Taxonomic notes on the crab-spider genera Nyctimus Thorell, 1877 and Zametopina Simon, 1909 (Araneae, Thomisidae) with descriptions of six new species from Southeast Asia

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2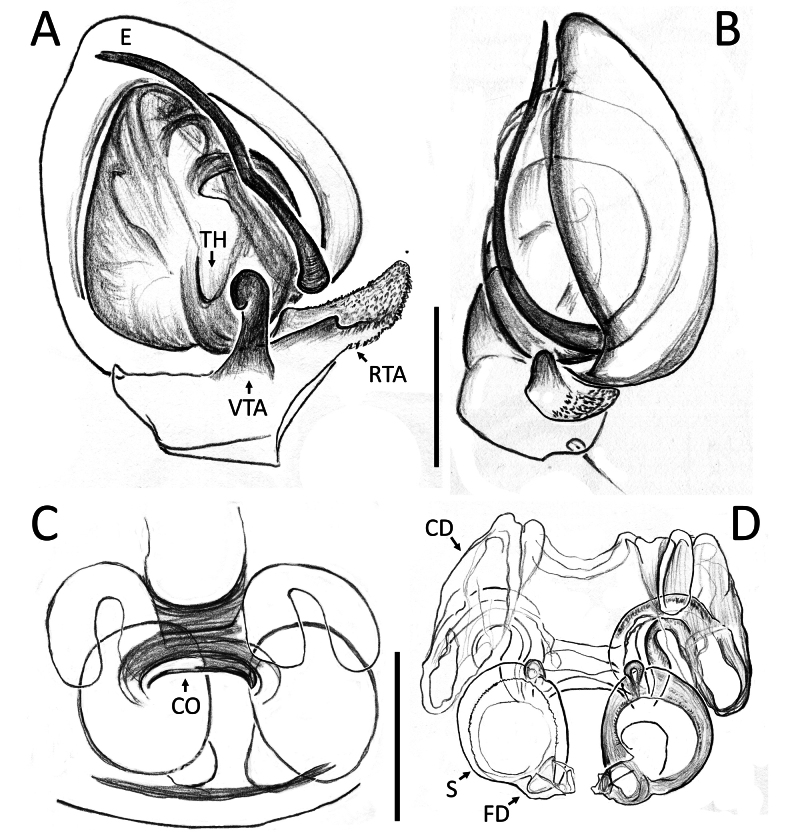 Figure 3
Figure 3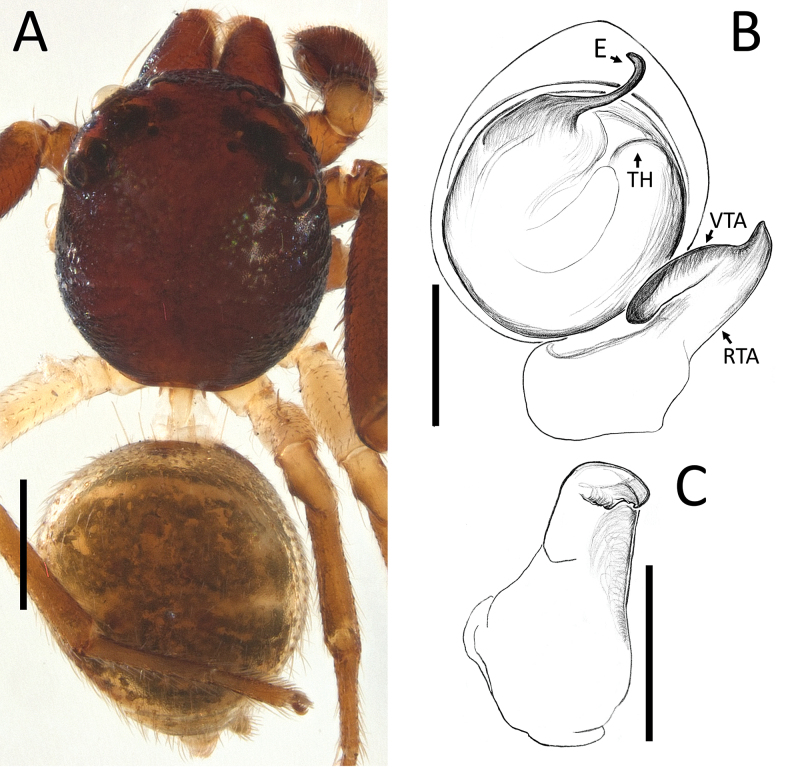 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6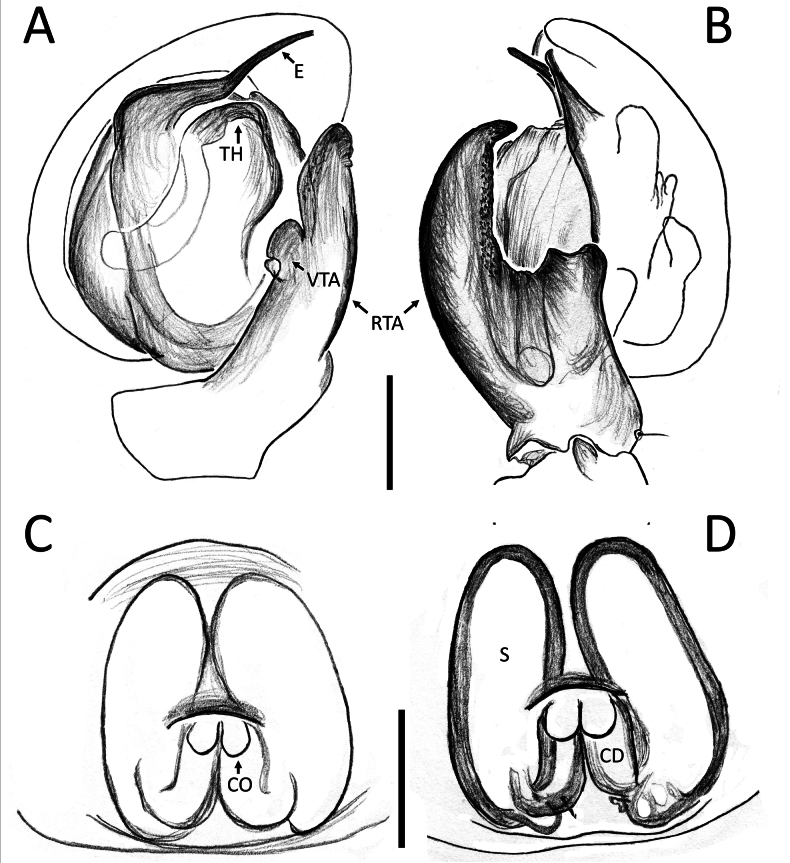 Figure 7
Figure 7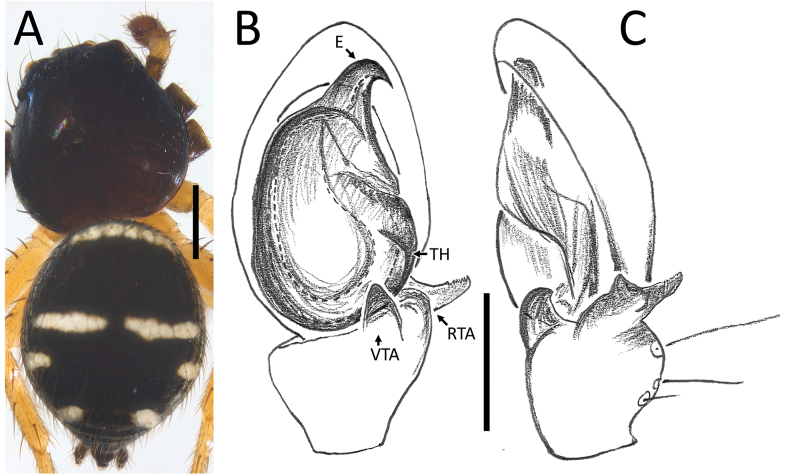 Figure 8
Figure 8 Figure 9
Figure 9 Figure 10
Figure 10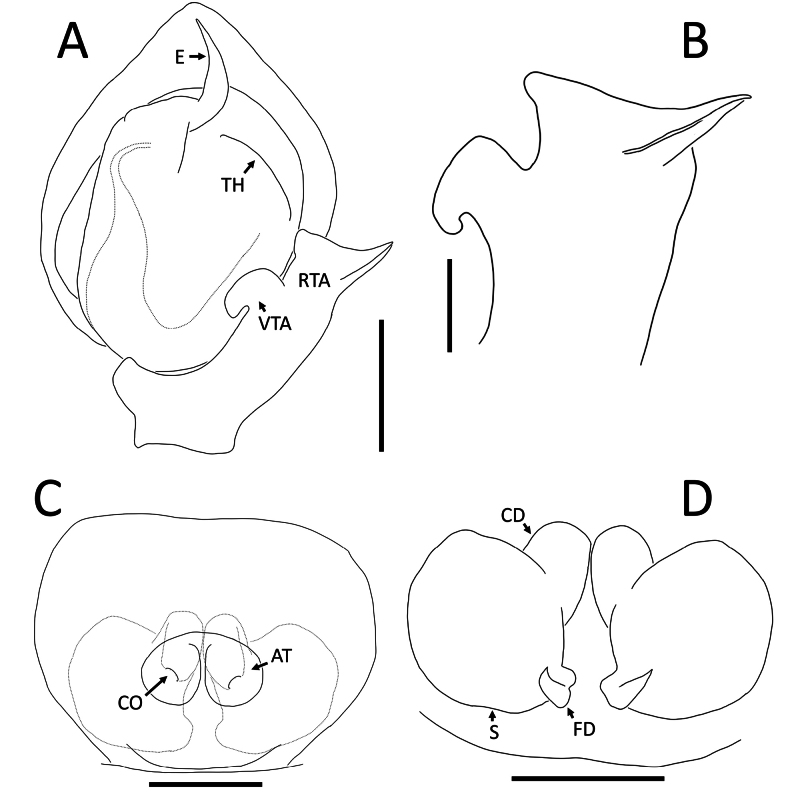 Figure 11
Figure 11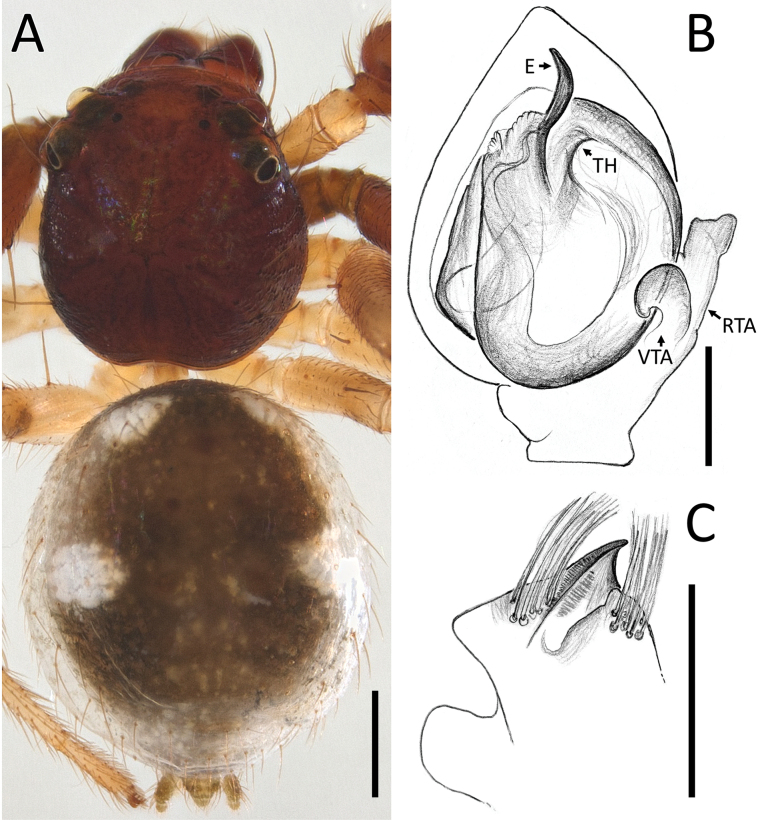 Figure 12
Figure 12 Figure 13
Figure 13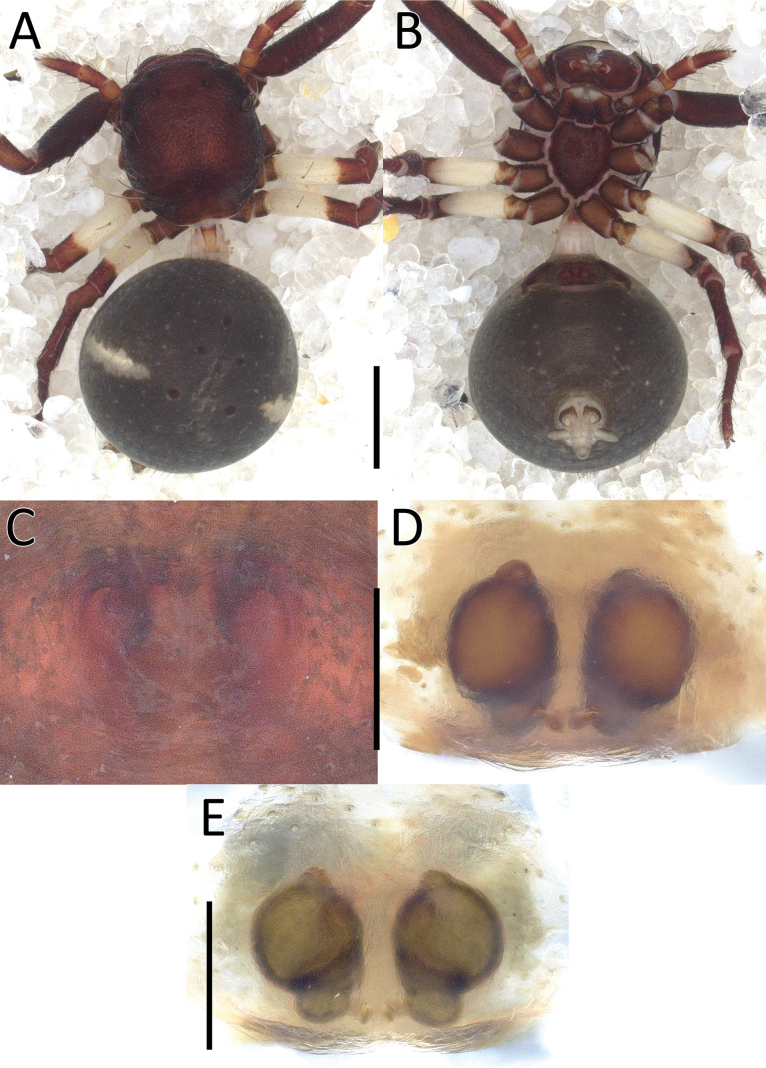 Figure 14
Figure 14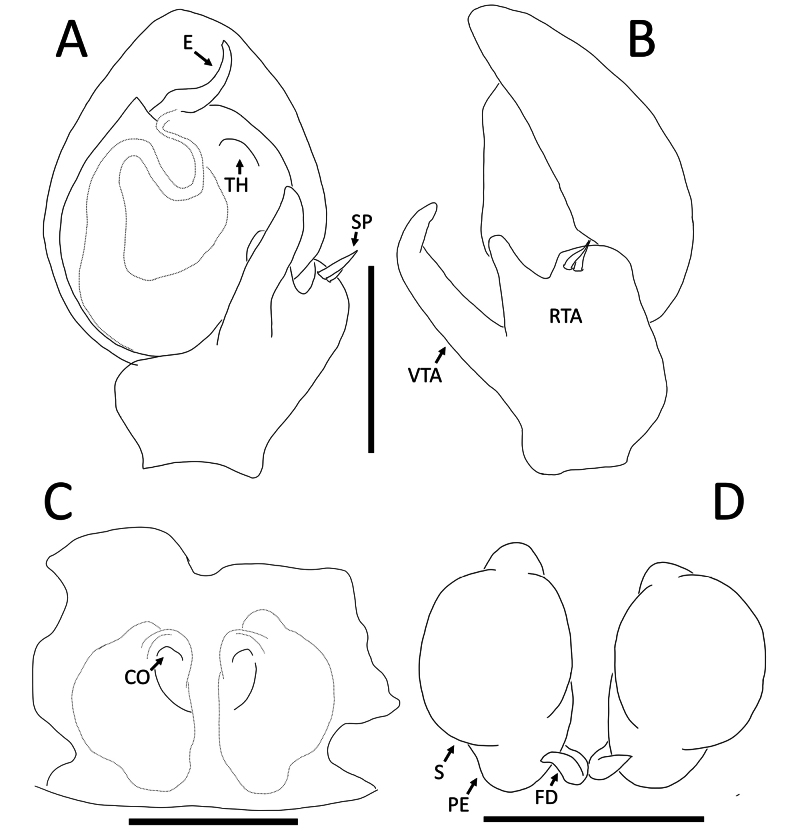 Figure 15
Figure 15 Figure 16
Figure 16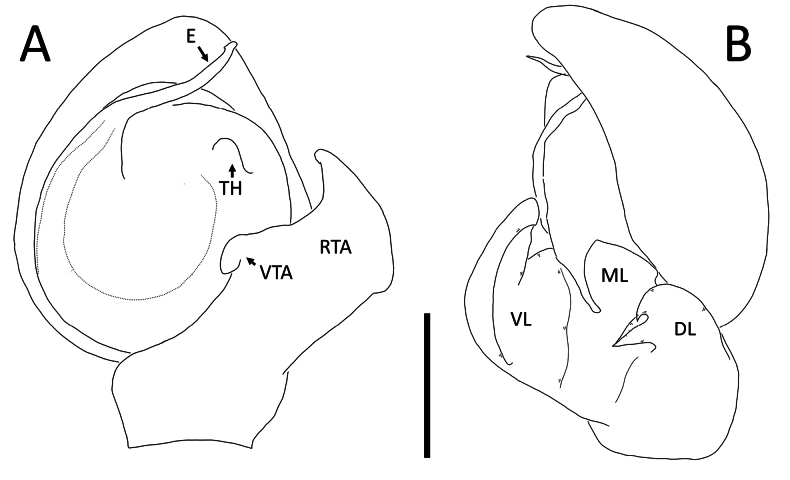 Figure 17
Figure 17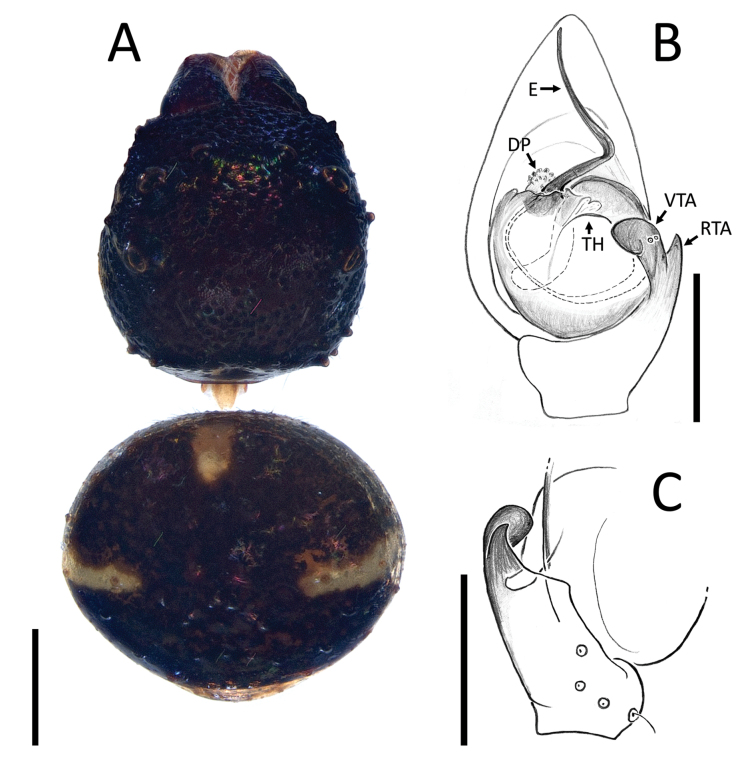 Figure 18
Figure 18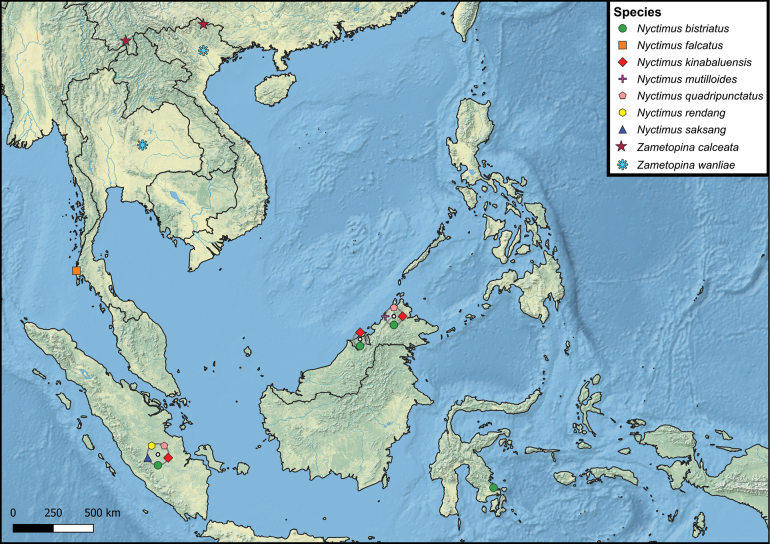 Figure 19
Figure 19- —Deutsche Forschungsgemeinschaft 501100001659 https://ror.org/018mejw64 http://doi.org/10.13039/501100001659
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSpider Taxonomy and Behavior Studies · Lepidoptera: Biology and Taxonomy · Orthoptera Research and Taxonomy
Introduction
Zametopina Simon, 1909, is a genus of crab spiders belonging to the family Thomisidae. Referred to as crab spiders, due to their elongated first two leg pairs and their habit of moving sideways, Thomisidae are among the most species-rich spider families with more than 2,100 species across 171 genera (World Spider Catalog 2025). Despite its inclusion in this diverse group, Zametopina remains relatively obscure and taxonomically unrevised. Initially, the genus comprised only a single species, Zametopina calceata Simon, 1909, which had been inadequately understood for decades due to a lack of illustrations in the original description. A significant update came with the work of Tang et al. (2010), who revisited and redescribed the type species. Through meticulous examination of the type specimen, they provided detailed illustrations and descriptions, thereby clarifying the diagnostic features of the genus and aiding in its taxonomic circumscription. This effort significantly enhanced the understanding of Zametopina and set the stage for future taxonomic studies. Recently, a second species, Zametopina wanliae Lin & Li, 2023, was described in Lin et al. (2023), further enriching the diversity of the genus.
