The Road Less Traveled: Exploring the Genomic Characteristics and Antimicrobial Resistance Potential of Acinetobacter baumannii From the Indigenous Orang Asli Community in Peninsular Malaysia
Soo‐Sum Lean, Denise E. Morris, Rebecca Anderson, Ahmed Ghazi Alattraqchi, David W. Cleary, Stuart C. Clarke, Chew Chieng Yeo

TL;DR
This study explores antibiotic-resistant Acinetobacter baumannii bacteria from healthy people in a remote Malaysian community, finding they share traits with hospital strains, suggesting public health importance.
Contribution
The study identifies novel genomic features and resistance genes in A. baumannii from an understudied indigenous community, revealing their potential clinical relevance.
Findings
Nine A. baumannii isolates from Orang Asli individuals showed genetic diversity, including new sequence types.
Isolates carried resistance genes like bla_OXA-51 and bla_ADC but remained susceptible to certain antibiotics.
Some isolates harbored mobile resistance gene modules on plasmids, indicating potential for spread.
Abstract
Acinetobacter baumannii is widely recognized as a multidrug‐resistant pathogen, although most public genome datasets are biased toward hospital‐derived isolates. Little is known about A. baumannii isolates from healthy individuals in the community. This study analyzed genome sequences from nine A. baumannii isolates obtained from the upper respiratory tract of the indigenous Orang Asli in their rural community in Peninsular Malaysia. Genomic analysis revealed genetic diversity, including three new Pasteur sequence types (STs) and six novel Oxford STs. One isolate, A. baumannii 19064, belonged to Global Clone 8 (GC8), a lineage linked to clinical infections. Core genome phylogeny showed these community isolates interspersed with clinical isolates from a nearby hospital, indicating potential pathogenicity under suitable conditions. All isolates carried intrinsic bla OXA‐51‐like…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
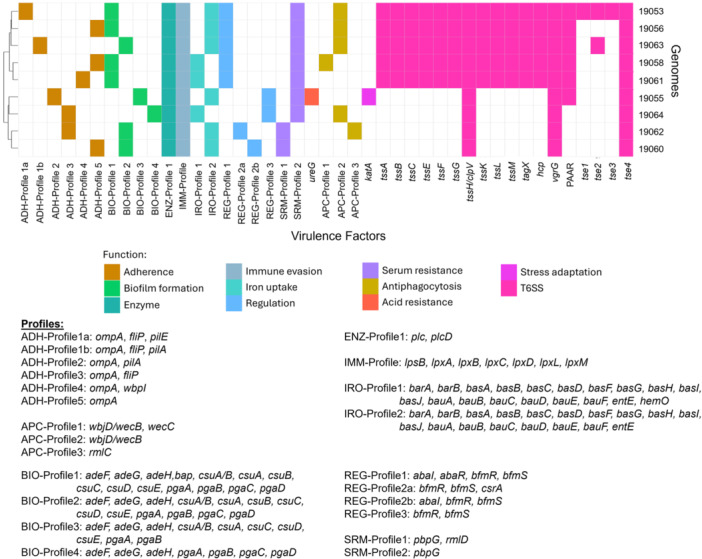 Figure 1
Figure 1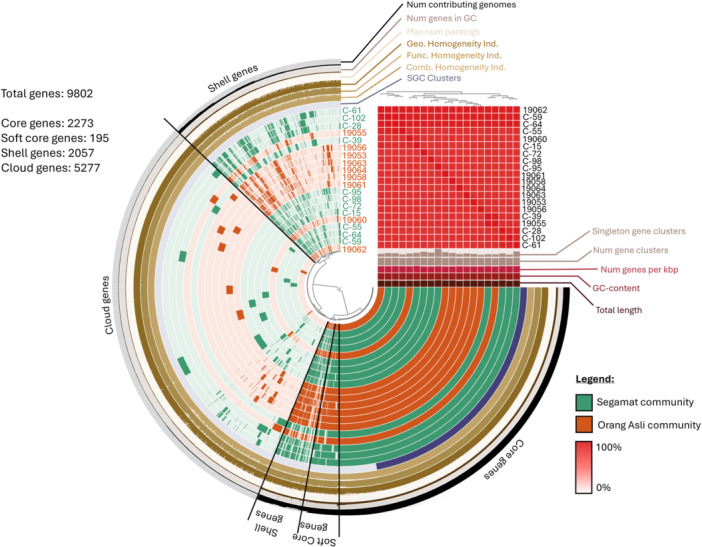 Figure 2
Figure 2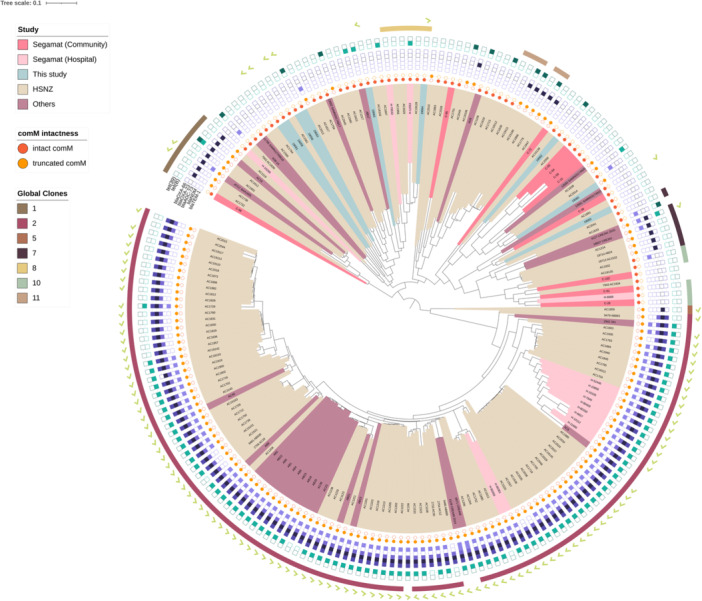 Figure 3
Figure 3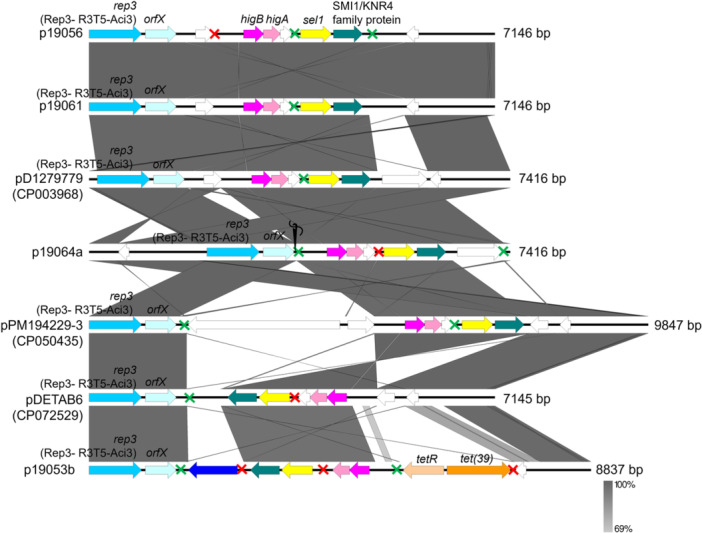 Figure 4
Figure 4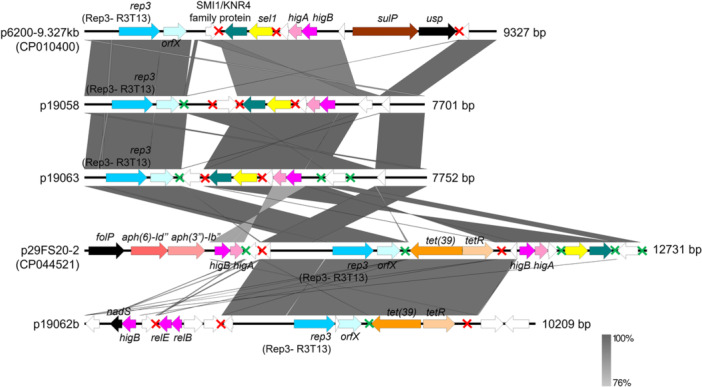 Figure 5
Figure 5|
| 19053 | 19055 | 19056 | 19058 | 19060 | 19061 | 19062 | 19063 | 19064 |
|---|---|---|---|---|---|---|---|---|---|
| Location/source | KSP/N | KSP/NP | KB/N | KB/N | KB/NP | KB/N | KB/NP | KB/N | KB/N |
| Species identification |
|
|
|
|
|
|
|
|
|
| Accession no. |
JBNPBH 000000000 |
JBNPBI 000000000 |
JBNPBJ 000000000 |
JBNPBK 000000000 |
JBNPBL 000000000 |
JBNPBM 000000000 |
JBNPBN 000000000 |
JBNPBO 000000000 |
JBNPBP 000000000 |
| Closest neighbor (fastANI) |
104305267 (98.1849%) |
STDY7716391 (99.8833%) |
STDY7716201 (97.9228%) |
STDY7715392 (98.7137%) |
104305309 (98.8467%) |
104305313 (98.0632%) |
03174920 (98.6532%) |
03174917 (97.9238%) |
104305269 (99.8239%) |
| Total genome size (bp) | 3,821,374 | 3,697,509 | 3,748,932 | 3,904,855 | 3,767,717 | 3,865,738 | 3,844,544 | 3,828,269 | 3,664,686 |
| No. of contigs | 62 | 166 | 32 | 44 | 126 | 35 | 149 | 47 | 47 |
| No. of plasmids | 2 | 0 | 1 | 1 | 1 | 1 | 2 | 1 | 2 |
| GC content (%) | 38.91 | 39.04 | 38.97 | 38.87 | 38.95 | 38.75 | 38.99 | 38.81 | 38.78 |
| N50 | 478,783 | 67,766 | 458,370 | 254,121 | 77,219 | 477,098 | 73,853 | 369,580 | 195,600 |
| N90 | 68,035 | 15,343 | 166,877 | 88,568 | 17,967 | 165,195 | 17,420 | 116,120 | 67,830 |
| L50 | 3 | 17 | 3 | 6 | 16 | 2 | 14 | 3 | 7 |
| L90 | 11 | 61 | 8 | 18 | 49 | 8 | 53 | 10 | 20 |
| No. of coding sequences | 3579 | 3428 | 3497 | 3661 | 3582 | 3573 | 3674 | 3576 | 3490 |
| MLSTOxf | ST3542* | ST3543* | ST3415 | ST3541* | ST871 | ST3544* | ST3545* | ST3540* | ST585 |
| MLSTPas | ST2832* | ST2114 | ST2522 | ST2834* | ST2700 | ST470 | ST2635 | ST2833* | ST10 |
| K loci | KL121 | KL96 | KL170# | KL202# | KL112 | KL172# | KL81 | KL183# | KL108 |
| OC loci | OCL2 | OCL13 | OCL2 | OCL4 | OCL6 | OCL4 | OCL6 | OCL2 | OCL2 |
| Antimicrobial resistance genes/phenotype |
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 19053 | 19055 | 19056 | 19058 | 19060 | 19061 | 19062 | 19063 | 19064 | |
| Beta‐lactams:
Resistance gene(s) Meropenem (10 µg) |
|
|
|
|
|
|
|
|
|
| 22.3 (S) | 26.6 (S) | 24.2 (S) | 29.1 (S) | 28.6 (S) | 25.7 (S) | 21.0 (S) | 27.0 (S) | 24.6 (S) | |
| Aminoglycoside resistance gene(s) |
|
| — | — |
| — |
| — |
|
| Tetracyclines:
Resistance gene(s) Tetracycline Doxycycline |
| — | — | — | — | — |
| — | — |
| 64.0 (R) | 12.0 (S) | 4.0 (S) | 6.0 (S) | 6.0 (S) | 4.0 (S) | 128.0 (R) | 4.0 (S) | 6.0 (S) | |
| 4.0 (S) | 4.0 (S) | 4.0 (S) | 4.0 (S) | 4.0 (S) | 4.0 (S) | 4.0 (S) | 4.0 (S) | 4.0 (S) | |
| Sulfonamide resistance genes(s) | — | — | — | — | — | — |
| — | — |
| Fluroquinolones:
Mutation(s) in quinolone resistance‐determining regions of Ciprofloxacin (5 µg) | — | — | — | — | — | — | — | — | — |
| 23.5 (S) | 25.2 (S) | 24.7 (S) | 28.3 (S) | 28.0 (S) | 28.0 (S) | 21.7 (S) | 26.8 (S) | 24.8 (S) | |
| Efflux pumps |
|
|
|
|
|
|
|
|
|
- —This study was funded by a Newton Fund Institutional Links award to S.C.C. (Grant No. 172686537), and two University of Southampton HEFCE Newton Fund Official Development Assistance (ODA) awards, one
