Exploring the associations between preen oil bacterial, chemical and proteomic profiles of passerines
I. Maureen Baars, Jakub Mrázek, Jakub Kreisinger, Ivan Mikšík, Maurine W. Dietz, Joana Falcao Salles, B. Irene Tieleman, Veronika Gvoždíková Javůrková

TL;DR
This study explores the connections between bacteria, chemicals, and proteins in preen oil from eight bird species, revealing possible roles in producing signals and antimicrobial compounds.
Contribution
First characterization of preen oil chemical and proteomic profiles in passerines and exploration of bacteriome contributions.
Findings
Preen oil profiles varied among birds, with bacteria like Streptococcus and Corynebacterium being common.
Chemical profiles included alcohols, ketones, and carboxylic acids, while proteomic functions included proteolysis and stress response.
Riparian species showed higher chemical diversity and more mVOC-producing bacteria, suggesting habitat-specific adaptations.
Abstract
Preen gland bacteria are thought to be the key producers of preen oil components such as chemosignalling molecules including volatile organic compounds (VOCs) and antimicrobial compounds including peptides and antimicrobial VOCs. However, data on the preen oil bacteriome and chemical composition are limited to a small subset of bird species, and the presence of antimicrobial peptides is largely unexplored. Here, we performed an exploratory study to characterize, for the first time, the preen oil chemical and proteomic profiles and to explore the possible contribution of the bacteriome to the production of preen oil VOCs and