By contrast, the genus Nyctimus Thorell, 1877, has received limited taxonomic scrutiny since its initial description. Historically, Nyctimus included two species: the type species Nyctimus bistriatus Thorell, 1877, from Indonesia (Sumatra and Sulawesi), and Nyctimus trimeni (Simon, 1895) from South Africa. Originally described as Zametopias trimeni, the latter is likely misplaced, indicating a need for taxonomic revision. Additionally, Zametopias speculator Thorell, 1892, was transferred to Nyctimus and synonymised with N. bistriatus by Lehtinen (2016). However, this transfer was made without accompanying illustrations or a comprehensive re-examination of the type material, leaving the taxonomy of both species inadequately resolved.
This paper aims to address some of these gaps by reviewing the poorly known species of both Zametopina and Nyctimus, primarily drawing on specimens collected from tropical Asia by the late Christa L. Deeleman-Reinhold, as well as from the EFForTS project in the lowlands of Jambi Province, Sumatra, Indonesia (Drescher et al. 2016). This review aims to clarify the taxonomy and provide updated descriptions, contributing to a more robust understanding of these genera and their roles in the biodiversity of the regions they inhabit. The study highlights the importance of revisiting historical collections and underscores the need for continued exploration and documentation of lesser-known spider taxa.
Materials and methods
Sample collection
Specimens from the late Christa L. Deeleman-Reinhold’s personal collection, housed at her residence, were selected by Suresh P. Benjamin (SPB) and transferred to the Zoological Research Museum Alexander König (Bonn, Germany, ZFMK) for this study. These specimens, along with the rest of her collection, will ultimately be deposited at Naturalis Biodiversity Center, Leiden, Netherlands (RMNH). Spiders from Jambi Province were part of a collection of more than half a million canopy arthropods (Pollierer et al. 2023) sampled by canopy fogging across rainforest, jungle rubber, and monocultures of rubber and oil palm in the lowlands of Jambi, Sumatra, Indonesia as part of the German-Indonesian EFForTS research project (Drescher et al. 2016). Overall, more than 10,000 spider individuals from at least 30 families and 400 species were captured (Ramos et al. 2022; Dhiya’ulhaq et al. 2024). The fogging method is detailed in Pollierer et al. (2023) and the EFForTS plot design and general study aims in Drescher et al. (2016).