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Vibrio bacteria research studies · Salmonella and Campylobacter epidemiology
Introduction
1
Carbapenem‐resistant Acinetobacter baumannii is recognized by the World Health Organization (WHO) as the top critical priority pathogen, posing the highest threat to public health due to limited treatment options (World Health Organization 2024). Unsurprisingly, the majority of reported A. baumannii genomes and those deposited in public databases, such as NCBI Genomes and the PubMLST genome collection, are mainly of hospital origin. These isolates are obtained from patients with hospital‐acquired infections, such as ventilator‐associated pneumonia, meningitis, blood stream infections, and urinary tract infections (Bian et al. 2021; Cui et al. 2023; Shelenkov et al. 2024). As a result, the public A. baumannii genome datasets are heavily skewed toward dominant hospital‐associated clones, notably members of the notorious Global Clone 2 (GC2) lineage (Shelenkov et al. 2023). While keeping track of hospital‐related A. baumannii is essential due to their formidable antimicrobial resistance (AMR) and clinical relevance, they only represent a subset of the species. These strains often differ significantly from those found in the normal flora of healthy populations (Muzahid et al. 2023). A. baumannii from healthy communities remains largely understudied, more so for indigenous communities that are isolated from urban populations.
The Orang Asli are indigenous people in Peninsular Malaysia comprising several ethnic subgroups who retained their aboriginal language, customs and lifestyle (Mahmud et al. 2022). They are only a minor population in Malaysia (0.8% of the population in Peninsular Malaysia based on the year 2020 census) and often fall behind national socioeconomic, education and healthcare improvement plans. Despite government resettlement programs, many Orang Asli communities remain in rural areas due to their lifestyle preferences (Pah Rokiah Syed Hussain et al. 2017; Mahmud et al. 2022). Some of these isolated communities are difficult to reach, limiting their access to modern medicines, such as antibiotics and vaccines (Mohd Rosman et al. 2020; Chew et al. 2022). This restricted interaction with urban communities and healthcare has led to the formation of what we term here as “genomic capsules”—distinct microflora genomes unique to the Orang Asli community and their respective tribes. As a result, opportunistic pathogens such as A. baumannii harbored by the Orang Asli may differ from strains commonly present in urban hospitals.
A. baumannii from hospitals have been well‐studied over the past two decades, gaining their notoriety due to their multidrug resistance (MDR), extensive drug resistance, and pan‐drug resistance (PDR) characteristics (Shi et al. 2024). The Malaysian Ministry of Health has published annual National Surveillance of Antibiotic Resistance (NSAR) Reports since 2003. Beginning in the 2010s, more than 50% of A. baumannii isolates have been reported to be resistant to carbapenems (i.e., imipenem and meropenem), which are the drugs of choice for treatment. However, the NSAR data set is limited to participating hospitals, which are mainly in the urban and suburban areas of Malaysia (Ministry of Health 2023). While the WHO's Tracking AMR Country Self‐Assessment Survey (TrACSS) Country Report emphasizes the importance of addressing AMR at the community level to enhance infection prevention and control efforts (WHO 2022), there remains a significant knowledge gap regarding AMR profiles and genomic characteristics of bacterial pathogens within the indigenous communities in Malaysia. Notably, a recent study investigating A. baumannii isolates from human fecal samples in a community in Segamat, Malaysia, revealed phylogenomic clustering of four community‐derived strains with two isolates from the town's main tertiary hospital. This finding suggests the potential persistence and circulation of certain A. baumannii strains across both community and healthcare settings (Muzahid et al. 2023). In this study, we aim to provide a genomic snapshot of A. baumannii that were isolated during an all‐age, upper respiratory tract microbial carriage study undertaken among two rural Orang Asli communities in the state of Terengganu, located on the eastern coast of Peninsular Malaysia in 2017. A previous investigation of Klebsiella pneumoniae isolates from a broader indigenous cohort revealed the predominance of ST23, which is commonly associated with clinical K. pneumoniae infections, and of concern, a proportion of these isolates harbored genes that categorized them as hypervirulent (Das et al. 2024). Here, we present the genomic analysis of A. baumannii isolates recovered from the upper respiratory tract of the Orang Asli and show the genetic diversity of this hitherto unexplored A. baumannii “genomic capsule.” Our findings offer insights into strains of novel sequence types (STs), their patchwork of unique and shared mobile genetic elements, AMR, and virulence genes.