antimicrobial peptides (bacteriocins) in eight passerine species, each represented by a single individual. Preen oil bacteriome, chemical and proteomic profiles varied among birds. The bacterial profiles were dominated by the genera Streptococcus, Lactococcus,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
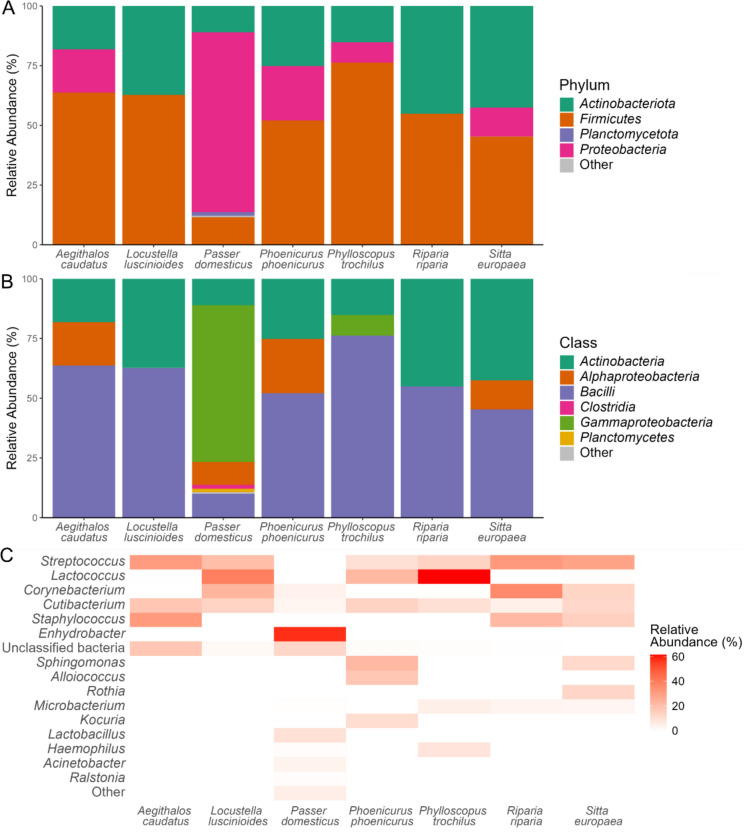 Figure 1
Figure 1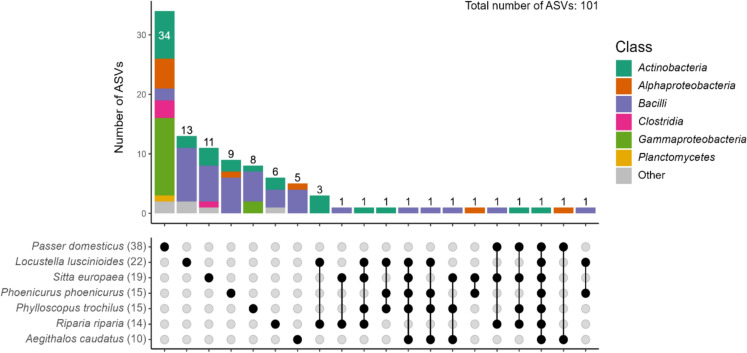 Figure 2
Figure 2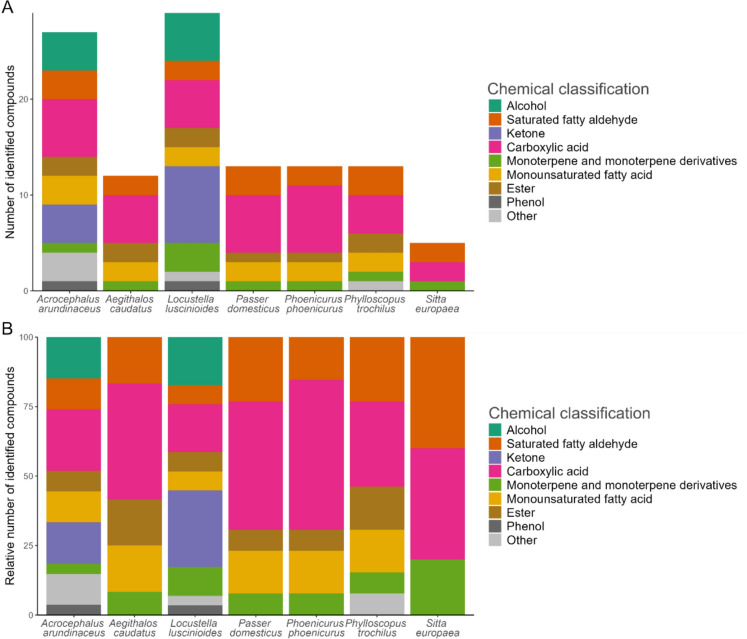 Figure 3
Figure 3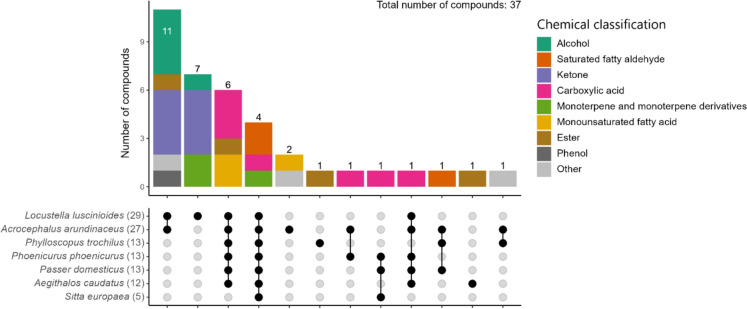 Figure 4
Figure 4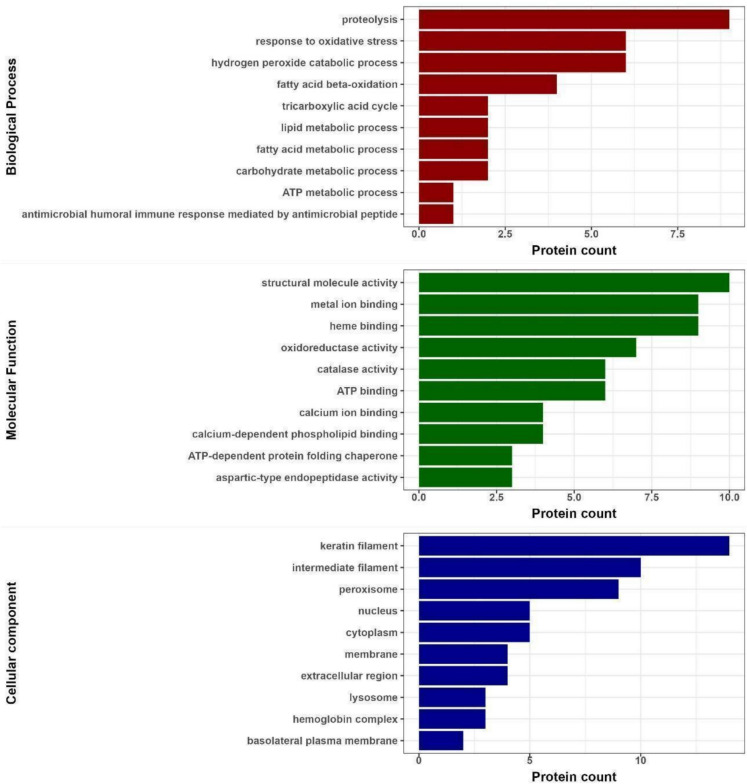 Figure 5
Figure 5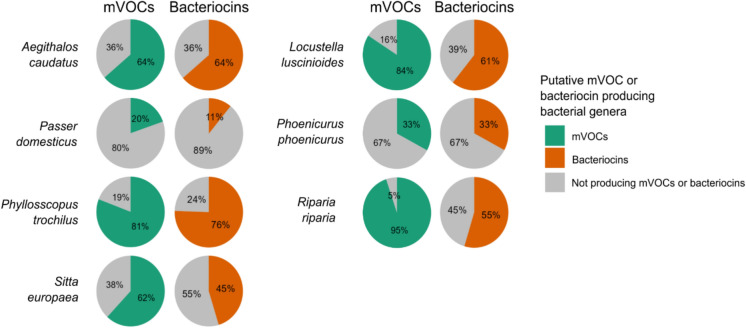 Figure 6
Figure 6- —https://doi.org/10.13039/501100003246Nederlandse Organisatie voor Wetenschappelijk Onderzoek
- —https://doi.org/10.13039/501100001824Grantová Agentura České Republiky
- —Institutional Research Support of the Czech Academy of Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsIdentification and Quantification in Food · Aquaculture disease management and microbiota · Marine animal studies overview
Introduction
Bacteria found in the preen glands of birds have been proposed to fulfil functional roles for the host, raising the question regarding the origin—host or bacterial—of the chemosignalling and antimicrobial functions associated with preen oil (Martín-Vivaldi et al. 2010; Whittaker et al. 2019). The preen gland, present in almost all bird species, secretes preen oil that birds spread on their plumage during preening to waterproof feathers (Jacob and Ziswiler 1982; Moreno-Rueda 2017). Besides feather maintenance and protection against