Identification and photography
The general methodology follows Benjamin (2011) and Ileperuma Arachchi and Benjamin (2019). Specimens used for habitus illustrations were placed in 70% EtOH and photographed with a Zeiss AxioCam HRc camera mounted on a dissecting microscope (Zeiss Discovery V20) with top illumination and a magnification of up to 150×. Images were edited using the Zeiss ZEN Pro software package. Unless otherwise stated, depicted structures are of the left male palp. Alternatively, some specimens were examined under a ZEISS Stemi 2000 microscope. Female genitalia were excised from the specimen’s body and then cleared with 10% KOH for at least one hour to examine the internal copulatory organs. Imaging the specimens was done using a Keyence VHX-7000 digital microscope system or a Zeiss AxioCam HRc camera mounted on a dissecting microscope (Zeiss Discovery V20). The description of colouration is based on specimens in ethanol. Measurements of legs are given as total length (femur, patella, tibia, metatarsus, tarsus). Missing legs or leg segments are marked with “-”, and legs with missing segments do not have their total length recorded. Unmentioned legs did not have their length measured. All measurements are in millimeter.
Abbreviations
ALE anterior lateral eye
AME anterior median eye
CO copulatory opening
FD fertilization duct
PLE posterior lateral eye
PME posterior median eye
RTA retrolateral tibial apophysis of male palp
VTA ventral tibial apophysis of male palp
Repositories
GOETAnimal Ecology Group, J.-F. Blumenbach Institute of Zoology and Anthropology, University of Göttingen, Göttingen, Germany (Jochen Drescher, Stefan Scheu)
MCSNMuseo Civico di Storia Naturale Giacomo Doria, Genoa, Italy (Maria Tavano)
MCZMuseum of Comparative Zoology, Massachusetts (Gonzalo Giribet)
MNHNMuséum National d’Histoire Naturelle, Paris (Christine Rollard)
MZBMuseum Zoologicum Bogoriense, Bogor, Indonesia (Cahyo Rahmadi)
RMNHNaturalis Biodiversity Center, Leiden (Karen van Dorp)
SMFNaturmuseum Senckenberg, Frankfurt a. M., Germany (Jana Grüger, Peter Jäger)
ZMHZoologisches Museum Hamburg, Hamburg, Germany (Nadine Dupérré, Danilo Harms)
Results
Family Thomisidae Sundevall, 1833
Nyctimus
Taxon classificationAnimaliaAraneaeThomisidae
Thorell, 1877
10DC5F15-1D15-5FB7-AE1F-EDF552C40744
Type species.
Nyctimus bistriatus Thorell, 1877.
Diagnosis.
