A new cave-dwelling species of Trimma (Teleostei, Gobiidae) from the Red Sea, with notes on Red Sea endemism in Trimma spp

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1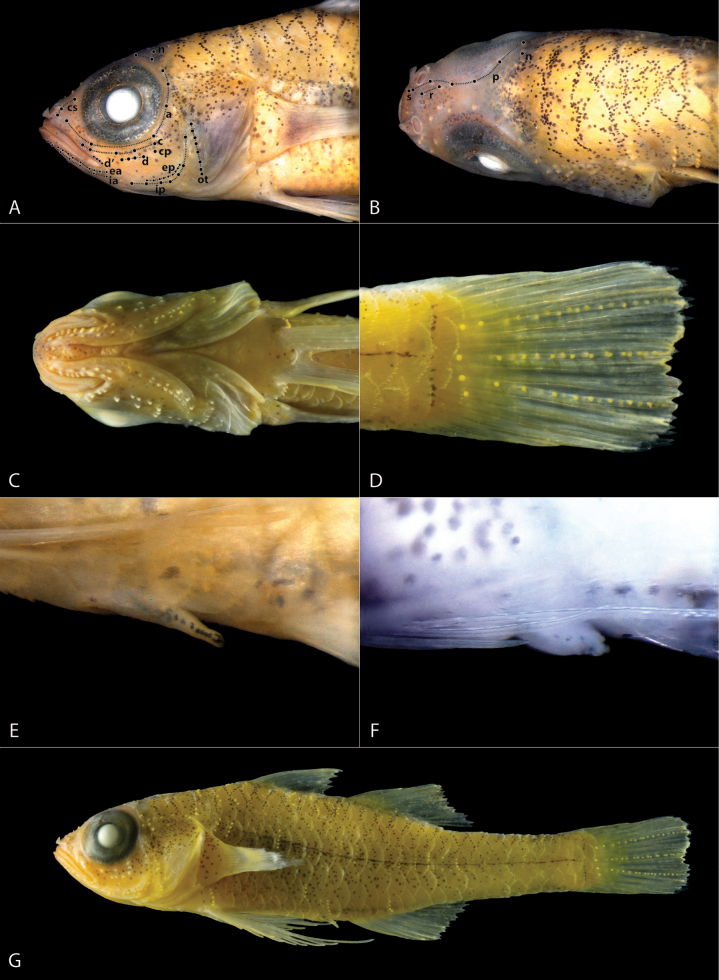 Figure 2
Figure 2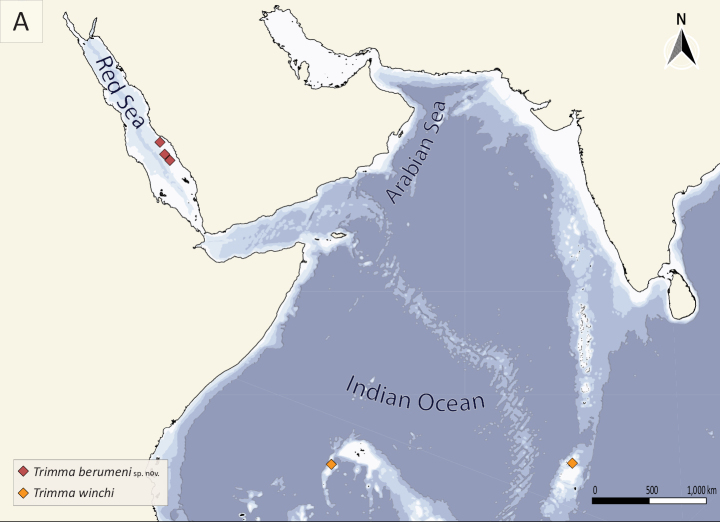 Figure 3
Figure 3 Figure 4
Figure 4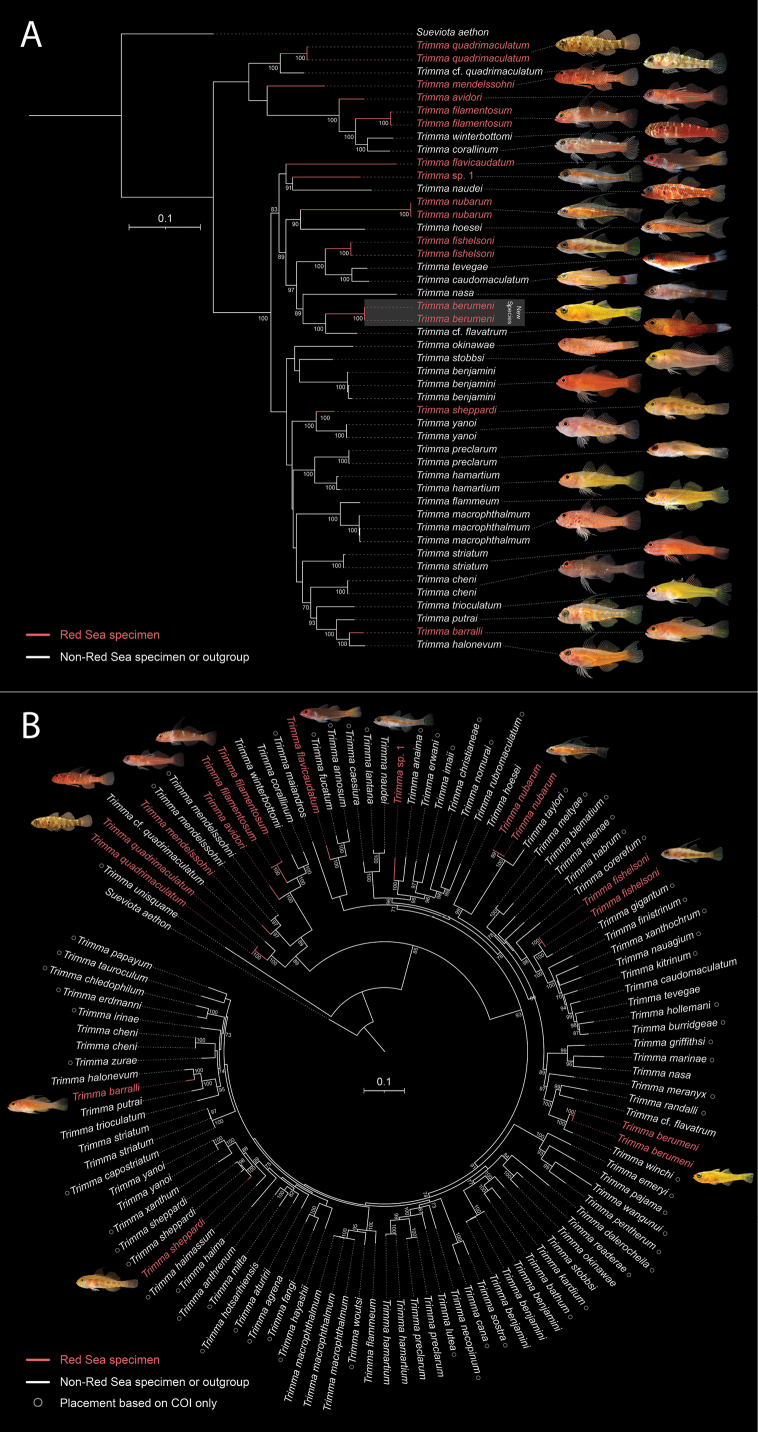 Figure 5
Figure 5|
| ||
|---|---|---|
| Geographic range | Farasan Banks, southern Red Sea | Seychelles and Chagos Archipelago |
| Dorsal fin | VI + I,7 | VI + I,8 |
| Elongate dorsal spines | None; second spine longest reaching to base of second dorsal-fin spine when adpressed | Second spine elongate, reaching posteriorly just beyond base of last dorsal-fin ray when adpressed (absent in Seychelles specimens) |
| Second dorsal-fin length | Last ray not elongate, reaching posteriorly 23–30% of caudal peduncle length | Last ray elongate, reaching posteriorly two-thirds of caudal peduncle length |
| Anal-fin length | Last ray not elongate, reaching posteriorly 23–30% of caudal peduncle length | Last ray elongate, reaching posteriorly two-thirds of caudal peduncle length |
| Dorsal-fin color (alive) | Black longitudinal basal stripe composed of large melanophores with scattered iridocytes; single yellow longitudinal band above it; distal third hyaline | Black longitudinal basal stripe composed of large melanophores with scattered iridocytes; a central yellow longitudinal band, then a narrow hyaline band; distal quarter yellow |
| Anal-fin color (alive) | No yellow pigmentation, mostly translucent with red, white, and black chromatophores | Mostly yellow, with basal band of melanophores and iridocytes |
| Caudal-fin color (alive) | Yellow with translucent dorsal and ventral margins | Yellow throughout with distal margin of melanophores |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFish Biology and Ecology Studies · Identification and Quantification in Food · Ichthyology and Marine Biology
Introduction
The gobiid genus Trimma Jordan & Seale, 1906 is one of the most speciose genera of coral reef-associated fishes in the Indo-Pacific, with over 110 valid described species. Commonly known as pygmygobies due to their small body size (typically <30 mm SL), members of this genus can be found on most Indo-Pacific coral reefs. Along with the similarly diverse and small-bodied Eviota Jenkins, 1903, they frequently represent one of the most abundant gobiids on hard substrates of coral reefs (Winterbottom 2019). Like Eviota, species of Trimma frequently show strong site fidelity and microhabitat specialization, likely contributing to their rapid diversification. New species are being discovered frequently, and molecular analyses suggest that many widespread species may represent complexes of multiple cryptic species with restricted geographic distributions (Winterbottom et al. 2020).