mechanical wear, preen oil may also function to protect the avian host against feather degradation and infections caused by pathogenic bacteria and ectoparasites (Møller et al. 2010; Moreno-Rueda 2011, 2017; Videvall et al. 2021). There are many in vitro studies showing that preen oil inhibits the growth of plumage bacteria, including feather-degrading bacteria (Braun et al. 2018; Burger et al. 2004; Law-Brown 2001; Magallanes et al. 2016; Ruiz-Rodríguez et al. 2015; Shawkey et al. 2003; Soler et al. 2008). This antimicrobial activity may be mediated by a variety of preen oil antimicrobial compounds, including volatile organic compounds (VOCs) and proteins (Braun et al. 2018; Martín-Vivaldi et al. 2010). However, in vivo experimental studies found less consistent evidence for the antimicrobial activity of preen oil when comparing birds with and without access to their preen glands (Czirják et al. 2013; Giraudeau et al. 2013).
Preen oil might also be involved in the inter- and intraspecific communication of birds: the VOCs present in preen oil have the potential to produce odours and mediate interindividual communication (Grieves et al. 2022; Whittaker and Hagelin 2021). Recent insights have shown that birds have a functional olfactory system and that odours play a larger role in their communication than previously thought (Grieves et al. 2022; Whittaker and Hagelin 2021). For example, VOCs in preen oil have been found to serve as olfactory signals affecting mate choice, recognition, and predator and ectoparasite avoidance in birds (Grieves et al. 2022; Whittaker and Hagelin 2021).