Small thomisids, 2–5 mm in body length. Prosoma cuboid, as long as wide, with granulated surface and bearing long macrosetae (Fig. 1). Body colouration dark with white/yellow markings, mostly in the form of spots and bars. Opisthosoma with five large sigillae arranged in a triangular formation Embolus is weakly curved and possesses a thick base. Tegulum with a “hood” (Fig. 3A). Tibial apophyses are well developed, VTA present, either free (Fig. 3A) or hook-shaped and attached to RTA (Fig. 7A). Epigynum highly variable, spermatheca rounded or oval, single pair (Fig. 3C, D).
Nyctimus bistriatus Thorell, 1877 from Borneo; habitus, dorsal view. A. Male (RMNH.ARA.17873); B. Female (RMNH.ARA.17875). Scale bars: 0.5 mm.
Species composition.
Nyctimus bistriatus Thorell, 1877 (♂♀), Nyctimus falcatus Benjamin & Dhiya’ulhaq, sp. nov. (♂), Nyctimus kinabaluensis Benjamin & Dhiya’ulhaq, sp. nov. (♂♀), Nyctimus mutilloides Benjamin & Dhiya’ulhaq, sp. nov. (♂), Nyctimus quadripunctatus Dhiya’ulhaq & Benjamin, sp. nov. (♂♀), Nyctimus rendang Dhiya’ulhaq & Benjamin, sp. nov. (♂♀), Nyctimus saksang Dhiya’ulhaq & Benjamin, sp. nov. (♂), Nyctimus trimeni (Simon, 1895).
Nyctimus
bistriatus
Taxon classificationAnimaliaAraneaeThomisidae
Thorell, 1877
632872F7-93A8-58A6-A18B-482B3EEB3773
Nyctimus bistriatus Thorell, 1877: 499; Lehtinen 2016: 166 (as Nyctimus binotatus, lapsus). Zametopias speculator Thorell, 1892: 123.
Type material.
**Holotype. ** Indonesia – Sulawesi, Southeast Sulawesi Province • ♂; Kendari; 1874; O. Beccarii leg.; holotype of Nyctimus bistriatus Thorell, 1877; MCSN. Examined.
Other material examined.
Brunei – Tutong District • 1♂; Bukit Sulang; 1982; N. Stork leg.; canopy fogging in primary forest; RMNHRMNH.ARA.17873. Indonesia – Sumatra, Jambi Province • 1♀; Sarolangun, Bukit Duabelas National Park; 01°58'55.2"S, 102°45'02.6"E; elev. 73 m; 7 Oct. 2013; J. Drescher leg.; canopy fogging in rainforest; GOET 2013_BF2.1_AraThom051N_001 (to be transferred to MZB). Malaysia – Sabah State • 1♂; Kinabalu National Park, Poring Hot Springs; 05°59'N, 116°42'E; elev. 600–700 m; 27 Feb. 1996; A. Floren leg.; canopy fogging on Aporosa lagenocarpa tree, in primary forest; RMNHRMNH.ARA.17874. • 1♀; Kinabalu National Park, Poring Hot Springs; 05°59'N, 116°42'E; elev. 600–700 m; 28 Mar. 1998; A. Floren leg.; canopy fogging on Barringtonia tree, in primary forest; RMNHRMNH.ARA.17875. • 1♂; Kinabalu National Park, Poring Hot Springs; 05°59'N, 116°42'E; elev. 600–700 m; 28 Mar. 1998; A. Floren leg.; canopy fogging on Dacryodes laxa tree, in primary forest; RMNHRMNH.ARA.17876.
Diagnosis.
Males can be easily distinguished from all other congeners by the longer embolus, more than length of cymbium (in all congeners, the embolus is at most half the length of cymbium; Fig. 3A, B) and the denticulate RTA. Females can be easily distinguished by the single CO; CD arched, resembling a rams’ horn, more than twice the length of spermathecae (in all congeners, the CO is paired, and the CD are not longer than spermathecae; Figs 2C, D, 3C, D).
Nyctimus bistriatus Thorell, 1877 female from Sumatra (2013 BF2.1 AraThom051N 001). A. Habitus, dorsal view; B. Prosoma, frontal view; C. Epigynum, ventral view; D. Epigynum, dorsal view, cleared. Scale bars: 1 mm (A); 0.5 mm (B); 0.2 mm (C, D).
Nyctimus bistriatus Thorell, 1877 from Borneo. A. Male (RMNH.ARA.17873), left palp, ventral view; B. ibid., retrolateral view; C. Female (RMNH.ARA.17875), epigynum; C. ibid., ventral view; D. ibid., dorsal view. Abbreviations: CD = copulatory duct; CO = copulatory opening; E = embolus; FD = fertilization duct; RTA = retrolateral tibial apophysis; S = spermatheca; TH = tegular hood; VTA = ventral tibial apophysis. Scale bars: 0.2 mm.
Description.
Male (RMNH.ARA.17873; Figs 1A, 3A, B). Total length 2.85. Prosoma length 1.42; width 1.26. Opisthosoma length 1.43; width 1.17. Diameter of eyes: AME 0.09; ALE 0.15; PLE 0.14 PME 0.07. Interdistances between eyes: AME–AME 0.50; AME–ALE 0.10; ALE–ALE 0.86; PME–PME 0.70; PME–PLE 0.20; ALE–PLE 0.18; AME–PME 0.13; PLE–PLE 1.09. Leg measurements: leg I 3.3 (1.0, 0.5, 1.0, 0.8, 0.5). Prosoma cuboid, almost as long as wide; dorsally blackish brown, surface granulated; laterally with two rows of long, curved macrosetae originating on small tubercles. All eyes are very far apart, especially between the median eyes. Chelicerae, labium and sternum blackish brown; sternum with small dents. Legs dark brown except femora III and IV, which are yellow; femora and tibiae I–IV with two long spines; patellae I–IV with one distal long spine. Opisthosoma dorsally greyish black with many long setae; five large sigillae present, arranged in a triangular formation; venter greyish black.
Palp (Fig. 3A, B): cymbium oval. Embolus long, whip-like, looping over the retrolateral side of bulb, half of its length covered by the cymbium. Tegular hood small, located close to the base of bulb. RTA large, roughly trapezoid, laterally protruding, distal half with denticles on all sides. VTA long, digitiform; apically flexed with tip ventrally curved.
Female (RMNH.ARA.17875; Figs 1B, 3C, D). Total length 3.32. Prosoma length 1.15; width 1.30. Opisthosoma length 2.17; width 1.95. Diameter of eyes: AME 0.08; ALE 0.13; PLE 0.16; PME 0.06. Interdistances between eyes: AME–AME 0.56; AME–ALE 0.10; ALE–ALE 0.86; PME–PME 0.64; PME–PLE 0.21; ALE–PLE 0.18; AME–PME 0.16; PLE–PLE 1.09. Leg measurements: leg I 5.3 (1.6, 0.6, 1.3, 1.0, 0.8). Habitus as in male, except opisthosoma dorsally lighter than male and the presence of white spots as shown in Fig. 1B.
Epigynum (Figs 2C, D, 3C, D): exterior plate roughly pentagonal. Atrium inconspicuous. CO oval, single, large, CD long; divided into a very wide, ramshorn-shaped initial part and a narrow, C-shaped later part. Spermatheca globular, approximately as wide as CO; anteriorly with a small bump; posteriorly extended.
Distribution.
Brunei, Malaysia (Borneo: Sabah), Indonesia (Sumatra: Jambi; Sulawesi: Southeast Sulawesi) (Fig. 19).
Nyctimus
falcatus
Taxon classificationAnimaliaAraneaeThomisidae
Benjamin & Dhiya’ulhaq sp. nov.
10A23D92-4157-560C-B4FD-69AF2B3C67A8
https://zoobank.org/8DF08156-9B26-4343-95A5-641924B7AC50
Etymology.
The species epithet falcatus is derived from the Latin word falx/falcis, meaning “sickle” or “scythe.” It refers to the distinctly curved, sickle-shaped embolus of the male palp, a diagnostic feature of the species.
Type material.
**Holotype. ** Thailand – Phang Nga Province • ♂; Khao Lak National Park; no more label data; RMNHRMNH.ARA.17861.
Diagnosis.
Males are very similar to N. saksang sp. nov., in having a similar sickle-shaped embolus, but they can be easily distinguished from it by the large, digitiform VTA, attached distally on the RTA (the corresponding part in N. saksang, sp. nov., is much smaller and attached medially on the ventral lobe of RTA; Fig. 4B vs Figs 16C, 17A).
Description.
Male (holotype RMNH.ARA.17861; Fig. 4). Total length: 2.36. Prosoma length 1.23; width: 1.05. Opisthosoma length 1.13; width 1.02. Diameter of eyes: AME 0.08; ALE 0.15; PLE 0.14; PME 0.05. Interdistances between eyes: AME–AME 0.27; AME–ALE 0.08; ALE–ALE 0.59; PME–PME 026; PME–PLE 0.27; ALE–PLE 0.18; AME–PME 0.14; PLE–PLE 0.79. Prosoma cuboid, almost as long as wide, dark reddish brown; surface weakly granulated; laterally adorned with long macrosetae, each arising from a small tubercle. Chelicerae and sternum coloured as prosoma. Front legs brown; back legs brown except for the white femora III and IV. Opisthosoma round, light brown; dorsally covered with scutum; five large sigillae present, arranged in a triangular formation.