The Red Sea harbors a distinctive assemblage of Trimma species, with most being endemic to the region (Bogorodsky and Randall 2019). Most of these species show strong associations with specific reef zones or microhabitats. Trimma flavicaudatum (Goren, 1982) and T. avidori (Goren, 1978) often represent the dominant cryptobenthic species on many Red Sea reefs, in some cases with dozens of individuals per square meter (Coker et al. 2018). However, their niche overlap is limited, with typically only one of the species dominating in each habitat. Some species, including T. barralli Winterbottom, 1995 and T. sheppardi Winterbottom, 1984, primarily inhabit depths of at least 30 m, while others, such as T. mendelssohni (Goren, 1978), are common in the upper 10 m of the reef. Associations with caves, which are common for Trimma species throughout the Indo-Pacific, are also present in the Red Sea. Both T. fishelsoni Goren, 1985 and T. nubarum Winterbottom, Bogorodsky & Alpermann, 2023 live in cave-associated groups. The former prefers small groups of low density, while the latter can be observed in dense schools of upwards of 30 individuals, hovering above the substrate near the entrances of caves (pers. obs. of first author).
During biodiversity surveys of offshore reefs in the Farasan Banks region of the Saudi Arabian Red Sea, we discovered a distinctive, yellow-colored species of Trimma inhabiting cave systems at depths between 15–30 m. While often co-occurring with other cave-associated Trimma species, this taxon was typically restricted to areas slightly deeper inside the cave and more closely associated with the cave substrate. Herein we describe this new species based on morphological and molecular evidence and discuss its relationship to other members of the genus, particularly its apparent sister species T. winchiWinterbottom 1984 from the western Indian Ocean.
Materials and methods
Morphological measurements and meristic counts follow standard protocols established by Richard Winterbottom to ensure comparability with previous work (Winterbottom 2019; Winterbottom et al. 2023 and references cited therein). Likewise, the format of the diagnosis and description follows the sequence of characters in the key to the genus (Winterbottom 2019) and other recent species descriptions (Winterbottom et al. 2023). We documented cephalic sensory papillae following established row naming conventions (Winterbottom 2011; Winterbottom and Erdmann 2015). Character ranges are reported as minimum – holotype – maximum, followed by the mean and number of specimens in parentheses. Values of the holotype are presented in bold.
Specimens were collected from cave habitats at depths of 15–30 m using clove oil anesthetic and hand nets. Live and fresh coloration was documented through photographs taken immediately after collection and during in situ observations. Ex-situ photographs were taken in an upright photo tank with a Nikon Z7 camera body, Nikon Z MC 105 mm f/2.8 macro lens, and a Godox MF12 Macro flash. To enhance visibility of key taxonomic features, particularly the cephalic sensory papillae, specimens were temporarily stained with cyanine blue 5R (acid blue 113).
For molecular analyses, we extracted genomic DNA from tissue samples using the Qiagen DNeasy Blood & Tissue Kit. A region of the cytochrome c oxidase subunit I (COI) gene was amplified using the primer sets FishF2 and FishR2 (Ward et al. 2005) and GOBYL6468 and GOBYH7696 (Thacker 2003). Resulting sequences were then cleaned and aligned in Geneious Prime 2025.0.3 to produce a 648-bp alignment. Pairwise genetic distances between species were calculated using MEGA11 using the Kimura 2-parameter (K2P) model (Kimura 1980; Tamura et al. 2021) on the COI alignment. We also amplified three additional loci from the same specimens: mitochondrial 16S rRNA using primers 16SarL and 16SbrH (Palumbi et al. 1991), nuclear S7 ribosomal protein intron 1 (S7I1) using primers S7I1F and S7I1R (Chow and Hazama 1998), and nuclear Protease III (Ptr) using primers PtrF2 and PtrR2 (Yamada et al. 2009). Ptr was included because it can help resolve deeper splits and has been used in the similarly speciose genus Eviota (Tornabene et al. 2013).
The final concatenated alignment consisted of 648 bp COI, 588 bp Ptr, 904 bp S7I1, and 608 bp 16S. The multi-locus dataset comprised specimens from our studies and collections, including all described Red Sea Trimma, 21 non-Red Sea Trimma, and two likely undescribed taxa: one Red Sea species similar to T. anaima Winterbottom, 2000 and one Madagascar species similar to T. quadrimaculatum Hoese, Bogorodsky & Mal, 2015. These specimens are currently being worked on by the authors, but they were included here due to their relevance to Red Sea endemism in Trimma spp.