Materials and Methods
2
Sampling and Isolation of A. baumannii From Orang Asli Communities
2.1
Swabs were taken from two Orang Asli villages, namely, Kampung Sungai Pergam and Kampung Berua, in the state of Terengganu on the east coast of Peninsular Malaysia. Nasal swabs and nasopharyngeal swabs were taken from each participant, as described (Cleary et al. 2021). Conventional bacteriology of the samples were carried out using Columbia Blood Agar (CBA), CBA with chocolated horse blood agar, CBA with colistin and naladixic acid agar, CBA with chocolated horse blood and Bacitracin, and Lysed Gonococcal (GC) selective agar (all culture media from Oxoid, UK) (Cleary et al. 2021). Preliminary identification of presumptive Acinetobacter spp. isolates was done using the MALDI Biotyper (Bruker, UK) at the Portsmouth Microbiology Laboratories, UK.
Antibiotic Susceptibility Tests
2.2
A. baumannii was spread over Mueller‐Hinton agar plates (MH, Oxoid, UK). Susceptibilities to the antibiotics meropenem and ciprofloxacin were determined by disk diffusion using the appropriate antibiotic disk (meropenem, 10 µg; ciprofloxacin, 5 µg) (Oxoid, UK), whereas tetracycline and doxycycline susceptibilities were determined by placing minimum inhibitory concentration (MIC) E test strips (bioMérieux, France) onto the surface of the agar. All agar plates were incubated at 35°C ± 1°C for 18 ± 2 h. Susceptibility was determined against the EUCAST Clinical Breakpoint guidelines (2024).
Genome Sequencing and Assemblies
2.3
Genomic DNA of the nine A. baumannii isolates was extracted using the QIAmp DNA Mini extraction kit (Qiagen, UK) per the manufacturer's instructions. Concentration of genomic DNA was determined using a Qubit 2.0 fluorometer (Thermo‐Fisher, UK). Whole genome sequencing was performed on a MiSeq (Illumina, UK) short‐read platform at a commercial sequencing provider (MicrobesNG, UK) using the 500‐cycle v2 reagent kit to generate 2 × 150 bp paired‐end reads. Raw reads obtained were then quality assessed and trimmed using fastp (available from https://github.com/OpenGene/fastp; Chen 2023). Genome assemblies were carried out using Unicycler (available from https://github.com/rrwick/Unicycler; Wick et al. 2017), followed by evaluation using Quast (available from https://github.com/ablab/quast).
Bioinformatics Analyses
2.4
Average nucleotide identity (ANI) of the assembled genomes to reference genomes was determined using fastANI (available from https://github.com/ParBLiSS/FastANI; Jain et al. 2018). Annotation of the genomes was performed using Prokka (available from https://github.com/tseemann/prokka; Seemann 2014). Conventional multilocus sequence typing (MLST) profiles of the assembled genomes were determined using mlst (available from https://github.com/tseemann/mlst) and matched to the PubMLST database (https://pubmlst.org/organisms/acinetobacter-baumannii). MLST profiles determined from the two available schemes, namely, the Oxford and Pasteur schemes, were used to identify the corresponding Global Clones (GCs). Serotyping based on A. baumannii surface polysaccharide loci, namely, capsule K loci (KL) and lipo‐oligosaccharide OC loci (OCL), was carried out using Kaptive v3.0.0b6 (available from https://github.com/klebgenomics/Kaptive; Wyres et al. 2020; Cahill et al. 2022).
Genotypic resistance profiles of the genomes were determined using AMRFinderPlus (available from https://github.com/ncbi/amr; Feldgarden et al. 2021) and ABRicate (available from https://github.com/tseemann/abricate), whereby databases from CARD (Alcock et al. 2023) and ResFinder (Zankari et al. 2012) were utilized for the latter approach. Virulome of the assembled genomes was determined using ABRicate, utilizing database from Virulence Factor DataBase (Liu et al. 2022). Findings were then compared with the results obtained from VFAnalyzer (available from https://www.mgc.ac.cn/cgi-bin/VFs/v5/main.cgi). Mobile genetic elements such as plasmids, insertion sequence (IS) elements, and resistance island (RI) hotspots were also determined from the genomes. Plasmids were identified using PlasmidFinder (available from https://github.com/genomicepidemiology/plasmidfinder; Carattoli et al. 2014), whereas classification of the plasmid replication protein (Rep) was performed using an in‐house built script, pREPonly (https://github.com/lean-SS/pREP-only), utilizing the AcinetobacterPlasmidTyping database (available from https://github.com/MehradHamidian/AcinetobacterPlasmidTyping; Lam et al. 2023). pdif sites in the plasmids found were identified using a combination of pdif finder (https://github.com/mjshao06/pdifFinder) (Shao et al. 2023) and manual search as outlined by Ambrose and Hall (2024). Toxin‐antitoxin (TA) systems were determined using the TADB 3.0 database (https://bioinfo-mml.sjtu.edu.cn/TADB3/index.php) (Guan et al. 2024). IS elements were screened using ISEScan (available from https://github.com/xiezhq/ISEScan; Xie and Tang 2017) and ISfinder‐sequences database through the Prokka —protein option (available from https://github.com/thanhleviet/Isfinder-sequences; Siguier 2006), whereas the comM RI hotspot (Hamidian and Hall 2017) was determined through local BLAST.
Pangenome and Phylogenetic Analysis
2.5
The pangenomes of the nine Orang Asli A. baumannii isolates were compared with other community A. baumannii genomes using the Anvi'o platform (available from https://github.com/merenlab/anvio; Delmont and Eren 2018; Eren et al. 2021). Through the use of the anvi‐pan‐genome program, the pangenomes were determined and then visualized through anvi‐display‐pan, which links to the Anvi'o server. Core genome phylogenetic analysis of A. baumannii genomes was performed using Roary (available from https://sanger-pathogens.github.io/Roary/; Page et al. 2015) and VeryFastTree with the GTR+CAT model, which combines the General Time Reversible (GTR) nucleotide substitution model with a Constant Rate Across Sites (CAT) approximation (Piñeiro et al. 2020; Piñeiro and Pichel 2024), and visualized with iTOL v7 (Letunic and Bork 2024).
Results and Discussions
3
Preliminary Genomic Analysis of A. baumannii From the Orang Asli
3.1
A total of thirteen presumptive Acinetobacter spp. isolates were obtained from the Orang Asli carriage studies at Kampung Sungai Pergam (n = 3) and Kampung Berua (n = 10). These were from the nasopharyngeal swabs of a total of 130 participants (of which 68 were from Kampung Sungai Pergam and 62 from Kampung Berua) (Cleary et al. 2021). Whole genome sequencing was performed on all thirteen isolates, out of which nine were shown to be A. baumannii (Table 1). The remaining four isolates were determined to be A. nosocomialis.
Genome assemblies of the nine A. baumannii Orang Asli isolates showed total genome sizes that ranged from ~3.7 to 3.9 Mbp (Table 1). ANI of the genomes revealed > 97% nucleotide identities to A. baumannii genomes, including those from Thailand (SAMEA104305267, SAMEA104305309, SAMEA104305313, and SAMEA104305269), Vietnam (5577STDY7716391, 5577STDY7716201, and 5577STDY7716392), and Malaysia (SAMN03174920 and SAMN03174917), as listed in Table 1. In silico epidemiological typing of the assembled genomes, which includes traditional MLST and surface polysaccharide loci typing, showed that each genome is distinct with its own ST, KL, and OCL types. MLST profiles based on the Oxford scheme revealed six new STs, with only A. baumannii 19056, 19060, and 19064 that were identified as preexisting ST3415_Oxf_, ST871_Oxf_, and ST585_Oxf_, respectively (Table 1). The new Oxford STs were submitted and assigned by the PubMLST curators as ST3540_Oxf_, ST3541_Oxf_, ST3542_Oxf_, ST3543_Oxf_, ST3543_Oxf_, ST3544_Oxf_, and ST3545_Oxf_ (Table 1). Additionally, the Pasteur MLST scheme identified three novel STs from the nine A. baumannii genomes, and these were assigned as ST2832_Pas_, ST2833_Pas_, and ST2844_Pas_. The six remaining genomes each belonged to different preexisting Pasteur STs (Table 1). The identification of new STs suggests the possible emergence of genetically distinct lineages within the Orang Asli community. The A. baumannii isolates with the new STs, although currently mostly susceptible, are capable of acquiring new resistance and virulence genes. Their presence in a community setting is noteworthy as they could represent a new lineage with the potential to spread and evolve, similar to how the hospital‐associated GCs, such as GC2, have emerged (Hamidian and Nigro 2019).
