN, AMP, and TET against four Gram-positive and four Gram-negative bacterial strains. Notably, a fourfold and eightfold potentiation was observed against the drug-resistant strains S. aureus MRSA ATCC 43300 and E. coli AG100 when combined with AMP and GEN, respectively. Rosmarinic acid methyl ester (7) demonstrated slightly greater potency than rosmarinic acid (6), as evidenced by a twofold MIC reduction of CIP against E. faecalis ATCC 29212 and of AMP against Salmonella Typhimurium SEO1, as well as a more pronounced reduction in the MIC of CIP against K. pneumoniae ATCC 700603. Although previous studies have reported synergistic effects of rosmarinic acid (6) in combination with antibiotics such as vancomycin, ofloxacin, and amoxicillin against S. aureus (Ekambaram et al. 2016), these interactions were not observed in the present study. Zhang et al. (2024) also demonstrated synergy between 6 and the cephalosporin ceftiofur in anti-MRSA activity. Therefore, the potentiating effects observed for 6 and 7 in this study represent novel findings.
Among the tested compounds, fulgidic acid (8) was the only one featuring an aliphatic chain structure. Likely due to the absence of a phenolic moiety, it exhibited only a weak ability to potentiate antibiotic activity. A twofold reduction in the MICs was observed in combination with CIP, AMP, and GEN against S. aureus ATCC 29213 (with AMP and GEN), B. subtilis ATCC 6633 (with GEN and CIP), and K. pneumoniae ATCC 700603 (with GEN).
When comparing the effectiveness of naringenin (1) and apigenin (2), which differ only by the presence of a C-2/C-3 double bond, both exhibited similar potentiating effects in combination with CIP against S. aureus ATCC 29213 and S. aureus MRSA 43300. However, apigenin (2) showed no substantial influence on antibiotic susceptibility in other strains, particularly Gram-negative bacteria (Kincses et al. 2024). 5,6-Dihydroxy-7,3′,4′-trimethoxyflavone (4), which also possesses a flavone backbone like 2, demonstrated similarly limited activity compared with naringenin (1), with its most notable effect being a fourfold MIC reduction of B. subtilis ATCC 6633 when combined with GEN.
Our assays provided insights into the antibiotic adjuvant potential of cilicione-a (5), a dihydrochalcone derivative. While previous studies have demonstrated that both natural and synthetic chalcones can significantly enhance the activity of various antibiotics (Lee et al. 2010; Tran et al. 2012; Božić et al. 2014), these investigations were limited to classical chalcones. In contrast, compound 5 is a 7,8-dihydro-7,8-dihydroxychalcone, representing a structurally distinct scaffold. Notably, 5 exhibited its most pronounced potentiating effect in combination with CIP against K. pneumoniae and with AMP against MRSA, each resulting in a fourfold reduction in MIC.
In certain cases, combinations of compounds 1 and 4–8 with CIP, GEN, and TET against the tested Gram-negative bacterial strains resulted in a twofold increase in MICs. Such increases in MIC are of particular concern in drug discovery and clinical therapy, as it may lead to reduced treatment efficacy, the requirement for higher antibiotic doses, or prolonged infections. Although clinically significant antagonistic interactions have been reported only infrequently, some examples include combinations of β-lactam and macrolide antibiotics with other bacteriostatic agents (Acar 2000).
The observed enhancement of antibiotic activity by the investigated compounds may arise through several mechanisms. One common mechanism is the increased permeability of the bacterial cell envelope, which facilitates antibiotic uptake. In other instances, direct physical interactions at the target site can enhance antibacterial activity. More complex synergy may involve alterations in bacterial physiology that indirectly improve antibiotic efficacy (Bollenbach 2015). For example, rosmarinic acid (6), has been shown to inhibit bacterial surface proteins, potentially impairing the ability of pathogens to evade antibiotic action. In MRSA and S. aureus, co-administration of 6 with a glycopeptide antibiotic significantly reduced the expression of microbial surface components recognizing adhesive matrix molecules compared with rosmarinic acid (6) alone (Ekambaram et al. 2016). Naringenin (1) exerts antimicrobial effects on MRSA strains by decreasing membrane fluidity in both hydrophilic and hydrophobic regions of the bacterial membranes (Duda-Madej et al. 2022).
Conclusions
This study demonstrated that S. hortensis is a valuable source of secondary metabolites (1–10) with the potential to enhance the efficacy of conventional antibiotics. Although six of the isolated compounds (1, 4–8) exhibited no intrinsic antibacterial activity at concentrations up to 200 µM, their use at sub-inhibitory levels significantly potentiated the activity of CIP, GEN, AMP, and TET against both Gram-positive and Gram-negative bacterial strains, including drug-resistant variants. The observed potentiating effects are likely mediated through multiple mechanisms, including alterations in membrane structure and permeability, inhibition of adhesion-related protein expression, and interference with key metabolic pathways. These findings underscore the potential of natural product–antibiotic combinations to improve therapeutic outcomes, reduce required antibiotic doses, and support strategies aimed at combating antimicrobial resistance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Acar JF. 2000. Antibiotic synergy and antagonism. Med Clin North Am. 84(6):1391–1406. 10.1016/S 0025-7125(05)70294-711155849 · doi ↗ · pubmed ↗
- 2Ahmed B, Al-Howiriny TA. 2007. Two new hydroxy chalcone derivatives from Thymus cilicicus. Z. Naturfoesch B. 62(1):121–124. 10.1515/znb-2007-0118 · doi ↗
- 3Akoury E. 2019. Isolation and structural elucidation of rosmarinic acid by nuclear magnetic resonance spectroscopy. Am Res J Chem. 1(1):17–23. 10.21694/2577-5898.17003 · doi ↗
- 4Ayaz M et al. 2019. Synergistic interactions of phytochemicals with antimicrobial agents: potential strategy to counteract drug resistance. Chem Biol Interact. 308:294–303. 10.1016/j.cbi.2019.05.05031158333 · doi ↗ · pubmed ↗
- 5Bollenbach T. 2015. Antimicrobial interactions: mechanisms and implications for drug discovery and resistance evolution. Curr Opin Microbiol. 27:1–9. 10.1016/j.mib.2015.05.00826042389 · doi ↗ · pubmed ↗
- 6Božić DD, Milenković M, Ivković B, Ćirković I. 2014. Antibacterial activity of three newly-synthesized chalcones & synergism with antibiotics against clinical isolates of methicillin-resistant Staphylococcus aureus. Indian J Med Res. 140(1):130–137.25222788 PMC 4181146 · pubmed ↗
- 7Davoodi M, Rustaiyan A, Ebrahimi SN. 2018. Monoterpene flavonoid from aerial parts of Satureja khuzistanica. Rec Nat Prod. 12(2):175–178. 10.25135/rnp.19.17.06.109 · doi ↗
- 8Dhanda G, Acharya Y, Haldar J. 2023. Antibiotic adjuvants: a versatile approach to combat antibiotic resistance. ACS Omega. 8(12):10757–10783. 10.1021/acsomega.3c 0031237008128 PMC 10061514 · doi ↗ · pubmed ↗