A maximum-likelihood tree was inferred using W-IQ-TREE (Trifinopoulos et al. 2016) via the web interface at http://iqtree.cibiv.univie.ac.at/. Protein-coding genes (COI and Ptr) were partitioned by codon position; all loci were also partitioned by gene. The best-fit substitution model was automatically selected. Branch support was assessed with 1000 ultrafast bootstrap replicates. Sueviota aethon Peinemann, Pombo-Ayora & Tornabene, 2024 was included as an outgroup to root the tree. The phylogenetic tree was visualized using Interactive Tree Of Life (iTOL) v6 (Letunic and Bork 2024) and finalized in Adobe Illustrator 29.2.1.
In addition to the multi-locus dataset, we constructed a supermatrix that combines the full concatenated data for our specimens (COI, Ptr, S7I1, 16S) with COI-only terminals for Trimma species not represented in our collection. We queried the Barcode of Life Data System (BOLD) for all such species, selecting one representative COI sequence per species, with preference for longer sequences and for records identified by recognized experts. These public COI sequences were aligned to the COI partition of our concatenated dataset; the remaining three loci for those added taxa were coded as missing data. This preserves the higher signal of our multi-locus sampling while placing it in the context of the majority of described species of Trimma. Supp. material 1 lists, for each added species, the species name as used in the tree, the BOLD Process ID and, where available, the corresponding GenBank accession number, the name of the specimen identifier, and the collection locality. Supp. material 2 lists information for specimens in our collection used for the multi-locus dataset and the COI and 16S GenBank accession numbers generated for this study. The final supermatrix tree included all 33 Trimma species from our multi-locus dataset plus 60 additional COI-only species from BOLD, for a total of 93 Trimma species.
Type specimens have been catalogued and deposited in the ichthyology collection of the Royal Ontario Museum (ROM). All fieldwork was conducted under approval 20IAUCUC05 issued by the KAUST Institutional Animal Care and Use Committee (IACUC).
Results
Trimma
berumeni
sp. nov.
Taxon classificationAnimaliaGobiiformesGobiidae
89DBA89D-CCF6-54A8-AD75-6C1DB12FC0C0
https://zoobank.org/9759ABBF-2807-4C55-9925-7598731B789B
Figs 1, 2 Mikey’s golden pygmygoby
Type locality.
Saudi Arabia, Red Sea: Farasan Banks, 18.5041°N, 40.6606°E.
Type material.
Holotype • ROM 116871, 13.6 mm SL male, Red Sea, Saudi Arabia, Farasan Banks, 18.5041°N, 40.6606°E, inside cave of exposed reef wall, 27 m depth, collected with clove oil and hand net, V. N. Peinemann, 5 June 2024.
Paratypes • ROM 116872, n = 2, 13.3 mm SL male, Red Sea, Saudi Arabia, Farasan Banks, 18.5041°N, 40.6606°E, inside cave of exposed reef wall, 27 m and 15 m depth respectively, collected with holotype • ROM 116873, 13.8 mm SL male, Red Sea, Saudi Arabia, Farasan Banks, 18.8581°N, 40.3788°E, inside cave of exposed reef wall, 15 m depth, V. N. Peinemann, 6 June 2024 • ROM 116874, n = 3, 10.7 mm SL female, 10.9 mm SL male, and 10.7 mm SL male, Red Sea, Saudi Arabia, Farasan Banks, 19.8375°N, 39.9297°E, inside cave of exposed reef wall, 30 m depth, V. N. Peinemann, 19 May 2022.
Diagnosis.
A species of Trimma distinguished by the following combination of characters: predorsal midline with 7–8 scales; fifth pelvic-fin ray unbranched and 40–42% length of fourth ray; cheek and opercle scaleless; all pectoral-fin rays unbranched; body bright yellow throughout and without bars in life; dorsal fin VI + I,7; no elongate spines in dorsal fin; bony interorbital width 81–84% of pupil diameter; iris golden-yellow with melanophores except for golden ring around pupil and inverted golden triangle extending from ventral margin of pupil; fins translucent with yellow-green longitudinal band in central third of dorsal fins; a thin black midlateral line from above pectoral-fin base to caudal-fin base, visible in life and in preservative.
Description.