Nyctimus falcatus Benjamin & Dhiya’ulhaq, sp. nov., male holotype (RMNH.ARA.17861). A. Habitus, dorsal view; B. Left palp, ventral view; C. Left palpal tibia, retrolateral view. Abbreviations: E = embolus; RTA = retrolateral tibial apophysis; TH = tegular hood; VTA = ventral tibial apophysis. Scale bars: 0.5 mm (A); 0.2 mm (B, C).
Palp (Fig. 4B, C): cymbium oval. Embolus sickle-shaped. Tegulum round, with a small triangular projection next to the embolus; tegular hood small. RTA wide trapezoid; VTA large, digitiform, attached distally on the RTA.
Female. Unknown.
Distribution.
Only known from the type locality (Thailand: Phang Nga) (Fig. 19).
Remarks.
The specimen is partially damaged: both first legs are broken and incomplete, and the remaining legs are mostly missing or fragmented.
Nyctimus
kinabaluensis
Taxon classificationAnimaliaAraneaeThomisidae
Benjamin & Dhiya’ulhaq sp. nov.
A719868B-03C7-5999-BD87-2DD47B99BD6E
https://zoobank.org/1F2D858B-2FED-4397-BE73-9B841300D782
Etymology.
The specific epithet is derived from the name of the type locality.
Type material.
**Holotype. ** Malaysia – Sabah State • ♂; Kinabalu National Park, Poring Hot Springs; 05°59'N, 116°42'E; elev. 600–700 m; 27 Feb. 1996; A. Floren leg.; canopy fogging on Barringtonia tree, in primary forest; RMNHRMNH.ARA.17862.
Other material examined.
Brunei – Tutong District • 1♀; Lamunin, Bukit Sulang; 1982; N. Stork leg.; canopy fogging; RMNHRMNH.ARA.17859. Indonesia – Sumatra, Jambi Province • 1♀; Sarolangun, Bukit Duabelas National Park; 01°58'55.2"S, 102°45'02.6"E; elev. 73 m; 7 Oct. 2013; J. Drescher leg.; canopy fogging in rainforest; GOET 2013_BF2.1_AraThom004N_001 (to be transferred to MZB).
Diagnosis.
Males can be easily distinguished from all other congeners by the embolic base abruptly bent 90° towards the retrolateral side of the cymbium and the tapered, bent embolus (Fig. 7A, B). Females are distinguishable by the long, oval spermatheca (globular or short-oval in all other congeners; Figs 6C, D, 7C, D).
Description.
Male (holotype RMNH.ARA.17862; Figs 5A, B, 7A, B). Total length 3.24. Prosoma length 1.61; width 1.50. Opisthosoma length 1.63; width 1.31. Diameter of eyes: AME 0.11; ALE 0.20; PLE 0.17; PME 0.08. Interdistances between eyes: AME–AME 0.44; AME–ALE 0.11; ALE–ALE 0.81; PME–PME 0.45; PME–PLE 0.36; ALE–PLE 0.21; AME–PME 0.17; PLE–PLE 1.15. Leg measurements: leg I 5.0 (1.5, 0.6, 1.3, 1.0, 0.6). Prosoma cuboid, almost as long as wide, dark reddish brown; surface weakly granulated; laterally adorned with long macrosetae, each arising from a small tubercle. Chelicerae and sternum coloured as prosoma. Endites brown with white tip. Front legs brown; back legs brown except for the white femora. Opisthosoma oval, greyish brown with white spots (most dense in the anterior half) and paired white bars in the middle; anteriorly with a white border; five large sigillae present, arranged in a triangular formation.
Nyctimus kinabaluensis Benjamin & Dhiya’ulhaq sp. nov., from Borneo. A. Male holotype (RMNH.ARA.17862), prosoma, dorsal view; B. ibid., opisthosoma, dorsal view; C. Female (RMNH.ARA.17859), habitus, dorsal view. Scale bars: 0.5 mm.
Nyctimus kinabaluensis Benjamin & Dhiya’ulhaq, sp. nov., from Sumatra. A. Female (2013_BF2.1_AraThom004N_001), habitus, dorsal view; B. Prosoma, frontal view; C. Epigynum, ventral view; D. Epigynum, dorsal view, cleared. Scale bars: 1 mm (A); 0.5 mm (B); 0.2 mm (C, D).
Nyctimus kinabaluensis Benjamin & Dhiya’ulhaq, sp. nov., from Borneo. A. Male (RMNH.ARA.17859), left palp, ventral view; B. ibid., retrolateral view; C. Female (RMNH.ARA.17862), epigynum, ventral view; D. ibid., dorsal view. Abbreviations: CD = copulatory duct; CO = copulatory opening; E = embolus; RTA = retrolateral tibial apophysis; S = spermatheca; TH = tegular hood; VTA = ventral tibial apophysis. Scale bars: 0.2 mm.
Palp (Fig. 7A, B): cymbium oval. Embolus elongated, tapering, with a 150° bend in the middle. Tegular hood small. RTA bilobed; ventral lobed very large, claw-shaped; dorsal lobe much smaller than ventral lobed, roughly triangular with an obtuse tip and a small triangular projection; VTA hook-shaped, small, attached medially on the RTA.
Female (RMNH.ARA.17859; Figs 5C, 7C, D). Total length: 3.38. Prosoma length 1.48; width 1.27. Opisthosoma length 1.90; width 1.70. Diameter of eyes: AME 0.07; ALE 0.17; PLE 0.14; PME 0.06. Interdistances between eyes: AME–AME 0.38; AME–ALE 0.12; ALE–ALE 0.74; PME–PME 0.39; PME–PLE 0.33; ALE–PLE 0.22; AME–PME 0.17; PLE–PLE 0.98. Leg measurements: leg I 4.6 (1.3, 0.4, 1.1, 1.0, 0.8). Habitus as in male.
Epigynum (Fig. 7C, D): exterior plate large, roughly square with rounded sides. Atrium oval. CO diagonally oriented, anteriorly facing. CD arched. Spermatheca long oval, with a slight bump anteriorly.
Distribution.
Brunei; Malaysia (Borneo: Sabah); Indonesia (Sumatra: Jambi) (Fig. 19).
Nyctimus
mutilloides
Taxon classificationAnimaliaAraneaeThomisidae
Benjamin & Dhiya’ulhaq sp. nov.
5A830190-ADB7-526B-8CE7-B63CF6B773FB
https://zoobank.org/04BD7F24-E0FF-49A5-A849-3C9430494FA4
Etymology.
The specific epithet is derived from the latin name of the velvet ant family Mutillidae, and the Greek-derived suffix -oides, meaning “resembling” or “like.” Thus, mutilloides translates to “mutillid-like,” referring to the spider’s striking abdominal patterns resembling abdominal markings of certain trogaspidiine mutillids.
Type material.
**Holotype. ** Malaysia – Sabah State • ♂; Kinabalu National Park, Poring Hot Springs; 05°59'N, 116°42’E; elev. 600–700 m; 9 Feb. 1997; A. Floren leg.; canopy fogging; RMNHRMNH.ARA.17882.
Diagnosis.
Nyctimus mutilloides, sp. nov., can be readily distinguished from its congeners by its vivid white markings on the opisthosoma, forming a pattern highly suggestive of mutillid wasps (spots on all congeners). Furthermore, the male palp has a claw-shaped embolus that juts out from the distal margin of the bulb (different in other congeners; Fig. 8B, C).
Nyctimus mutilloides Benjamin & Dhiya’ulhaq, sp. nov., male holotype (RMNH.ARA.17882). A. Habitus, dorsal view; B. Left palp ventral view; C. Left palp retrolateral view. Abbreviations: E = embolus; RTA = retrolateral tibial apophysis; TH = tegular hood; VTA = ventral tibial apophysis. Scale bars: 0.5 mm (A); 0.2 mm (B, C).
Description.
Male (holotype RMNH.ARA.17882; Fig. 8). Total length 2.80. Prosoma length 1.22; width 1.23. Opisthosoma length 1.57; width 1.22. Diameter of eyes: AME 0.07; ALE 0.12; PLE 0.10; PME 0.08. Interdistances between eyes: AME–AME 0.16; AME–ALE 0.11; ALE–ALE 0.39; PME–PME 0.2; PME–PLE 0.18; ALE–PLE 0.10; AME–PME 0.07; PLE–PLE 0.69. Prosoma cuboid, almost as long as wide, slightly flattened, weakly granulated, dark brown with darker margins, laterally adorned with long macrosetae; cephalic region elevated. Legs slender, pale yellow with indistinct annulations. No conspicuous spines observed. Opisthosoma oval, dorsoventrally flattened, with white spots and bands on black background.
Palp (Fig. 8B, C): cymbium slender, oval; tip rounded. Embolus claw-shaped, curved with a pointed tip, emerging distally from the tegulum. Tegulum oval; tegular hood wide, located retrolaterally near the base. RTA bilobed; both lobes roughly triangular; dorsal lobe with a serrated edge. VTA short, thumb-shaped, and slightly curved.
Female. Unknown.
Distribution.
Known only from the type locality (Malaysia: Borneo: Sabah) (Fig. 19).
Remarks.