A. baumannii surface polysaccharide loci typing based on the KL, which were responsible for the production of acinetamic acid (Lam et al. 2022), showed that each of the nine Orang Asli A. baumannii genomes belonged to distinct KL types (Table 1). Notably, four novel capsule types (i.e., KL170, KL202, KL172, and KL183) were detected, indicating previously unreported characteristics and underscoring the uniqueness of these Orang Asli A. baumannii isolates. In contrast, analysis of the outer core lipo‐oligosaccharide loci (OCL) showed OCL2 (n = 4) to be the most common type, followed by OCL4 (n = 2) and OCL6 (n = 2) (Table 1). Nevertheless, A. baumannii 19055 was identified as the lesser‐studied OCL13 type, which was originally described in A. baumannii strains associated with community‐acquired pneumonia in the Northern Territories, Australia (Meumann et al. 2019). The A. baumannii capsule plays a crucial role in its ability to cause disease, primarily by protecting the bacterium from the host immune system and desiccation (Rakovitsky et al. 2021; Talyansky et al. 2021). Novel capsule types could therefore have a significant impact on the bacteria's survival and virulence.
Genotypic AMR Observation Revealed Nonmainstream β‐Lactamases
3.2
The presence of genes encoding β‐lactamases (bla) has become the hallmark of A. baumannii, not only as key determinants of carbapenem resistance (Shi et al. 2024), but also through the intrinsic bla OXA‐51‐like genes, which have been adopted as one of the typing methods for A. baumannii over the past two decades (Shelenkov et al. 2023). The recent categorization of GCs also incorporated various bla genes as characteristic traits observed in certain GCs, for example, members of the GC8 lineage commonly harbor both bla OXA‐23 and bla OXA‐68 (Shelenkov et al. 2023). Although bla genes, such as bla OXA‐23, bla OXA‐51, and bla OXA‐66, are widespread among A. baumannii globally (Li et al. 2023; Shelenkov et al. 2023; Shi et al. 2024), a different spectrum of bla genes was found from the Orang Asli A. baumannii in this study (Table 2).
The bla OXA‐51 family (or bla OXA‐51‐like) is comprised of the well‐characterized bla OXA‐51 gene and its numerous variants, such as bla OXA‐64, bla OXA‐66, and others (Li et al. 2023). In this study, several variants were detected in the Orang Asli A. baumannii genomes, including bla OXA‐68, bla OXA‐98, bla OXA‐120, bla OXA‐377, bla OXA‐424, bla OXA‐510, and bla OXA‐555. Only two genomes (i.e., A. baumannii 19058 and 19061) carried bla OXA‐51 itself, while the remaining seven harbored distinct variants of the bla OXA‐51 family (Table 2). The spectrum of genes of the bla OXA‐51 family aligns with the findings of Muzahid et al. (2023), who reported that community‐derived A. baumannii isolates from Segamat, Peninsular Malaysia, also carried a wide range of bla OXA‐51 variants. In the Segamat A. baumannii isolates, bla OXA‐120 was the most prevalent variant, followed by bla OXA‐441, bla OXA‐510, bla OXA‐69, bla OXA‐98, and bla OXA‐412. None of these variants were found among the A. baumannii isolates from the Orang Asli community.
However, the presence of the intrinsic bla OXA‐51 family of genes is not considered a definite marker for carbapenem resistance in A. baumannii due to the low affinities of the OXA‐51 family of β‐lactamases to meropenem and imipenem, as well as the very low expression levels of the bla OXA‐51‐like genes. In certain cases, the presence of ISAba1 directly upstream of the bla OXA‐51‐like gene provides a strong promoter which increases its expression level, leading to carbapenem resistance but this also depends on the bla OXA‐51 variant that is being overexpressed (Nigro and Hall 2018). All nine Orang Asli A. baumannii isolates were phenotypically carbapenem susceptible, and none of their genomes contained ISAba1 (or related elements) upstream of the bla OXA‐51‐like genes. None of the nine Orang Asli A. baumannii isolates also harbored acquired bla OXA‐encoded carbapenemase genes, such as bla OXA‐23, bla OXA‐24, and bla OXA‐58, which have been directly implicated in carbapenem resistance, particularly in clinical A. baumannii isolates (Hamidian and Nigro 2019). This was similarly reported by Muzahid et al. (2023), where the acquired carbapenemase gene bla OXA‐23 was only identified in their Segamat hospital isolates, which are carbapenem resistant. Likewise, our recent study of a 10‐year collection of A. baumannii from the main tertiary hospital in Terengganu also revealed the predominance of the bla OXA‐23 gene among the carbapenem‐resistant isolates (Din et al. 2025).
Another class of intrinsic β‐lactamase found in A. baumannii genomes is the AmpC cephalosporinase variants, which are designated Acinetobacter‐derived cephalosporinases (ADCs). Overproduction of ADCs resulting from insertion of ISAba1 or similar IS elements upstream of the bla ADC gene has been shown to be responsible for the development of resistance toward extended‐spectrum cephalosporins and, in some cases, carbapenems (Tian et al. 2011; Bhattacharya et al. 2014; Shi et al. 2024). Seven different variants of ADCs were detected from the genomes of the nine Orang Asli A. baumannii isolates (Table 2), and, in all cases, ISAba1 or similar elements were absent upstream of the encoding gene, suggesting that these genes were either not expressed or were expressed at low levels in their hosts. ADC‐25 was identified in two of the Orang Asli A. baumannii isolates (i.e., 19053 and 19061; Table 2), and this variant was found to be the seventh most prevalent ADC variant among A. baumannii isolates globally (Mack et al. 2025). Four of the eight ADC variants identified here (i.e., ADC‐99 in A. baumannii 19062, ADC‐238 in 19060, ADC‐279 in 19055, and ADC‐312 in 19058) were listed by Mack et al. (2025) as variants that were rarely found in A. baumannii. By comparison, the A. baumannii isolates from the Segamat community also presented a different spectrum of bla ADC genes, where they mainly harbored bla ADC‐154 and bla ADC‐156 (Muzahid et al. 2023), but both variants were absent in our Orang Asli isolates. However, bla ADC‐238, which was found in A. baumannii C‐65 from the Segamat community, was also found in A. baumannii 19060 from our Orang Asli collection. The ADC‐238 variant was listed as a less‐frequently encountered variant (Mack et al. 2025), and its singular presence in both these community‐based studies supports this finding. Intriguingly, hospital isolates from Segamat (Muzahid et al. 2023) and Terengganu (Din et al. 2025) showed a uniform pattern of bla ADC‐73 being the most prevalent, agreeing with the analysis presented by Mack et al. (2025), which revealed bla ADC‐73 as the most prevalent ADC variant in A. baumannii isolates globally, with the exception of isolates from North America.
Hence, in terms of the class C (ADC) and class D (OXA) β‐lactamases, the intrinsic variants harbored by the A. baumannii community isolates showed diversity with little in common, although both studies (i.e., this study and the Segamat study) were in Peninsular Malaysia. Even between communities, the AMR profiles vary (Meumann et al. 2019; Muzahid et al. 2023), and thus, there is more to be learned about A. baumannii from the Orang Asli community, which is further elaborated in the following sections.
Other Resistance Genes in the Orang Asli Community A. baumannii Isolates
3.3
Two out of the nine Orang Asli A. baumannii isolates (i.e., 19053 and 19062) were found to harbor the tet(39) tetracycline resistance gene (Table 2), which encodes a tetracycline efflux pump of the major facilitator superfamily (Agersø and Guardabassi 2005). This differs from the A. baumannii community isolates from Segamat, in which no tetracycline resistance genes were detected. Both A. baumannii 19053 and 19062 were phenotypically tetracycline resistant (with MIC values of 64 and 128 µg/mL, respectively) but doxycycline susceptible (both with MIC values of 4 µg/mL). In contrast, Malaysian A. baumannii hospital isolates predominantly carried the tet(B) gene (n = 60/126 from Hospital Sultanah Nur Zahirah (HSNZ) (Din et al. 2025); n = 12/15 from Segamat Hospital (Muzahid et al. 2023)), with only one isolate from HSNZ harboring tet(A) and 10/126 carrying tet(39) (Din et al. 2025). All A. baumannii hospital isolates harboring the tet(39) and tet(A) genes were tetracycline resistant and minocycline susceptible, whereas those that harbored the tet(B) gene were mostly resistant to tetracycline but showed intermediate susceptibility to minocycline (Din et al. 2025). Meumann et al. (2019) reported two A. baumannii isolates from community‐onset pneumonia in Australia, which harbored both the tet(B) and tet(39) genes, but phenotypic susceptibility testing for tetracyclines was not performed in their study. The two tetracycline‐resistant Orang Asli A. baumannii isolates, 19053 and 19062, harbored the tet(39) gene on plasmids, which will be elaborated in a later section.