Based on holotype and 6 paratypes 10.7–13.8 mm SL. Dorsal fin VI + I,7, second spine longest but not elongate extending to base of second dorsal-fin spine when adpressed; all rays of second dorsal fin branched; second dorsal fin not elongate, reaching posteriorly 23–26–30 (27%, 7) of caudal peduncle length; anal fin I, 8, all rays branched; anal fin not elongate, reaching posteriorly 23–30 (27%, 7) of caudal peduncle length; pectoral-fin rays 14–15, all unbranched; pectoral-fin reaching posteriorly to vertical above urogenital papilla; pelvic fin I, 5, fifth ray unbranched and 40–42 (41%, 7) length of fourth ray, which reaches posteriorly to between base of first and third anal-fin ray, pelvic rays 1 to 4 with one branching point each; basal membrane approximately 9–10% of fourth pelvic-fin ray; no fraenum; caudal fin with 2 dorsal and 2–3 ventral segmented unbranched rays, and 6 dorsal and 5 ventral segmented branched rays; caudal fin with four vertically aligned papillae at base and three rows of 9–10 papillae each extending from the vertically aligned papillae to posterior margin of caudal fin (Fig. 2D); male urogenital papilla elongated and narrow (Fig. 2E; female urogenital papilla short and bulbous (Fig. 2F).
Lateral scales 24; anterior transverse scales 7–8, posterior transverse scales 7–8; cheek and opercle scaleless; 7–8 scales on predorsal midline; body scales ctenoid; circumpeduncular scales 12; scale rows in ventral midline between base of last anal-fin ray and first procurrent caudal-fin ray 7–8.
Gill opening extending anteroventrally to between middle and posterior third of pupil (Fig. 2C); anterior naris tubular reaching anteriorly across upper lip to posterior margin of lower lip; posterior naris oval and pore-like with raised rim, separated from bony front of orbit by 1.6–2 times its diameter (Fig. 2B); bony interorbital width 81–84% of pupil diameter (82%, 7); no dermal ridge on midline of nape extending anteriorly from origin of first dorsal fin.
Caudal peduncle depth as percentage of caudal peduncle length 30–41 (37.2%, 7); head length as percentage of SL 27–29–31 (29.3%, 7); horizontal eye diameter 40–45 (43.1%, 7); snout length 16–20 (18.9%, 7); and upper-jaw length 32–33 (32.6%, 7) as percentage of head length
Number of papillae in each row (Fig. 2A): a = 6 (4); b = 1–2 (5); c = 5 (5); d = 3–4 (5); d’ = 3–4 (5); p = 6 (5); e-anterior = 7–9 (5); e-posterior = 7–9 (5); i-anterior = 6–7 (5); i-posterior = 7 (5); cs” = 3 (5); ot = 7–8 (4); oi = 3–4 (4); u = 4 (3); cp = 1 (5); n = 1 (5); f = 2–3 (5).
Color pattern, live (Fig. 1). Body bright yellow throughout. Head primarily yellow with orange coloration around the jaws, snout, interorbital region and below the ventral margin of the eye. Individuals can activate five translucent whitish bars along dorsal margin of body, from above the pectoral fin base to the caudal peduncle, sometimes visible in situ. Head and body with numerous small melanophores scattered throughout, running along scale pockets in the dorsal third of body, and scattered without patterns below the midlateral line. Specimens smaller than 11 mm SL with few melanophores on ventral half of body. A thin black midlateral line running along body from above pectoral-fin base to caudal-fin base. Iris golden-yellow, peppered with melanophores, except in a golden ring surrounding the pupil and an inverted golden triangle extending from ventral margin of the pupil. Dorsal fins translucent with dense red, white, and black chromatophores along basal third, a yellow-green longitudinal band running along central third, and sparse red, white, and black chromatophores along distal third. Anal fin mostly translucent with red, white, and black chromatophores, without yellow-green pigmentation. Dorsal and ventral fifths of caudal fin translucent with scattered red and white chromatophores, central section of caudal fin yellow to yellow-green. Pectoral fins and pelvic fins hyaline.
Freshly collected Trimma berumeni sp. nov., holotype, ROM 116871. Photograph: V. N. Peinemann.
Close-up of key features of Trimma berumeni sp. nov., preserved in ethanol, ROM 116871 (A, B, C, D, E, G), ROM 116874 (F). A. Lateral view of head showing cephalic sensory papillae, specimen stained with cyanine blue, papillae highlighted with black dots, papilla rows connected by dotted lines and labeled accordingly; B. Dorsal view of head, showing cephalic sensory papillae and nasal apparatus, specimen stained with cyanine blue, papillae highlighted with black dots, papilla rows connected by dotted lines and labeled accordingly; C. Ventral view of head, showing cephalic sensory papillae, gill opening, and pelvic-fin base; D. Caudal fin with sensory papillae; E. Male urogenital papilla; F. Female urogenital papilla, specimen stained with cyanine blue; G. Lateral view of whole fish (preserved in 75% ethanol).