The white dots and stripes on the opisthosoma of N. mutilloides are reminiscent of those of mutillid wasps, especially those of the tribe Trogaspidiini (see figs 26–37 in Okayasu 2023). Whether this constitutes a form of Batesian mimicry is unclear, but it is possible given that certain Gnaphosidae, Salticidae, and Clubionidae seem to mimic velvet ants (Nentwig 2008). Its presence in canopy fogging samples indicates an arboreal lifestyle, a noteworthy ecological trait within the genus. Further surveys and molecular data could shed light on its placement within Nyctimus and potential mimicry evolution in this group.
Nyctimus
quadripunctatus
Taxon classificationAnimaliaAraneaeThomisidae
Dhiya’ulhaq & Benjamin sp. nov.
14EA4F9E-30F1-5BC0-907E-E5D2595607ED
https://zoobank.org/EA408798-5CD6-4401-B44E-F6127489C4C1
Etymology.
The specific epithet is taken from Latin, referring to the four white spots on the opisthosoma.
Type material.
**Holotype. ** Indonesia – Sumatra, Jambi Province • ♂; Batang Hari, Hutan Harapan Conservation Area; 02°11'15.3"S, 103°20'36.0"E; elev. 69 m; 7 Aug. 2013; J. Drescher leg.; canopy fogging in rainforest; GOET 2013_HF4.2_AraThom001N_001 (to be transferred to MZB). Paratypes. Indonesia – Sumatra, Jambi Province • 1♂1♀; Batang Hari, Hutan Harapan Conservation Area; 02°10'42.4"S, 103°19'58.2"E; elev. 54 m; 21 Jul. 2013; J. Drescher leg.; canopy fogging in rainforest; GOET 2013_HF3.2_AraThom001N_001–2 (to be transferred to MZB) • Malaysia – Sabah State • 1♂; Kinabalu National Park, Poring Hot Springs; 05°59'N, 116°42'E; elev. 600–700 m; 28 Mar. 1998; A. Floren leg.; canopy fogging on Barringtonia tree, in primary forest; RMNHRMNH.ARA.17858. • 1♂; Kota Marudu, Kampong Sorinsim; 06°06'N, 116°50'E; elev. 600–700 m; 2 Mar. 1997; A. Floren leg.; canopy fogging in 15 years old secondary forest; RMNHRMNH.ARA.17860.
Other material examined.
Malaysia – Sabah State • 1♂; Kinabalu National Park, Poring Hot Springs; 05°59'N, 116°42'E; elev. 600–700 m; 27 Feb. 1996; A. Floren leg.; canopy fogging on Aporosa subcaudata tree, in primary forest; RMNHRMNH.ARA.17857.
Diagnosis.
Males are easily distinguished from other congeners by the anvil-shaped RTA ending in two acute lobes (Fig. 11A, B). Females can be distinguished from N. bistriatus by the much thinner, shorter CD and paired CO (Figs 10C, D, 11C, D; vs single CO in N. bistriatus, Figs 2C, 3C), and from N. rendang, sp. nov., by the spermatheca lacking a prominent posterior extension (Figs 10D, 11D; vs Figs 14D, 15D).
Description.
Male (holotype 2013_HF4.2_AraThom001N_001; Figs 9, 11A, B). Total length 2.95. Prosoma length 1.35; width 1.23. Opisthosoma length 1.60; width 1.22. Diameter of eyes: AME 0.07; ALE 0.16; PLE 0.16; PME 0.06. Interdistances between eyes: AME–AME 0.38; AME–ALE 0.08; ALE–ALE 0.68; PME–PME 0.34; PME–PLE 0.32; ALE–PLE 0.19; AME–PME 0.18; PLE–PLE 0.98. Clypeus height 0.00. Leg measurements: leg I 4.25 (1.19, 0.49, 1.06, 0.90, 0.61); leg II 3.89 (1.11, 0.42, 0.90, 0.81, 0.65); leg III 2.63 (0.86, 0.35, 0.57, 0.49, 0.36); leg IV 2.74 (0.83, 0.27, 0.66, 0.52, 0.46). Prosoma cuboid, almost as long as wide, box-shaped, weakly granulated, highest at mid-point, light brown; lateral margins furnished with a row of long macrosetae. Lateral eyes large, situated on tubercles on the lateral edge of the prosoma. Chelicerae, endites, and labium light brown. Sternum light brown. Legs I light brown; others yellow-brown, with sparse spines and black setae. Opisthosoma oval, brown, with two pairs of large white spots. Spinnerets brown.
Nyctimus quadripunctatus Dhiya’ulhaq & Benjamin, sp. nov., male holotype (AraThom001N_2013_HF4.2_001). A. Habitus, dorsal view; B. Habitus ventral view; C. Right palp, mirrored, ventral view; D. Right palp, mirrored, retrolateral view. Scale bars: 1 mm (A, B); 0.2 mm (C, D).
Nyctimus quadripunctatus Dhiya’ulhaq & Benjamin, sp. nov., female paratype (AraThom001N_2013_HF3.2_001). A. Habitus, ventral view; B. Habitus, dorsal view; C. Epigynum, ventral view; D. Epigynum, dorsal view, cleared. Scale bars: 1 mm (A, B); 0.2 mm (C, D).
Nyctimus quadripunctatus Dhiya’ulhaq & Benjamin, sp. nov. A. Male holotype (AraThom001N_2013_HF3.2_001), right palp, mirrored, ventral view; B. ibid., RTA, ventro-retrolateral view; C. Female paratype (AraThom001N_2013_HF3.2_001), epigynum, ventral view; D. ibid., dorsal view. Abbreviations: CD = copulatory duct; CO = copulatory opening; E = embolus; RTA = retrolateral tibial apophysis; S = spermatheca; TH = tegular hood; VH = ventral hook. Scale bars: 0.2 mm.
Palp (Figs 9C, D, 11A, B): cymbium oval. Embolus sickle-shaped; base slightly wrinkled. Tegular hood very wide. RTA large, rectangular, anvil-shaped; dorsally bifurcated, ending into two long, acute lobes, hidden behind a brush of setae; VTA hook-shaped attached medially on RTA.
Female (paratype 2013_HF3.2_AraThom001N_002; Figs 10, 11C, D). Total length 4.81. Prosoma length 1.95; width 1.77. Opisthosoma length 2.86; width 2.65. Diameter of eyes: AME 0.09; ALE 0.20; PLE 0.18; PME 0.07. Interdistances between eyes: AME–AME 0.53; AME–ALE 0.14; ALE–ALE 0.97; PME–PME 0.52; PME–PLE 0.39; ALE–PLE 0.23; AME–PME 0.24; PLE–PLE 1.37. Leg measurements: leg I 5.47 (1.66, 0.57, 1.34, 1.13, 0.77); leg II -; leg III 3.69 (1.22, 0.47, 0.87, 0.63, 0.50); leg IV 4.12 (1.36, 0.46, 0.96, 0.76, 0.58). Habitus as in males, except prosoma and leg I reddish brown.
Epigynum (Figs 10C, D, 11C, D): exterior plate large, hexagonal-shaped with wide anterior side. Atrium conspicuous, inverted heart-shaped with a separation in the middle. CO diagonally oriented, posteriorly facing. CD arch-shaped. Spermatheca elongated, peanut-shaped, with a small bump anteriorly.
Distribution.
Malaysia (Borneo: Sabah); Indonesia (Sumatra: Jambi) (Fig. 19).
Remarks.
Specimens from Borneo show slight differences in shape of embolus and dorsal tip of RTA (Fig. 12B, C).
Nyctimus quadripunctatus Dhiya’ulhaq & Benjamin, sp. nov., non-type male specimen from Borneo (RMNH.ARA.17857). A. Habitus, dorsal view; B. Left palp, ventral view; C. Left palpal tibia, retrolateral view. Abbreviations: E = embolus; RTA = retrolateral tibial apophysis; TH = tegular hood; VH = ventral hook. Scale bars: 0.5 mm (A); 0.2 mm (B, C).
Nyctimus
rendang
Taxon classificationAnimaliaAraneaeThomisidae
Dhiya’ulhaq & Benjamin sp. nov.
043239DA-7E40-5B97-9B41-29D6254CD462
https://zoobank.org/D674C659-6FE3-41A2-9484-300E4C2A918D
Etymology.
The specific epithet refers to a meat dish associated with the cuisine of the Minangkabau people from West Sumatra. The meat is slow-cooked in a plethora of spices, giving it a dark colouration. The dark colour of N. rendang, sp. nov., is reminiscent of the colour of this dish.
Type material.
**Holotype. ** Indonesia – Sumatra, Jambi Province • ♂; Sarolangun, Air Hitam, Desa Baru; 02°01'49.5"S, 102°46'14.8"E; elev. 57 m; 12 Jul. 2013; J. Drescher leg.; canopy fogging in jungle rubber plantation; GOET 2013_BJ6.1_AraThom010N_001 (to be transferred to MZB). Paratypes. Indonesia – Sumatra, Jambi Province • 1♂1♀; same data as holotype; GOET 2013_BJ6.1_AraThom010N_002, 003 (to be transferred to MZB). • 2♂♂; Batang Hari, Bajubang, Singkawang; 01°47'07.9"S, 103°16'37.4"E; elev. 56 m; 18 Jun. 2013; J. Drescher leg.; canopy fogging in jungle rubber plantation; GOET 2013_HJ4.2_AraThom010N_001, 002 (to be transferred to ZMH).
Other material examined.