Resistance to aminoglycosides in A. baumannii is mainly mediated by the possession of genes encoding aminoglycoside acetyltransferase (aac), nucleotidyltransferase (ant), and/or phosphotransferase (aph) (Shi et al. 2024). Five out of the nine A. baumannii Orang Asli isolates carried the ant(3″)‐IIa gene (Table 2), whereas Muzahid et al. (2023) reported the presence of this gene in all 12 of their A. baumannii strains that were isolates from the community in the town of Segamat. Nevertheless, only two of the 12 Segamat community isolates showed resistance to amikacin, and all were gentamicin susceptible, suggesting that some of these aminoglycoside resistance genes were either not expressed or expressed at a very low level. Phenotypic resistance to aminoglycosides was, however, not tested for the Orang Asli A. baumannii isolates in this study.
Sulfonamide resistance in A. baumannii is usually mediated by sul1 and/or sul2 genes (Sköld 2001), with sul2 predominantly reported from Southeast Asian countries and the Asia‐Pacific region (Bian et al. 2021; Brito et al. 2022; Din et al. 2025). Only one Orang Asli isolate, A. baumannii 19062, was found to harbor the sul2 gene (Table 2), which was absent in the Segamat community isolates (Muzahid et al. 2023). In contrast, nearly 50% (61/126) of the A. baumannii hospital isolates from HSNZ, Terengganu, harbored the sul2 gene (Din et al. 2025), whereas in Hospital Segamat, the gene was identified in 2/15 of the A. baumannii isolates. The significance of the carriage of the sul2 gene in the solitary Orang Asli A. baumannii isolate is currently unknown, but sul2 is known to be present on mobile elements, such as plasmids and transposons (Jeon et al. 2023). Plasmid analysis appeared to rule out the carriage of sul2 in either of the two plasmids found in A. baumannii 19062 (see subsequent Section 3.7), but this does not rule out its location on other mobile elements such as transposons or genomic islands in the chromosome.
The Virulome of A. baumannii From the Orang Asli Community
3.4
The isolation of A. baumannii from diverse sources, including clinical settings, soil, and wastewater, highlights its ability to persist across various environmental niches, facilitating its widespread dissemination, colonization, and pathogenicity (Harding et al. 2018). This persistence, evidenced by stress resistance and biofilm formation, is likely supported by the acquisition and/or inheritance of multiple virulence factors (VFs). In this study, the majority of identified VFs are associated with adhesion, biofilm formation, and quorum‐sensing regulation (Figure 1). These findings are in agreement with Muzahid et al. (2023), who reported similar virulome profiles in A. baumannii isolates from the Segamat community, including genes related to adhesion (e.g., ompA, fliP, pilA, and pilE), biofilm formation (e.g., adeFGH, bap, csuA/B, csuABCDE, and pgaABCD), and quorum sensing (e.g., abaI and abaR) (Choi et al. 2009; Lannan et al. 2016; Ahmad et al. 2023). However, slight variations in VF combinations were observed between these two Malaysian community studies. Notably, the biofilm gene combination we termed BIO‐Profile 1 (Figure 1) was dominant in our isolates but was absent in those from the Segamat community. Despite these differences, the presence of numerous shared VFs underscores their potential role in supporting the persistence of the A. baumannii isolates in their niche and their capacity to cause infection.
Profiles of various virulence factors (VFs) identified from the Orang Asli Acinetobacter baumannii genomes (n = 9). The profiles were abbreviated according to their biological functions: adhesion (ADH), antiphagocytosis (APC), biofilm formation (BIO), phospholipase enzyme (ENZ), immune evasion (IMM), iron uptake (IRO), regulation (REG), and serum resistance (SRM); singleton genes were labeled as is.
Survival of A. baumannii in harsh environmental conditions requires mechanisms for the acquisition of micronutrients, such as iron scavenging through the production of acinetobactin in iron‐limiting environments (Lannan et al. 2016; Harding et al. 2018). The presence of iron uptake genes is thus a signature VF in A. baumannii and the combination of iron acquisition genes designated IRO‐Profile2 (Figure 1) was also observed in the Segamat community isolates along with isolates from the Australian Northern Territory community (Meumann et al. 2019; Muzahid et al. 2023).
All nine A. baumannii isolates displayed the full complement of the lps‐lpx genes (designated IMM‐Profile; Figure 1), which are tagged as VFs that function in immune evasion. Deficiency in lpxC has been shown to cause the loss of the LPS layer in A. baumannii, leading to the development of colistin resistance (Kamoshida et al. 2020). The full suite of the lps‐lpx genes was found in the Segamat A. baumannii isolates, and this included hospital isolates that were identified as resistant to colistin (n = 2) and polymyxin B (n = 4); nevertheless, a more detailed analysis of possible mechanisms for polymyxin resistance, including mutations in the lps‐lpx genes, was not presented (Muzahid et al. 2023).
The Type‐6 Secretion System (T6SS) is utilized by A. baumannii to release toxic effector proteins into the neighboring environment, offering a competitive advantage to the pathogen in multispecies environments (Carruthers et al. 2013) and also allowing A. baumannii to spread, invade, and resist host immune responses (Shadan et al. 2023). The T6SS main cluster (T6MC), which encompasses the tssA, tssB, tssC, tssD/hcp, tssE, tssF, tssG, tssH/clpV, tssK, tssL, tagX, vgrG, and PAAR genes (Fitzsimons et al. 2018; Lewis et al. 2019), was identified in five of the nine Orang Asli A. baumannii isolates (Figure 1). These genes are responsible for T6SS apparatus assembly, whereby TssA functions as the priming protein (also known as the cap), TssBC forms the sheath, TssD/Hcp the secretion tube with VgrG and PAAR proteins as the spike. The spike (i.e., VgrG and PAAR) teams with the wedge (i.e., TssK and TssEFG) to form the baseplate (Fitzsimons et al. 2018; Marazzato et al. 2022). The structure was then supported by the membrane complex formed by TssJ, TssM, and TssL proteins connecting between inner and outer membranes (Fitzsimons et al. 2018; Marazzato et al. 2022). The T6SS found in the five A. baumannii genomes (Figure 1) was further identified as T6SS‐1A (i.e., 19053, 19056, and 19063) and T6SS‐1B (i.e., 19058 and 19061), according to the classification of Repizo et al. (2019). These five A. baumannii isolates also encode the Tse4 effector (Figure 1), which function as an amidase (Lewis et al. 2019; Repizo et al. 2019). Other effectors were also present in the Orang Asli isolates, with A. baumannii 19063 and 19053 encoding an additional Tse2 (predicted to function as a DNase), while A. baumannii 19053 also encodes additional Tse1 (predicted lipase producer), and Tse3 (effector of unknown function) (Lewis et al. 2019; Repizo et al. 2019). Conversely, only one of the Segamat community isolates, A. baumannii C‐98, harbored the complete T6SS, whereas the hospital isolates contained the full suite of T6SS genes (Muzahid et al. 2023). The majority of the A. baumannii hospital isolates from Terengganu also harbored the full T6MC (n = 94/126; or 74.6%) (Din et al. 2025). This suggests that the Orang Asli A. baumannii isolates may be better adapted to survival in a multispecies environment with also the capacity to invade and colonize their host, should the opportunity arise; however, such possibilities would require further experimental validation.
The Orang Asli A. baumannii Isolates Were Genetically Diverse
3.5
Community isolates of A. baumannii represent a pool of unexplored genomes when compared with the more well‐studied clinical isolates. Therefore, many A. baumannii isolates presented in community studies belonged to novel STs that have yet to be classified into any clonal complexes. We pooled together the A. baumannii genomes obtained from the Orang Asli in this study, along with the genomes obtained from fecal samples of the community in the town of Segamat (Muzahid et al. 2023), for pangenome analysis. The analysis showed genetic diversity among these community isolates from the 5277 cloud genes identified (53.84%) when compared with the 2273 core genes (23.19%) (Figure 2). The lower ratio of core genes indicated low homogeneity between the A. baumannii community isolates, highlighting the uniqueness of the bacterium in each population. Even within the Segamat population itself, the higher diversity of the community A. baumannii isolates was apparent, as compared with the hospital isolates from the same town (Muzahid et al. 2023). Of interest, there did not seem to be any apparent geographical clustering between the two populations (Figure 2), and this was evident when examining the core genome phylogenetic tree that was generated using both community and clinical isolates of A. baumannii from Malaysia (Figure 3). Segamat is a township in the state of Johor and is approximately 390 km south of the state of Terengganu, where the sampling was carried out in the Orang Asli rural settlements.