Color pattern, preserved (Fig. 2G). Head and body primarily light yellow in large and recently preserved specimens, slowly fading to white in small specimens older than a year. Melanophores remain scattered throughout. A thin black midlateral line just under scales, extending from above pectoral-fin base to caudal-fin base. Iris densely packed with melanophores, appearing mostly black. Dorsal and anal fins hyaline with melanophores scattered in the posterior third of each fin, growing denser towards the posterior margin of the fin. Pectoral, pelvic, and caudal fins hyaline.
Etymology.
Named in honor of Michael L. Berumen in recognition of his substantial contributions to our understanding of the ecology and biodiversity of Red Sea coral reefs. Mikey’s golden pygmygoby is suggested as the common name.
Distribution and habitat.
Trimma berumeni sp. nov. inhabits caves of exposed offshore reefs, where it moves along the surface of cave roofs and walls in small groups of three to ten individuals (occasionally it is also solitary). It is typically found deep within caves, rarely seen within the first 80 cm of a cave entrance. The habitat is similar to that of its sister species, Trimma winchi, in the Seychelles (Ryan Daly pers. comm.).
Specimens were collected at depths between 15 and 30 m. While only two specimens were observed shallower than 20 m, the species is relatively common in caves at 30 m. Our collections and surveys were limited to 30 m, but it is likely that the species extends to greater depths. We observed this species exclusively on reefs with steep walls that extend well into the mesophotic.
The species is present throughout much of the Farasan Banks in the southeastern Red Sea (Fig. 3). Despite extensive collections in similar habitats along the Saudi Arabian coastline north of the Farasan Banks, from Jeddah to Tiran, we have not observed this species anywhere but the Farasan Banks. It is possible that the species is endemic to the southern half of the Red Sea.
Map showing collection localities of Trimma berumeni sp. nov. (red diamonds; present study) and its sister species T. winchi (orange diamonds; Winterbottom 2019) in the Red Sea and Indian Ocean.
Comparisons
In the key to the genus (Winterbottom 2019), T. berumeni keys out to couplet 71 but does not match either of the two species in the couplet. Trimma berumeni differs from both in having 7 dorsal fin rays (vs 9 in Trimma imaii Suzuki & Senou, 2009 and 8 in Trimma matsunoi Suzuki Sakaue & Senou, 2012). It additionally differs from T. imaii in having a bony interorbital width wider than 80% of pupil width (vs 40% in T. imaii) and from T. matsunoi in having 7–8 predorsal midline scales (vs 6 in T. matsunoi). The general body, fin, and eye color is also clearly distinct in each species.
In overall morphology, the T. berumeni is most similar to T. winchi, which is currently known from the Seychelles and the Chagos Archipelago (Fig. 4A). They share a uniform yellow body color, have a similar habitat, and a geographically close but non-overlapping distribution. The COI sequences of T. berumeni and T. winchi differ by a K2P distance of 6.4% (see Suppl. material 3 for K2P distances of Red Sea Trimma and sister taxa in Fig. 5B). Several traits can be used to separate the two species. These are summarized in Table 1. Notably, the two Chagos (type locality) male specimens of T. winchi have an elongate dorsal spine, which is absent in Ryan Daly’s photographs of T. winchi from the Seychelles. It has not been evaluated whether this represents a distinct species or intraspecific variation. In Table 1 we follow the original description in listing dorsal-spine elongation for T. winchi, but the variation warrants further investigation.
Comparison of species similar to Trimma berumeni sp. nov. in the Red Sea and Indian Ocean. A. T. winchi, holotype, ROM 41477, fresh, Salomon Atoll, Chagos Archipelago; B. T. fishelsoni, fresh, Straits of Tiran (northern Red Sea / Gulf of Aqaba), Saudi Arabia; C. T. fishelsoni, live, Obstruction Reef (Thuwal, central Red Sea), Saudi Arabia. Photographs: A. by R. Winterbottom; B, C. by V. N. Peinemann.
Phylogenetic placement of Trimma berumeni sp. nov. and Red Sea Trimma diversity. A. Maximum-likelihood tree from a concatenated alignment of COI, 16S, Ptr, and S7I1 for 33 species (all Red Sea Trimma plus selected Indo-Pacific relatives). Bootstrap values >70% are shown for internal nodes (1,000 ultrafast bootstrap replicates). Sueviota aethon is the outgroup. Scale bar represents 0.1 substitutions per site. Red branches and labels denote Red Sea specimens; among Red Sea Trimma, all species except T. mendelssohni and T. sheppardi are considered endemic species. Photographs correspond to the species listed at the tips, in order from top to bottom (photo credits: Red Sea T. quadrimaculatum S. V. Bogorodsky; all others V. N. Peinemann & L. Pombo-Ayora). Images are of the sequenced specimen or a conspecific from the same locality. B. Maximum-likelihood tree from a supermatrix (concatenated sequences from panel A + 60 additional species with COI only; total 93 species). Bootstrap values >70% are shown for internal nodes (1,000 ultrafast bootstrap replicates). Scale bar represents 0.1 substitutions per site. Red labels indicate Red Sea taxa; open circles mark tips placed using COI only. Photos are shown only for Red Sea species. Sequence sources, vouchers, and GenBank/BOLD accession numbers for all taxa shown are listed in Suppl. material 1 (COI-only taxa) and Supp. material 2 (multi-locus taxa).