Recent studies suggest that symbiotic bacteria in the preen gland may synthesize the chemicals, including VOCs, found in preen oil (Martín-Vivaldi et al. 2010; Whittaker et al. 2019). Bacteria that are known VOC producers were found in the preen glands of several species (Pearce et al. 2017; Rodríguez-Ruano et al. 2018; Whittaker et al. 2019), and bacteria isolated from Dark-eyed junco preen glands produced commonly found preen oil VOCs in vitro (Whittaker et al. 2019). The preen oil VOC profile and VOC abundance in preen oil were altered after the removal of the preen gland bacteriome using antibiotics in Eurasian hoopoes and Dark-eyed juncos, respectively (Martín-Vivaldi et al. 2010; Whittaker et al. 2019). This suggests that preen gland bacteria might be responsible for the production of at least part of the preen oil VOCs.
In addition to VOCs, the preen gland bacteria may produce other antimicrobials such as bacteriocins (ribosomally synthesized antimicrobial peptides) (Martín-Vivaldi et al. 2010; Soler et al. 2008). Bacterial strains isolated from preen glands of multiple bird species showed antimicrobial activity against indicator bacterial strains (including feather degrading bacteria) in vitro (Bodawatta et al. 2020; Martínez-Renau et al. 2022; Ruiz-Rodríguez et al. 2013, 2012; Soler et al. 2024), and in some strains this antimicrobial activity is mediated by the production of bacteriocins (Martín-Platero et al. 2006; Ruiz-Rodríguez et al. 2013, 2009).
The bacteriome of the preen gland thus likely produces VOCs and other antimicrobial compounds that help birds prevent feather degradation (VOCs and other antimicrobial compounds) and play a role in inter- and intraspecific communication (VOCs). Yet, detailed knowledge of the bacterial communities in the preen gland is limited to only a few bird species (Bodawatta et al. 2020; Grieves and Gloor2025; Grieves et al. 2021; Pearce et al. 2017; Rodríguez-Ruano et al. 2018; Videvall et al. 2021; Whittaker et al. 2019). Additionally, data on the presence of antimicrobial peptides in preen oil are limited to only two peptides in one bird species (House sparrow) (Carneiro et al. 2020), and no data exist on the complete proteomic profile of preen oil. Consequently, we have no insight into the variation in preen gland bacteriome composition, chemical (including VOCs) and proteomic profiles between birds, and the potential causes for this variation. Exploring preen oil bacterial, chemical and proteomic profiles in different bird species will be the foundation for the postulation of hypotheses on the causes of variation in preen gland bacteriome composition and preen oil chemicals (including VOCs) and antimicrobials in birds.
In this pilot study, we aim to i) explore the composition of the preen oil bacteriome, chemicals (including VOCs) and proteomic profiles, and ii) explore the association between the preen oil bacteriome and the presence of microbially derived preen oil VOCs (mVOCs) and antimicrobial peptides (represented by bacteriocins) in eight species of passerines.
Materials and methods
Study species and data collection
In total, we sampled 20 individuals (17 males:3 females) of 8 passerine species at 8 sites in the Czech Republic during March–May 2015–2016 (Table S1; Electronic Supplementary Material). The numbers of individuals and species used for preen oil analyses were as follows: 8 individuals of 8 species for the bacteriome profiling, 7 individuals of 7 species for the chemical/VOC profiling and 5 individuals of 4 species for the proteomic profiling (see Electronic Supplementary Table S1 for details). Since we have one individual per species for each type of analysis, we refer to variation between samples as variation between birds, as we cannot distinguish between inter- and intra-specific variation in our study. We captured all individuals using mist nests during their pre-breeding period (1–3 weeks prior to the typical peak breeding dates in the study region (Šta̕stný and Hudec 2011); see Electronic Supplementary Table S1) to minimise the confounding effects of seasonal differences in the chemical profile of preen oil (Potier et al. 2018; Soini et al. 2007; Tuttle et al. 2014). We removed each bird from the mist net, immediately placed it into a clean cloth bag and collected preen oil using a sterile 10 μl end-to-end glass capillary tube (Hirschmann Laborgeräte, Germany) wearing sterile nitrile gloves. We cut off the end of each capillary containing preen oil, placed it in a dry, sterile 1.8 ml screw-capped cryotube (Thermo Scientific – Nunc) with a rubber O-ring sealing to minimize the loss of VOCs, and stored it in a cooler at 5 °C for < 6 h. We then stored the cryotubes at − 20 °C until the preen oil bacterial, volatile and proteomic profiling. Although all sampling and storage procedures were carried out with maximal caution, it is worth noting that losses of some very volatile organic compounds (VVOCs) occurred. This may result in incomplete VOC profiles presented in this study, as these compounds may be missing. However, it is impossible to fully prevent this during the sampling of preen oil from free-living birds.