Pangenome analysis of community Acinetobacter baumannii isolates from Malaysia that are currently available in the databases (n = 20). The orange‐colored tracks represent the Orang Asli community A. baumannii from this study (n = 9), whereas the green‐colored tracks represent the Segamat community strains (n = 11; isolates with the prefix “C”) that were previously published (Muzahid et al. 2023). Heatmap on the top right corner presents the average nucleotide identity (ANI) of A. baumannii from both communities, with all of them having > 97% identity. GC, Global Clone.
Midpoint‐rooted maximum‐likelihood phylogenetic tree of all Malaysian Acinetobacter baumannii genomes that are published (i.e., the current Orang Asli isolates, the Segamat hospital and community isolates described by Muzahid et al. 2023, and the 10‐year HSNZ isolates reported by Din et al. 2025), along with A. baumannii genomes originating from Malaysia that are found in PubMLST (as of January 30, 2025). Also shown are their categorization into the various Global Clone (GC) lineages, the presence/absence of predominant carbapenemase (purple boxes) and tetracycline resistance genes (green boxes), the intactness of the comM gene (orange circles), and the presence of plasmids of the Rep_3 family (light green tick marks). HSNZ, Hospital Sultanah Nur Zahirah.
A maximum‐likelihood phylogenetic tree was generated from the core genome alignment of 199 Malaysian A. baumannii genomes (Figure 3), and these included the genomes from this study, the Segamat study (both community and hospital isolates) (Muzahid et al. 2023), 126 genomes from a 10‐year collection of isolates from HSNZ, the main tertiary hospital in Terengganu (Din et al. 2025), and other clinical isolates of A. baumannii obtained from the PubMLST database (refer to Supporting Information Appendix 1 for the list of genomes). Hospital isolates of A. baumannii were predominantly ST2_Pas_, which were categorized under the GC2 lineage, and this is clearly evident in the phylogenetic tree where they are clustered in a distinct clade (Figure 3). GC2 is the predominant A. baumannii lineage globally (Nigro and Hall 2018), but the basis of this predominance is that the overwhelming majority of sequenced isolates were from hospitals (Shelenkov et al. 2023). As mentioned earlier, the Orang Asli A. baumannii genomes were scattered throughout the phylogenetic tree, as were the Segamat community isolates, whereas most of the Segamat hospital isolates were clustered together in the GC2 clade, much like the HSNZ isolates. Interestingly, one Orang Asli isolate, A. baumannii 19064 (ST10_Pas_; ST585_Oxf_), was found to be a member of the GC8 lineage. Similarly, Muzahid et al. (2023) had reported that one of their community isolates from Segamat was identified as a member of the GC1 lineage. Apart from these exceptional cases, none of the community A. baumannii isolates belonged to any GC clusters. Additionally, the Orang Asli A. baumannii was also distinct from the Segamat community isolates, with only A. baumannii 19062 distantly grouped with C‐72 (Figure 3). Although the majority of the Orang Asli isolates did not belong to any of the major GCs, we observed that they were interleaved with a few non‐GC clinical isolates from HSNZ in neighboring branches (Figure 3). It is possible that these community A. baumannii isolates were able to cause infections whenever the opportunity arises (i.e., potential pathogenicity), and thus, we see the relatively close genetic relationship between some of the Orang Asli isolates and the non‐GC hospital isolates. However, without specific infection data and proven clinical relevance, this remains speculative. Nevertheless, genomic data can inform future research on A. baumannii, particularly with broader longitudinal carriage studies, contact tracing, or case control analysis with clinical samples, and any transmission between the community and hospital isolates could be detected and proven.
Apart from the distinct clustering of various GCs (Figure 3), the Malaysian clinical A. baumannii isolates also presented a different catalog of dominant bla genes, which varies from the community A. baumannii resistome described earlier. The presence of bla TEM‐1 (Ambler Class A), bla NDM‐1 (Ambler Class B), bla ADC‐73 (bla ADC‐1‐like; Ambler Class C), bla OXA‐23, and bla OXA‐66 (Ambler Class D) was recorded from almost all the Malaysian GC2 genomes, and likewise for the tetracycline resistance gene tet(B) (Figure 3). One distinctive feature of A. baumannii hospital isolates is the presence of large AbaR RIs that were often inserted within the chromosomal comM gene (Meumann et al. 2019). The intactness of the comM gene was investigated for all the Malaysian A. baumannii genomes presented here. Not surprisingly, the comM gene was interrupted in all the GC2 isolates, whereas the proportion was 87.5% for GC1, 71.4% for GC7, and 66.7% for GC11 isolates (Figure 3). The non‐GC A. baumannii genomes have a much lower percentage of interrupted comM, and within the nine Orang Asli isolates, only A. baumannii 19060 was identified with disruption of the comM gene (Figure 3). Nevertheless, the genes that were inserted within comM in A. baumannii 19060 were not associated with AMR but rather those associated with metabolism, regulatory genes, and hypothetical proteins, much like what was described for the community‐onset isolates from the Australian Northern Territory (Meumann et al. 2019).
IS Elements
3.6
A total of 17 ISs belonging to five families, namely, IS3, IS5, IS91, IS630, and ISL3, were identified from the Orang Asli A. baumannii genomes (Supporting Information Appendix 2). ISAba43 of the ISL3 family (Cameranesi et al. 2020) was found in all nine A. baumannii genomes, with at least one copy of the IS in each genome (Supporting Information Appendix 2). Other IS families were found in fewer numbers with ISAba40, ISAba57, and ISAba63 of the IS3 family found in one genome each, and the remaining IS elements identified were found only in A. baumannii 19062 (Supporting Information Appendix 2). There were few reports for most of these IS elements except IS1006, which was identified in A. baumannii 19062 and is well‐known for its association with plasmid‐borne AMR regions (Harmer and Hall 2021; Varani et al. 2021; Hall 2022). However, the identified copy of IS1006 in A. baumanii 19062 was not found to be associated with any resistance genes.
Carriage of Plasmids in A. baumannii From the Orang Asli Community
3.7
Plasmids play an important role in the evolution of A. baumannii, being a major vehicle for the dissemination of antibiotic resistance genes (Lam and Hamidian 2024; Tobin et al. 2025). In a comprehensive survey of 439 mostly complete A. baumannii genomes, more than half (52%) contained one plasmid, 27% harbored two plasmids, while seven genomes contained 6–11 plasmids (Lam and Hamidian 2024). Plasmids of the Rep_3 family were by far, the most predominant with more than half of these plasmids harboring antibiotic resistance genes (Lam et al. 2023; Lam and Hamidian 2024; Tobin et al. 2025). Among the nine Orang Asli A. baumannii genomes, only one isolate, A. baumannii 19055, was without any detectable plasmids; three isolates (i.e., 19053, 19062, and 19064) harbored two plasmids each, while the remaining five isolates contained a plasmid each (Supporting Information Appendix 3). Eight of these plasmids were small plasmids (i.e., < 10 kb; Lean and Yeo 2017), ranging in size from 2178 to 8837 bp. Out of these eight small plasmids, only one plasmid, p19053a, belonged to the Rep_1 family, specifically the R1‐T6 type. p19053a was only 2178 bp and harbored the rep gene and two other hypothetical open reading frames (ORFs) (Supporting Information Appendix 4). Lam and Hamidian (2024) noted that the R1‐type plasmids are typically 2–3 kb in size and encode only the replication initiation protein along with one or two hypothetical proteins. None of these plasmids harbored AMR genes and p19053a follows the characteristics of the prototypical R1‐type plasmid.