Table 1.: Comparison of key characters between Trimma berumeni sp. nov. and T. winchi. Data for T. winchi based on Winterbottom (1984), Winterbottom (2019), and examination of a photograph of holotype.
Within the Red Sea, T. fishelsoni bears some similarity to T. berumeni. Both species have a primarily yellow body color, a yellow longitudinal band along the dorsal fins, and a primarily yellow caudal fin. Both species also have an overlapping distribution in the central to southern Red Sea and are associated with caves. Trimma berumeni can be distinguished from T. fishelsoni by having no elongate dorsal spines (vs second spine elongate and filamentous in T. fishelsoni), 7–8 predorsal midline scales (vs 9–14 in T. fishelsoni), scaleless cheek and operculum (vs scaled in T. fishelsoni), a basal membrane connecting the fifth pelvic fin rays (vs absent in T. fishelsoni). The live color of T. berumeni differs by the body being bright yellow throughout (vs yellow with a white to purple stripe above midlateral line in T. fishelsoni), head yellow throughout (vs primarily pale to pink in lower jaw and ventrally of eye in T. fishelsoni), no longitudinal yellow band on anal fin (vs usually present in T. fishelsoni), no blotches at caudal fin base (vs two large vertically aligned yellow-orange blotches at caudal fin base in T. fishelsoni), and no lines across iris (vs oblique blue line with orange-red margins across upper edge of pupil in T. fishelsoni).
Bogorodsky et al. (2016) extended the known range of T. fishelsoni from the northern Red Sea to the Farasan Banks, while noting some morphological differences between the populations. The southern population has no branching pectoral fin rays while northern ones have some. Southern populations lack the four saddle-like dorsal blotches described for Gulf of Aqaba specimens (more apparent in preservative) by Goren (1984). Gulf of Aqaba specimens (Fig. 4B) also appear to have a yellow patch at the posterior margin of the jaw that is absent or less distinctive in southern populations. We have collected specimens from the central Red Sea with a mostly pale dorsal body color and 5 or 7 dusky yellow dorsal blotches (either a single large or two smaller blotches are present under each dorsal fin) that were apparent in live, freshly dead, and preserved individuals (Fig. 4C). These individuals have no branching pectoral fin rays and a much fainter yellow color on the anal fin. While further analyses and additional specimens are needed to determine the nature of this variation between T. fishelsoni populations, the differences to T. berumeni discussed above are consistent across populations.
Discussion
The Red Sea is a peripheral sea characterized by high levels of endemism shaped by its geographic isolation and distinct environmental conditions. The Bab-el-Mandeb Strait limits gene flow between the Red Sea and the Indian Ocean, while upwelling systems along the eastern Somali and southern Arabian coasts create ecological barriers that further restrict dispersal. These factors have contributed to an exceptionally high level of endemism in Red Sea fishes (DiBattista et al. 2016a, 2016b; Bogorodsky and Randall 2019). New putative endemic species continue to be discovered frequently, including in the same regions and cave habitats as the new Trimma berumeni (Winterbottom et al. 2023; Nunes Peinemann et al. 2024).
With the description of Trimma berumeni, the number of described Trimma species recorded from the Red Sea rises to 10, of which eight are currently considered endemic species. Two species, T. mendelssohni and T. sheppardi, are currently recognized as widespread but exhibit relatively deep genetic divergence between their Red Sea and non-Red Sea populations (4–10% K2P distance). While further study is needed, these divergences suggest that the Red Sea lineages may represent cryptic endemic species rather than truly widespread taxa. In comparison, the K2P distance between T. berumeni and T. winchi is 6.4%. Geographically restricted haplogroups and potential cryptic speciation are well documented in Trimma spp. (Winterbottom et al. 2014, 2020, 2023). For completeness, we also included two putatively undescribed taxa, T. cf. quadrimaculatum (Madagascar) and a Red Sea T. sp. 1 similar to T. anaima, which each form Red Sea/non-Red Sea sister pairs in our trees (Fig. 5): the former to the Red Sea T. quadrimaculatum lineage (13.5% COIK2P) and the latter to T. anaima outside the Red Sea (15.6% COIK2P). They are included solely for context and are not treated further in this paper.