Preen oil bacteriome analysis
We extracted DNA from preen oil using the RTP® Bacteria DNA Mini kit (STRATEC Molecular GmbH, Berlin, Germany) following the isolation kit’s Protocol 5 (Isolation of microbial DNA from tissue biopsies). Negative DNA extraction was done along with the samples. For each sample (including the negative control), we diluted the isolated DNA tenfold in nuclease-free H_2_O (Qiagen, Hilden, Germany), from which we used 2 µL as a template in the PCR reaction. Next, we amplified the bacterial variable V4–V5 region of 16S rRNA gene using the specific primer pair BactB-F (GGATTAGATACCCTGGTAGT) and BactB-R (CACGACACGAGCTGACG) (Fliegerova et al. 2014), and performed the PCR reaction using EliZymeTM HS FAST MIX Red Master Mix (Elisabeth Pharmacon, Brno, Czech Republic). Thermal cycling conditions included an initial denaturation for 5 min at 95 °C, followed by 25 cycles consisting of 30 s at 95 °C, 30 s at 57 °C, 30 s at 72 °C, and a final elongation step at 72 °C for 5 min. We checked the length and quality of the amplicons by agarose gel electrophoresis (1.5%) and purified the PCR products using the Monarch® PCR & DNA Cleanup Kit (New England BioLabs, Ipswich, MA, USA). The negative DNA control did not amplify successfully and was therefore excluded from the library preparation.
We performed the library preparation using the NEBNext Fast DNA Library Prep Set for Ion Torrent (New England BioLabs, Ipswich, MA, USA) and the Ion Xpress Barcode Adapters 1–96 Kit (Thermo Fisher Scientific, Waltham, MA, USA). We analysed the length of the target amplicons of DNA libraries using the 2100 Bioanalyzer Instrument (Agilent Technologies, Santa Clara, CA, USA) and pooled the amplicons in equimolar ratios based on concentration determined using a KAPA Library Quantification Kit (KAPA Biosystems, Roche, Pleasanton, CA, USA). The template amplification and enrichment were performed by emulsion PCR in the Ion OneTouch™ 2 instrument using the Ion PGMTM HiQTM View OT2 Kit-400 (Thermo Fisher Scientific, Waltham, MA, USA). The enriched template was sequenced with the Personal Genome Machine (PGM™) System (Thermo Fisher Scientific,Waltham, MA, USA) using the Ion PGM™ Hi-Q™ View Sequencing solutions kit and the Ion 316™ Chip v2 BC according to the manufacturer’s protocols.
We first detected and trimmed gene-specific primer sequences with a skewer (Jiang et al. 2014). Using the R package dada2 (Callahan et al. 2016), we trimmed all reads to the same length (260 bp), eliminated low-quality reads (expected error rate per paired-end read > 2), and denoised quality-filtered reads using the parameters recommended for Ion Torrent data by the dada2 developers (i.e., HOMOPOLYMER_GAP_PENALTY = − 1 and BAND_SIZE = 32). We then created an abundance matrix that included the number of reads for each 16S rRNA amplicon sequencing variant (ASVs) in each sample. Using Uchime (Edgar et al. 2011) in conjunction with the gold.fna reference database (available at: https://drive5.com/uchime/gold.fa), we detected and eliminated chimeric ASVs. Subsequently, we assigned the taxonomy for non-chimeric ASVs with 50% posterior confidence by the RDP classifier (Wang et al. 2007) using the Silva database v.138 (Quast et al. 2012) as a reference.