Indonesia – Sumatra, Jambi Province • 1♀; Batang Hari, Hutan Harapan Conservation Area; 02°11'15.3"S, 103°20'36.0"E; elev. 69 m; 7 Aug. 2013; J. Drescher leg.; canopy fogging in rainforest; GOET 2013_HF4.1_AraThom036N_001.
Diagnosis.
Males are similar to N. quadripunctatus, sp. nov., in having a sickle-shaped embolus and in the general shape of the bulb, but the VTA is digitiform and free (Fig. 15A, B; vs hook-shaped, attached to RTA in N. quadripunctatus, Fig. 11A, B) and the RTA possesses a pillar-shaped ventral lobe and paired, thick spines (vs both absent). Females are distinct in having the spermathecae with a posterior extension and a short, barely noticeable CD (Fig. 11D).
Description.
Male (holotype 2013_BJ6.1_AraThom010N_001; Figs 13, 15A, B). Total length 2.41. Prosoma length 1.16; width 1.03. Opisthosoma length 1.25; width 1.21. Diameter of eyes: AME 0.07; ALE 0.15; PLE 0.14; PME 0.05. Interdistances between eyes: AME–AME 0.35; AME–ALE 0.08; ALE–ALE 0.62; PME–PME 0.32; PME–PLE 0.24; ALE–PLE 0.17; AME–PME 0.19; PLE–PLE 0.77. Leg measurements: leg I 3.36 (0.90, 0.29, 0.92, 0.75, 0.50); leg II 3.46 (1.04, 0.32, 0.84, 0.75, 0.51); leg III 2.11 (0.67, 0.20, 0.54, 0.38, 0.32); leg IV 2.25 (0.75, 0.18, 0.59, 0.42, 0.31). Prosoma cuboid, almost as long as wide, dark reddish brown; surface weakly granulated; laterally adorned with long macrosetae, each arising from a small tubercle. Chelicerae, and sternum coloured as prosoma. Endites brown with white tip. Front legs brown; back legs brown except for the white femora. Opisthosoma round, dark brown with paired white bars in the middle; dorsally covered with scutum; five large sigillae present, arranged in a triangular formation.
Nyctimus rendang Dhiya’ulhaq & Benjamin, sp. nov., male holotype (AraThom010N_2013_BJ6.1_001). A. Habitus, dorsal view; B. Habitus, ventral view; C. Left palp, ventral view; D. Left palp, retrolateral view. Scale bars: 1 mm (A, B); 0.2 mm (C, D).
Palp (Figs 13C, D, 15A, B): cymbium oval. Embolus sickle-shaped. Tegulum with a small triangular projection next to the embolus; tegular hood small. RTA bilobed; ventral lobe finger-shaped; dorsal lobe wide trapezoid, apically with a pair of short, thick spines. VTA long, finger-shaped, apically flexed.
Female (paratype AraThom010N_2013_BJ6.1_003; Figs 14, 15C, D). Total length 3.86. Prosoma length 1.67; width 1.43. Opisthosoma length 2.19; width 2.21. Diameter of eyes: AME 0.08; ALE 0.18; PLE 0.19; PME 0.07. Interdistances between eyes: AME–AME 0.55; AME–ALE 0.12; ALE–ALE 0.90; PME–PME 0.45; PME–PLE 0.34; ALE–PLE 0.23; AME–PME 0.23; PLE–PLE 1.11. Leg measurements: leg I 4.85 (0.71, 1.01, 1.18, 0.49, 1.46); leg II -; leg III 3.05 (1.00, 0.30, 0.75, 0.54, 0.46); leg IV 3.41 (1.14, 0.32, 0.83, 0.65, 0.47). Habitus as in male, except opisthosoma not covered in scutum.
Nyctimus rendang Dhiya’ulhaq & Benjamin, sp. nov., female paratype (AraThom010N 2013 BJ6.1 003). A. Habitus, dorsal view; B. Habitus, ventral view; C. Epigynum, ventral view; D. Epigynum, dorsal view; E. Epigynum, dorsal view, cleared. Scale bars: 1 mm (A, B); 0.2 mm (C–E).
Nyctimus rendang Dhiya’ulhaq & Benjamin, sp. nov. A. Male holotype (AraThom010N 2013 BJ6.1 001), left palp, ventral view; B. ibid., left palp, retrolateral view; C. Female paratype (AraThom010N_2013_BJ6.1_003), epigynum, dorsal view; D. ibid., epigynum, ventral view. Abbreviations: CO = copulatory opening; E = embolus; PE = posterior extension of spermatheca; RTA = retrolateral tibial apophysis; SP = spine; S = spermatheca; TH = tegular hood; VTA = ventral tibial apophysis. Scale bars: 0.2 mm.
Epigynum (Fig. 14C, D, 15C, D): exterior plate with irregular border. Atrium inconspicuous. CO semicircular, posteriorly facing. CD very short. Spermatheca oval, anteriorly with a large bump, posteriorly extended.
Distribution.
Indonesia (Sumatra: Jambi) (Fig. 19).
Remarks.
One specimen shows rounded rather than oval spermathecae (Fig. 14E) and is, thus, tentatively placed here, as no males from the same locality were found.
Nyctimus
saksang
Taxon classificationAnimaliaAraneaeThomisidae
Dhiya’ulhaq & Benjamin sp. nov.
CD696F87-6463-5550-B847-8967D3B46BE2
https://zoobank.org/D731CC5A-13CF-480B-9894-01BCC9A42F9E
Etymology.
The specific epithet refers to a meat-stew dish associated with the Batak people from North Sumatra. The meat is cooked in blood, giving it a dark colouration. The dark colour of Nyctimus saksang Benjamin & Dhiya’ulhaq, sp. nov., is reminiscent of the colour of this dish.
Type material.
**Holotype. ** Indonesia – Sumatra, Jambi Province • ♂; Batang Hari, Hutan Harapan Conservation Area; 02°09'48.9"S, 103°20'04.4"E; elev. 73 m; 6 Jul. 2013; J. Drescher leg.; canopy fogging in rainforest; GOET 2013_HF2.1_AraThom036N_001 (to be transferred to MZB).
Diagnosis.
Males can be easily distinguished from all other congeners by the very wide, axe-shaped RTA which consists of three large lobes (Figs 16C, D, 17).
Description.
Male (holotype 2013_HF2.1_AraThom036N_001; Figs 16, 17). Total length 3.11. Prosoma length 1.50; width 1.25. Opisthosoma length 1.61; width 1.40. Diameter of eyes: AME 0.08; ALE 0.17; PLE 0.14; PME 0.05. Interdistances between eyes: AME–AME 0.40; AME–ALE 0.10; ALE–ALE 0.73; PME–PME 0.38; PME–PLE 0.33; ALE–PLE 0.21; AME–PME 0.22; PLE–PLE 0.95. Leg measurements: leg I 4.20 (1.20, 0.44, 1.05, 0.89, 0.62); leg II 3.74 (1.10, 0.34, 0.91, 0.78, 0.61); leg III 2.78 (0.83, 0.37, 0.67, 0.52, 0.39); leg IV 2.87 (0.88, 0.26, 0.70, 0.60, 0.43). Prosoma cuboid, almost as long as wide, dark reddish brown; surface weakly granulated; laterally adorned with long macrosetae, each arising from a small tubercle. Chelicerae and sternum coloured as prosoma. Endites brown with white tip. Front legs brown; back legs brown except for the white femora. Opisthosoma round, dark brown with paired white bars in the middle that are almost touching each other; anteriorly with a white border; posteriorly with a white patch; dorsally covered with scutum; five large sigillae present, arranged in a triangular formation.
Nyctimus saksang Dhiya’ulhaq & Benjamin, sp. nov. male holotype (AraThom036N 2013 HF2.1 001). A. Habitus, dorsal view; B. Habitus, ventral view; C. Left palp, ventral view; D. Left palp, retrolateral view. Scale bars: 1 mm (A, B); 0.2 mm (C, D).
Nyctimus saksang Dhiya’ulhaq & Benjamin, sp. nov. male holotype (AraThom036N 2013 HF2.1 001). A. Left palp, ventral view; B. Left palp, retrolateral view. Abbreviations: DL = dorsal lobe; E = embolus; ML = medial lobe; RTA = retrolateral tibial apophysis; TH = tegular hood; VTA = ventral tibial apophysis; VL = ventral lobe. Scale bar: 0.2 mm.
Palp (Figs 16C, D, 17): cymbium oval. Embolus elongated, tapering; tip with a slight flange. Tegular hood small. RTA trilobed; ventral lobed finger-shaped; median lobe triangular, with dorsal lobe axe-shaped; dorsal edge slightly folded. VTA hook-shaped, small, attached medially on the ventral lobe of RTA.
Female. Unknown.
Distribution.
Only known from the type locality (Indonesia: Sumatra: Jambi) (Fig. 19).
Zametopina
Taxon classificationAnimaliaAraneaeThomisidae
Simon, 1909
68082F37-4A2F-5D83-A03E-FE75127061E0
Type species.
Zametopina calceata Simon, 1909.
Diagnosis.
Zametopina is similar to Nyctimus in habitus, differing in the presence of a unique denticulate process next to the embolus on the male palp (DP in Fig. 18B). No other oriental thomisid genus possesses this structure. For a detailed diagnosis see Tang et al. (2010).