Our 93-species maximum-likelihood tree places T. berumeni in a well-supported sister pairing with Trimma winchi (Fig. 5B). Although the placement of T. winchi is based solely on the COI gene (and it was not included in the multi-locus analysis; Fig. 5A), this relationship is further supported by similarities in their morphology and habitat preferences. The two species maintain distinct but adjacent geographic ranges, with T. berumeni found in the central to southern Red Sea and T. winchi in the western Indian Ocean. More broadly, our analyses suggest that most Red Sea Trimma have a sister taxon located outside the Red Sea (Fig. 5). This pattern is consistent with multiple colonization events rather than a single evolutionary radiation within the basin. Such phylogenetic overdispersion, where endemic species are scattered across various Indo-Pacific lineages, mirrors patterns observed in other Red Sea reef fish families (DiBattista et al. 2016b).
Despite the high number of endemics, in situ diversification within the Red Sea appears to be uncommon in fishes, although there is some evidence of the Red Sea exporting diversity to the Indian Ocean (Bowen et al. 2013; DiBattista et al. 2013). In Trimma, COIK2P distances (Supp. material 3) suggest Red Sea endemism could be as high as 100%, yet our analyses provide no strong evidence for diversification occurring within the basin. The distribution of Red Sea taxa across otherwise Indo-Pacific lineages is parsimoniously explained by repeated, independent colonizations of the Red Sea; while an older in-basin event followed by speciation outside the Red Sea cannot be excluded, the current evidence does not require it.
Interpretation of our phylogenetic results warrants caution given data limitations. Although Fig. 5A incorporates multiple loci and Fig. 5B broadens taxon coverage, most non-Red Sea species are represented by COI alone. Because the supermatrix is anchored on COI, deeper placements of COI-only terminals should be regarded as provisional. Accordingly, we use the expanded tree primarily to frame Red Sea/non-Red Sea sister pairings rather than to infer deeper relationships among Indo-Pacific lineages. Denser taxon sampling with additional loci may provide valuable insights into the origin of Red Sea Trimma and Trimma evolution overall.
Supplementary Material
XML Treatment for Trimma berumeni
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bogorodsky SV Randall JE (2019) Endemic fishes of the Red Sea. In: Rasul NMA Stewart ICF (Eds) Oceanographic and Biological Aspects of the Red Sea.Springer Oceanography. Springer International Publishing, Cham, 239–265. 10.1007/978-3-319-99417-8_14 · doi ↗
- 2Bogorodsky SV Suzuki T Mal AO (2016) Description of a new species of Priolepis (Perciformes: Gobiidae) from the Red Sea, a new record of Priolepis compita, and a distributional range extension of Trimma fishelsoni. Zootaxa 4150(2): 168–184. 10.11646/zootaxa.4150.2.527515653 · doi ↗ · pubmed ↗
- 3Bowen BW Rocha LA Toonen RJ Karl SA (2013) The origins of tropical marine biodiversity.Trends in Ecology & Evolution 28(6): 359–366. 10.1016/j.tree.2013.01.01823453048 · doi ↗ · pubmed ↗
- 4Chow S Hazama K (1998) Universal PCR primers for S 7 ribosomal protein gene introns in fish.Molecular Ecology 7: 1247–1263.9734083 · pubmed ↗
- 5Coker DJ Di Battista JD Sinclair-Taylor TH Berumen ML (2018) Spatial patterns of cryptobenthic coral-reef fishes in the Red Sea.Coral Reefs 37(1): 193–199. 10.1007/s 00338-017-1647-9 · doi ↗
- 6Di Battista JD Berumen ML Gaither MR Rocha LA Eble JA Choat JH Craig MT Skillings DJ Bowen BW (2013) After continents divide: Comparative phylogeography of reef fishes from the Red Sea and Indian Ocean.Journal of Biogeography 40(6): 1170–1181. 10.1111/jbi.12068 · doi ↗
- 7Di Battista JD Howard Choat J Gaither MR Hobbs J-PA Lozano-Cortés DF Myers RF Paulay G Rocha LA Toonen RJ Westneat MW Berumen ML (2016 a) On the origin of endemic species in the Red Sea.Journal of Biogeography 43(1): 13–30. 10.1111/jbi.12631 · doi ↗
- 8Di Battista JD Roberts MB Bouwmeester J Bowen BW Coker DJ Lozano-Cortés DF Howard Choat J Gaither MR Hobbs J-PA Khalil MT Kochzius M Myers RF Paulay G Robitzch VSN Saenz-Agudelo P Salas E Sinclair-Taylor TH Toonen RJ Westneat MW Williams ST Berumen ML (2016 b) A review of contemporary patterns of endemism for shallow water reef fauna in the Red Sea.Journal of Biogeography 43(3): 423–439. 10.1111/jbi.12649 · doi ↗