The remaining seven small plasmids were of the Rep_3 family, specifically the R3‐T5 (n = 4), R3‐T13 (n = 2), and R3‐T64 (n = 1) types (Supporting Information Appendices 3 and 4). Two of these R3 types (i.e., R3‐T5 and R3‐T13) were reportedly among the most abundant R3 types globally, but their distributions were mainly linked to minor STs (Lam and Hamidian 2024). Plasmid p19064a from the R3‐T5 type was found in an ST10_Pas_ host (A. baumannii 19064), which was the only isolate from this study that belonged to one of the known GC lineages, GC8, but did not harbor antibiotic resistance genes. Among these small plasmids, only p19053b, which was also of the R3‐T5 type, harbored the tet(39) tetracycline resistance gene (Figure 4). In contrast, A. baumannii clinical isolates from Malaysia, particularly those of the GC2 lineage, prevalently harbored an 8731‐bp plasmid we initially designated pAC12a (Lean et al. 2014; Lean and Yeo 2017; Din et al. 2025) and which is identical to the pA1‐1 plasmid (accession no. CP010782) that is harbored in a GC1 isolate obtained in 1982. This plasmid was previously typed as GR2 in an older scheme (Bertini et al. 2010) but has since been typed as R3‐T1, and almost half of the members of this group were identical or nearly identical to pA1‐1 (Lam et al. 2023). This supports the long‐standing association of this plasmid with the GC1 and GC2 lineages, but isolates of other STs have also been found that harbor this plasmid, albeit at a much lower frequency (Lam et al. 2023; Din et al. 2025). A characteristic feature of this plasmid is the presence of mobile pdif modules containing the sel1, abkBA TA genes, and a gene encoding a TonB‐dependent receptor (Lean and Yeo 2017; Lam et al. 2023). Notably, this plasmid type was absent from all A. baumannii isolates obtained from the Orang Asli population and the Segamat community, suggesting a possible specific association with clinical strains.
Comparisons of the R3‐T5‐type plasmids found in the Orang Asli Acinetobacter baumannii isolates with similar R3‐T5‐type plasmids in the database curated by Lam et al. (2023). The needle and thread icon shown for the p19064a plasmid map indicated that the plasmid was a composite that was stitched together from two separate contigs. The pdif sites are marked with green and red crosses representing XerC/D and XerD/C recognition sites, respectively.
Lam and Hamidian (2024) reported that almost half of the R3‐type plasmids were not associated with AMR determinants. The genetic structures of the R3‐T5 and R3‐T13‐type plasmids identified in this study (n = 6; Figure 4 and Supporting Information Appendix 4) showed the absence of AMR genes from five of them, except for the tet(39)–tetR genes in p19053b. The tet(39)–tetR genes are located within a mobile pdif module usually found in Acinetobacter plasmids (Blackwell and Hall 2017; Lean and Yeo 2017). A pdif module typically comprises one or two related genes flanked by pdif sites, which resemble the chromosomal dif site involved in site‐specific recombination. These 28 bp pdif sites contain binding regions for the recombinases XerC and XerD, similar to the chromosomal dif site near the bacterial chromosome terminus. The term pdif was used to differentiate these plasmid‐associated sites from chromosomal dif sites (Blackwell and Hall 2017; Castillo et al. 2017; Balalovski and Grainge 2020). Three other pdif modules were uncovered from p19053b, and these carry a nucleotide‐binding protein‐encoding gene, higBA TA genes, and a sel1 gene along with a gene encoding the SMI1/KNR4‐family protein (Figure 4; Supporting Information Appendix 5). Interestingly, the gene arrangement of the higBA and sel1‐SMI1/KNR4‐family pdif modules was observed in almost all the R3‐T5‐type plasmids identified in this study and also extends to the R3‐T13‐type plasmids (i.e., p19058 and p19063) (Figures 4 and 5; Supporting Information Appendix 4).
Comparative linear map of R3‐T13‐type plasmids identified from the Orang Asli Acinetobacter baumannii with closely related R3‐T13‐type plasmids found in the plasmid database presented by Lam et al. (2023). Red–green crosses representing pdif sites were labeled as indicated in the previous Figure.
Two plasmids of the R3‐T5‐type, namely, p19056 and p19061, were nearly identical, with both at 7146 bp (Figure 4). They were closely related to the pD1279779 plasmid (accession no. CP003968.1), which was slightly larger at 7416 bp with an additional hypothetical protein found downstream of the SMI1/KNR4‐family gene (Figure 4). Plasmid p19064a also appeared to be closely related to pD1279779, but the p19064a sequence was actually stitched together from two separate contigs (Figure 4), and whether there are additional genes that are lost in between these two contigs is not known. Interestingly, the host for pD1279779 was A. baumannii D1279779, which was isolated from a community‐acquired bacteremia patient in Northern Australia and typed as ST267_Pas_/ST942_Oxf_ (Farrugia et al. 2013). On the basis of the collection of Acinetobacter plasmids curated by Lam et al. (2023), the R3‐T5‐type plasmids identified in this study were related to pPM194229‐3 and pDETAB6, besides pD1279779. The p19053b plasmid, which harbored the tet(39)–tetR genes, is thus the sole R3‐T5‐type plasmid, which encode AMR determinants. This plasmid is more closely related to pDETAB6, which harbored two hypothetical ORFs instead of the tet(39)–tetR genes in p19053b (Figure 4).
AMR genes are also a rarity among the R3‐T13‐type plasmids. A search in the Acinetobacter plasmid repository posted by Lam et al. (2023) led to the discovery of p29FS20‐2 (accession no. CP044521.1) as the sole carrier of tet(39)–tetR, and the aminoglycoside resistance genes aph(3″)‐Ib and aph(6)‐Id in this plasmid type. Two of the three R3‐T13‐type plasmids identified in this study, p19058 and p19063, were not associated with AMR determinants; however, p19062b was the only R3‐T13‐type plasmid that harbored the tet(39)–tetR gene pair (Figure 5). Similar to p19053b, the tet(39)–tetR genes in p19062b were also located within a pdif module, a feature that was also observed in p29FS20‐2 (Figure 5). Plasmids p19058 and p19063 were more closely related to another R3‐T13‐type plasmid, p6200‐9.327 kb, but this plasmid contained additional genes encoding sulfate permease (sulP) and a universal stress protein (usp) (Figure 5). One of the interesting observations of note regarding these plasmids is the almost universal presence of the rep_3‐orfX backbone, the higBA, and sel1‐SMI1/KRN4 family modular arrangement of genes, covering also the R3‐T5‐type plasmids (Figures 4 and 5). The sole exception to this is plasmid p19062b, where the sel1‐SMI1/KRN4 pdif module was absent, and the putative TA module (identified as relBE by TADB 3.0) was distantly related to the higBA module usually found in the other similar plasmid‐types (Figure 5).
The other plasmids identified in the Orang Asli A. baumannii genomes are p19062a of the R3‐T64 type, p19060 of R3‐T26, and p19064b of R3‐T27 type (Supporting Information Appendix 4). These plasmid types are rarely encountered when compared with the R3‐T5 and R3‐T13 types (Lam et al. 2023). None of these plasmids harbor AMR determinants.
Even though only nine A. baumannii genomes were investigated in this study, the diversity of plasmids found reflects the diversity reported for the genus Acinetobacter (Lam and Hamidian 2024). Given the limited direct clinical antibiotic exposure in the studied community, it is plausible that plasmid‐mediated resistance (in particular, tetracycline resistance) is maintained and disseminated through environmental and ecological interactions. Local environmental sources (such as soil and water contaminated with antimicrobial residues) and animals (including livestock and wildlife) can act as reservoirs of resistance plasmids and provide opportunities for gene exchange between commensal, environmental, and pathogenic bacteria. Such interactions may help explain the persistence and spread of resistance determinants in these isolates despite low human antibiotic usage (Davies and Davies 2010; Aminov 2009). This is particularly true for tetracycline resistance, in which the antibiotic has been extensively used for decades in both human and veterinary medicine as well as in agriculture and aquaculture. This historical use could have created an initial selective pressure which led to the persistence of the resistance gene(s) in the bacterial population (Aminov 2009). Even if the exposure to tetracycline is now limited, the fitness cost of carrying the resistance gene might be low, which enables them to persist by being integrated into mobile genetic elements such as pdif modules and plasmids that are easily transmitted between bacteria. The tet(39) pdif module along with pdif modules harboring carbapenem and macrolide resistance genes have indeed been reported in diverse Acinetobacter spp. isolated from aquatic environments in South Australia (Tobin et al. 2024).
Perspectives, Limitations, and Suggestions for Future Studies
3.8
By conducting genomic analysis of A. baumannii strains from a previously understudied population, our work addresses a key WHO priority in its Global Action Plan on AMR (WHO 2015). The identification of resistance genes and their prevalence in this unique community provides new surveillance data from a region where such information is limited. This is essential for informing and strengthening national and regional AMR strategies, thereby contributing to the global surveillance network. Our findings on the presence of antimicrobial‐resistant A. baumannii in asymptomatic carriers from a community setting also underscore the importance of community‐level surveillance and infection prevention. WHO recognizes that AMR is not just a hospital problem (WHO 2015). For A. baumannii, there has been scarce data on the genomic characteristics of community‐origin isolates worldwide, resulting in the overwhelming prevalence of hospital‐origin GC2 genomes in the public databases (Lam and Hamidian 2024; Shelenkov et al. 2023; Hamidian and Nigro 2019). Our study also highlights the need for targeted public health interventions and awareness campaigns to mitigate the spread of resistant bacteria in nonclinical settings. This information is vital for developing effective community‐based strategies to limit the transmission of AMR.