Zametopina wanliae Lin & Li, 2023, male (MCZ6652). A. Habitus, dorsal view; B. Left palp, ventral view; C. Left palpal tibia, retrolateral view. Abbreviations: DP = denticulate process; E = embolus; RTA = retrolateral tibial apophysis; TH = tegular hood; VTA = ventral tibial apophysis. Scale bars: 0.5 mm (A); 0.2 mm (B, C).
Species composition.
Zametopina calceata Simon, 1909 and Zametopina wanliae Lin & Li, 2023.
Zametopina
calceata
Taxon classificationAnimaliaAraneaeThomisidae
Simon, 1909
6487E2EB-928A-57BE-84E8-F71297687052
Zametopina calceata Simon, 1909: Simon 1909: 123; Tang et al. (2010): 66, figs 1−18.
Type material.
**Holotype. ** Vietnam – Cao Bằng Province • ♂; “Boa Luc, dans Haut-Tonkin, à l’Ouest de Cao-Bang et prés la frontière du Yun-Nan et du Kuang-Si” (probably Bảo Lạc District); 1906 or 1907; de Pelacot leg.; MNHN 22937. Examined.
Diagnosis.
See Lin et al. (2023).
Description.
See Tang et al. (2010).
Distribution.
China (Yunnan), Vietnam (Cao Bằng) (Fig. 19).
Distribution of all described Nyctimus and Zametopina (Araneae: Thomisidae) species. Grey circles surrounding white dots indicate species collected from the same or very closely located sampling points in Brunei, Sabah (Malaysia), and Jambi (Indonesia).
Zametopina
wanliae
Taxon classificationAnimaliaAraneaeThomisidae
Lin & Li, 2023
83804B74-7CD8-5BBE-9E96-196B32D20079
Zametopina wanliae Lin & Li, 2023 in Lin et al. (2023): 79, figs 68A, B, 69I.
Material examined.
Thailand – Chaiyaphum Province • 2♂♂; Tat Ton National Park, Tat Fa Waterfall; 15°56.458'N, 102°5.855'E; elev. 600–700 m; 7. And 8. Mar. 2007; Tawit Jarupan et al. leg.; canopy fogging in rainforest; MCZ MCZ128814, MCZ6652.
Diagnosis.
See Lin et al. (2023).
Description.
Male (MCZ6652; Fig. 18). Total length 2.44. Prosoma length 1.22; width 1.14. Opisthosoma length 1.22; width 1.40. Diameter of eyes: AME 0.09; ALE 0.15; PLE 0.12; PME 0.09. Interdistances between eyes: AME–AME 0.30; AME–ALE 0.13; ALE–ALE 0.69; PME–PME 0.37; PME–PLE 0.35; ALE–PLE 0.23; AME–PME 0.07; PLE–PLE 0.85.
For a complete description, see Lin et al. (2023).
Distribution.
Vietnam (Vĩnh Phúc), Thailand (Chaiyaphum [new record]) (Fig. 19).
Discussion
A key question concerns whether the females assigned to Nyctimus kinabaluensis, sp. nov., are conspecific with the male holotype. These are the only female specimens of Nyctimus included in this study and possess an oval spermatheca and a unique configuration of the copulatory ducts. Interestingly, similar features are observed in the female of Zametopina calceata, as illustrated by Tang et al. (2010), raising the possibility of a male–female mismatch. Nevertheless, we consider such a mix-up unlikely given the strong resemblance in habitus between the males and females. The overall somatic morphology is consistent, and although the genitalia bear superficial similarities to Z. calceata, we regard the differences as sufficient to support the recognition of a distinct species.
This observation leads to a broader taxonomic consideration: whether all species currently attributed to Nyctimus should indeed belong to the same genus. Most species share a remarkably uniform somatic appearance, dark colouration with white markings, a cuboid, granulated prosoma furnished with long setae, and, in males, a prominent scutum. However, genital morphology across the group is highly variable. The type species, N. bistriatus, is morphologically divergent in several key respects: it has widely spaced median eyes, an exceptionally long embolus, a raspy retrolateral tibial apophysis (RTA), and a broad, elongate copulatory duct with a single copulatory opening. These traits set it apart from the other species and raise doubts about the coherence of the genus as currently defined.
All Nyctimus species described here exhibit a weakly granulated, dark-brown prosoma that peaks in height at mid-length and possesses lateral margins lined with long macrosetae, an unusual feature among Oriental Thomisidae. Comparable setae are known only from Tagulis mystacinus Simon, 1895, a species endemic to Sri Lanka, whose closest relatives are found in Africa (Benjamin and Ranasinghe 2019).
All species discussed here possess a highly modified ventral tibial apophysis (VTA) that varies in shape and size. Most known thomisids, informally referred to as “higher thomisids” or the Thomisus clade (Benjamin 2011; Ramírez 2014), represent the bulk of the family’s species diversity. These higher thomisids exhibit a distinct male palpal morphology, characterised by a disk-shaped tegulum (character 11 in Benjamin 2011) and a hook-shaped VTA, which appears to function as a guiding mechanism during hematodochal expansion and rotation (Huber 1994; character 320 in Ramírez 2014).
However, this specific form of VTA is only observed in Nyctimus bistriatus, N. quadripunctatus sp. nov., and Zametopina wanliae. In most of the species discussed here, the VTA is modified in its size, shape, and position on the apical surface of the tibia. In the absence of evidence to the contrary, the most parsimonious assumption is that all these apical ventral apophyses are homologous. If this character system were to be coded for a future phylogenetic matrix, we would propose the following coding scheme: Character #n: VTA, 0, present; 1, absent. Character #n+1: Position, distance from the tibial apex. Character #n+2: Relative size of the VTA.
Among the species discussed, N. mutilloides, sp. nov., is particularly distinctive. The male, for which no female is yet known, displays white dots and bands across the opisthosoma that strongly resemble the aposematic markings of mutillid wasps, whereas other congeners only bear spotted abdominal patterns. Additionally, the male palp is unique: the embolus is beak-shaped and projects prominently from the distal margin of the bulb. To our knowledge, no other thomisids in the Oriental region exhibit such a palp structure. Although this may indicate a distinct lineage within the genus, further material, particularly molecular data and female specimens, is required to test whether this species merits recognition at the genus level.
Clarifying these taxonomic relationships will require molecular phylogenetic data. At present, the evolutionary affinities and higher-level placement of these species remain uncertain. A robust phylogenetic framework would provide essential insights into lineage divergence, character evolution, and genus-level boundaries. In the absence of such data, hypotheses regarding species interrelationships remain speculative and lack firm genetic or evolutionary support. Future studies incorporating both morphological and molecular evidence will be crucial for resolving these questions and establishing a stable classification for this group of crab spiders.
Supplementary Material
XML Treatment for Nyctimus
XML Treatment for Nyctimus bistriatus
XML Treatment for Nyctimus falcatus
XML Treatment for Nyctimus kinabaluensis
XML Treatment for Nyctimus mutilloides
XML Treatment for Nyctimus quadripunctatus
XML Treatment for Nyctimus rendang
XML Treatment for Nyctimus saksang
XML Treatment for Zametopina
XML Treatment for Zametopina calceata
XML Treatment for Zametopina wanliae
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Benjamin SP (2011) Phylogenetics and comparative morphology of crab spiders (Araneae: Dionycha, Thomisidae).Zootaxa 3080(1): 1–108. 10.11646/zootaxa.3080.1.1 · doi ↗
- 2Benjamin SP Ranasinghe UGSL (2019) Redescription of Tagulis granulosus (Araneae: Thomisidae) from Sierra Leone.Arachnology 18(1): 22–23. 10.13156/arac.2018.18.1.22 · doi ↗
- 3Dhiya’ulhaq NU Ramos Gutierrez DE Nazarreta R Lia M Pakpahan B Buchori D Scheu S Drescher J (2024) Spiders of Jambi: a Guide to the EF For TS Collection. e-Publishing Penerbrit BRIN, Jakarta. 10.55981/brin.824 · doi ↗
- 4Drescher J Rembold K Allen K Beckschäfer P Buchori D Clough Y Faust H Fauzi AM Gunawan D Hertel D Irawan B Jaya INS Klarner B Kleinn C Knohl A Kotowska MM Krashevska V Krishna V Leuschner C Lorenz W Meijide A Melati D Nomura MPérez-Cruzado C Qaim M Siregar IT Steinebach S Tjoa A Tscharntke T Wick B Wiegand K Kreft H Scheu S (2016) Ecological and socio-economic functions across tropical land use systems after rainforest conversion. Philosophical Transactions of the Royal Society of London.Series B, Biological Sciences 371(1694): 20150275. 10.1098/rstb.2015.027 · doi ↗ · pubmed ↗
- 5Huber BA (1994) Genital bulb muscles in entelegyne spiders.The Journal of Arachnology 22: 75–76.
- 6Ileperuma Arachchi IS Benjamin SP (2019) Twigs that are not twigs: Phylogenetic placement of crab spiders of the genus Tmarus of Sri Lanka with comments on the higher level phylogeny of Thomisidae.Invertebrate Systematics 33: 575–595.
- 7Lehtinen PT (2016) Significance of oriental taxa in phylogeny of crab spiders (Thomisidae s.lat. and Stiphropodidae).Indian Journal of Arachnology 5: 143–171.
- 8Lin YJ Li SQ Pham DS (2023) Taxonomic notes on some spider species (Arachnida: Araneae) from China and Vietnam.Zoological Systematics 48(1): 1–99. 10.11865/zs.2023101 · doi ↗