A major limitation in this study is that these A. baumannii genomes were sequenced using the Illumina short‐read platform, which does not allow for complete genome assembly. Therefore, some of the plasmid architectures presented here should be taken with caution, as we could not ascertain if there are genes or genetic elements that are lost in the assembly of the draft genomes. This limitation is particularly relevant for plasmid p19064a, which was assembled across two separate contigs. Nevertheless, the fact that some of these plasmids have similar counterparts from clinical A. baumannii isolates is of concern, as they could serve as vehicles for the dissemination of AMR genes. The discovery of the tet(39)–tetR gene pair within a mobile pdif module in two distinct plasmids, p19053b and p19062b, underlines this likelihood and highlights the importance of continued genomic surveillance, particularly among the community.
Another limitation of this study is the small number of A. baumannii isolates that were obtained and analyzed (n = 9), thus necessitating future broader longitudinal studies for a better representation of the population. We also phenotypically tested a limited number of antimicrobials in this study (i.e., meropenem, ciprofloxacin, tetracycline, and doxycycline). Hence, increasing the number of antibiotics by including classes such as the aminoglycosides would be useful for a more comprehensive phenotypic resistance profile, more so as the aminoglycoside resistance gene, ant(3″)‐IIa, was detected in five isolates. The expression of these resistance genes can be validated by using quantitative real‐time reverse transcriptase PCR (qRT‐PCR), particularly for isolates in which the resistance gene is detected but the isolate remains phenotypically susceptible. Future studies should also focus on functional assays to determine the role of the new STs and capsule types in biofilm formation, resistance to environmental stresses, and host immune evasion. This will provide a clearer understanding of how these novel types might influence the ability of the bacteria to cause disease and resist antibiotics in clinical settings.
Conclusions
4
A. baumannii and its AMR mechanisms have long been central in the global fight against superbugs, as understanding these traits is essential for improving treatment strategies. Over the decades, numerous reports have highlighted the alarming rise in carbapenem‐resistant, MDR, and even PDR A. baumannii strains in clinical settings (Akeda 2021; C. H. Chen et al. 2023), suggesting that treatment practices themselves may contribute to the development of resistance (De Blasiis et al. 2024). In contrast, A. baumannii isolates from remote communities, such as the indigenous Orang Asli population studied here, exhibited lower levels of antibiotic resistance. However, data on community‐derived isolates remain scarce when compared with clinical isolates (Lam and Hamidian 2024).
Despite their relative antibiotic susceptibility, these community‐associated A. baumannii isolates still harbor a range of VFs and mobile genetic elements, including plasmids and ISs (Meumann et al. 2019; Muzahid et al. 2023), features well‐documented in hospital‐derived strains. Notably, one isolate from this study belonged to the GC8 clinical lineage, while Muzahid et al. (2023) previously identified a GC1 isolate in a more urbanized community setting in Malaysia. Moreover, the phylogenetic interleaving of these community isolates with certain non‐GC hospital strains suggests that they possess the capacity to cause infections and acquire resistance traits, akin to their clinical counterparts. The presence of shared genomic features in A. baumannii from a presumptive antibiotic‐naïve environment underscores the pathogen's inherent capacity for persistence and colonization, even in healthy individuals. Of concern is the detection of two A. baumannii isolates from this study that harbors tetracycline resistance genes in mobile pdif modules located in distinct plasmids with similarities to those isolated from clinical strains. This finding suggests the potential for further acquisition of resistance determinants and serves as a cautionary signal against the unregulated introduction of antibiotics into vulnerable populations, highlighting the need for a targeted public health policy that guides the use of antibiotics in these communities. Healthcare practitioners working with remote communities can use these insights to make more informed treatment decisions, adopting rapid diagnostic tests, if possible, to ensure that antibiotics are prescribed only when clinically necessary. This minimizes unnecessary antibiotic exposure, thereby potentially reducing the emergence and spread of AMR (Yau et al. 2021). Therefore, expanding genomic surveillance to include community‐derived A. baumannii strains, even from remote indigenous tribes, is indeed a useful endeavor. Although this is a road less taken, the knowledge obtained, particularly tracking the shifts in known and novel STs, will be invaluable for understanding the pathogen's broader epidemiological dynamics and informing future public health strategies.
Author Contributions
Soo Sum Lean: conceptualization (equal), data curation (lead), formal analysis (lead), investigation (equal), methodology (lead), software (lead), visualization (lead), writing – original draft (lead), writing – review and editing (equal). Denise E. Morris: investigation (equal), methodology (equal), project administration (equal), resources (equal). Rebecca Anderson: investigation (equal), methodology (equal), project administration (equal), resources (equal). Ahmed Ghazi Alattraqchi: investigation (equal), methodology (equal), resources (equal), software (equal). David W. Cleary: conceptualization (equal), formal analysis (equal), funding acquisition (equal), resources (equal), software (equal), supervision (equal), writing – review and editing (equal). Stuart C. Clarke: conceptualization (equal), funding acquisition (lead), supervision (lead), resources (lead), writing – review and editing (equal). Chew Chieng Yeo: conceptualization (equal), formal analysis (equal), resources (equal), supervision (equal), validation (equal), writing – original draft (equal), writing – review and editing (equal).
Ethics Statement
Ethical approval for isolates taken in Peninsular Malaysia was provided by Universiti Sultan Zainal Abidin (UniSZA) Ethics Committee: Approval No. UniSZA/C/1/UHREC/628–1(85) dated June 27, 2016, the Department of Orang Asli Affairs and Development (JAKOA): Approval No. JAKOA/PP.30.052Jld11[42], and by the University of Southampton Faculty of Medicine Ethics Committee (Submission ID 20831). Written informed consent was taken with parents/guardians providing consent for those < 18 years old.
Consent
All authors have provided consent for publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Appendix.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agersø, Y. , and L. Guardabassi . 2005. “Identification of Tet 39, a Novel Class of Tetracycline Resistance Determinant in Acinetobacter spp. of Environmental and Clinical Origin.” Journal of Antimicrobial Chemotherapy 55: 566–569. 10.1093/jac/dki 051.15761075 · doi ↗ · pubmed ↗
- 2Ahmad, I. , A. Nadeem , F. Mushtaq , et al. 2023. “Csu Pili Dependent Biofilm Formation and Virulence of Acinetobacter baumannii .” NPJ Biofilms and Microbiomes 9: 101. 10.1038/s 41522-023-00465-6.38097635 PMC 10721868 · doi ↗ · pubmed ↗
- 3Akeda, Y. 2021. “Current Situation of Carbapenem‐Resistant Enterobacteriaceae and Acinetobacter in Japan and Southeast Asia.” Microbiology and Immunology 65: 229–237. 10.1111/1348-0421.12887.33913535 · doi ↗ · pubmed ↗
- 4Alcock, B. P. , W. Huynh , R. Chalil , et al. 2023. “CARD 2023: Expanded Curation, Support for Machine Learning, and Resistome Prediction at the Comprehensive Antibiotic Resistance Database.” Nucleic Acids Research 51: D 690–D 699. 10.1093/nar/gkac 920.36263822 PMC 9825576 · doi ↗ · pubmed ↗
- 5Ambrose, S. J. , and R. M. Hall . 2024. “Variation in the Plasmid Backbone and dif Module Content of R 3‐T 33 Acinetobacter Plasmids.” Plasmid 129–130: 102722. 10.1016/j.plasmid.2024.102722.38631562 · doi ↗ · pubmed ↗
- 6Aminov, R. I. 2009. “The Role of Antibiotics and Antibiotic Resistance in Nature.” Environmental Microbiology 11: 2970–2988. 10.1111/j.1462-2920.2009.01972.x.19601960 · doi ↗ · pubmed ↗
- 7Balalovski, P. , and I. Grainge . 2020. “Mobilization of pdif Modules in Acinetobacter: A Novel Mechanism for Antibiotic Resistance Gene Shuffling?” Molecular Microbiology 114: 699–709. 10.1111/mmi.14563.32594594 · doi ↗ · pubmed ↗
- 8Bertini, A. , L. Poirel , P. D. Mugnier , L. Villa , P. Nordmann , and A. Carattoli . 2010. “Characterization and PCR‐Based Replicon Typing of Resistance Plasmids in Acinetobacter baumannii .” Antimicrobial Agents and Chemotherapy 54: 4168–4177. 10.1128/AAC.00542-10.20660691 PMC 2944597 · doi ↗ · pubmed ↗