We performed further filtering in QIIME2 (v 2022.11) (Bolyen et al. 2019), where we filtered ASVs assigned to Archaea, Eukaryota, Mitochondria, and Chloroplasts, as well as the ASV assigned to Delftia sp., as this abundant ASV, present in all samples, is often found to be a contaminant from DNA extraction kits (Salter et al. 2014). Next, we rarefied the ASV table at a sequencing depth of 2765 reads, based on alpha rarefaction curves (Fig. S1; Electronic Supplementary Material) that saturated at 2000 reads for both the number of observed ASVs and Shannon diversity and the read count of the first sample with read counts > 2000 (2769 reads). Rarefying removed 1 sample from the dataset (Acrocephalus arundinaceus), leaving 7 samples from 7 passerine species*.*
Preen oil untargeted chemical/VOC profiling
Preen oil samples were processed in a service laboratory (Mass Spectrometry Laboratory of the University of Chemistry and Technology Prague) using stir bar sorptive extraction (SBSE) coupled with gas chromatography-mass spectrometry (GC–MS), with slightly modified conditions, described in Soini et al. 2013 (Soini et al. 2013). Briefly, the glass microcapillary tube containing the preen oil sample was placed into a 20 mL capped glass vial with 2 mL of high-purity water and a Twister™ stir bar (10 mm, 0.5 mm film thickness, 24 μL polydimethylsiloxane, PDMS, volume). Adding 100 mg of ammonium sulphate was omitted for most of the samples because a pre-experimental trial with one preen oil sample originating from Sand Martin uncovered a problem with this method. Despite rinsing the stir bar with distilled water, ammonium sulphate remained on the surface of the stir bar. During subsequent thermal desorption, ammonium sulphate decomposed into NH_3_ and SO_2_, and especially the presence of NH_3_ caused significant “bleeding” of the CG column (mostly visible on one sand martin chromatogram), leading to rapid degradation of the stationary phase of the GC column and Twister™ stir bar destruction. As an internal standard, 8 ng of 7-tridecanone (Sigma Aldrich, Germany) was added to 5 μL of methanol in each vial. The sample was stirred at 800 rpm on an HSC 7 VELP magnetic stirrer (VELP Scientifica, Italy) for 60 min. Before extraction, all glassware was washed with acetone and dried at 80 °C. After extraction, the stir bars were gently rinsed with a small amount of distilled water, dried on paper tissue and placed in the Thermal Desorption Autosampler tubes (Gerstel GmbH, Germany) for subsequent gas-chromatography–mass spectrometric (GC–MS) analysis. Together with the samples, a standard mixture of n-alkanes (49,452-U Supelco, Sigma Aldrich, Germany) was analysed under the identical GC–MS conditions specified below to determine the retention behaviour of the individual analytes contained in the preen oil samples on the GC column.
For GC–MS profiling, a Trace GC Ultra (Thermo Scientific) gas chromatograph connected to an ISQ mass spectrometer (Thermo Scientific) with a Thermal Desorption Autosampler with a Cooled Injection System (TDSA–CIS 4) was used. The separation capillary was DB-5MS Agilent (30 m × 0.25 mm, i.d., 0.25 m film thickness). Samples were thermally desorbed in a TDSA automated system, followed by a splitless injection into the column with a CIS-4 cooled injection assembly. The temperature separation program on the GC column was as follows: 50 °C (4 min), then increasing to 290 °C at the rate of 10 °C/min 3 °C/min (hold time: 30 min).
To identify the organic compounds in the preen oil samples, the measured mass spectra of individual chromatographic peaks (controlled also against n-alkanes standard RIs) were compared with the NIST 14 mass spectra database (https://chemdata.nist.gov/) with match factor cut-off scores ranging between 900 and 950 (i.e. excellent match). Blank measurements of the Twister and all backgrounds were subtracted from the chromatograms before further processing. We assigned our identified organic compounds as (semi-)volatile organic compounds ((S)VOC) if the compound vapour pressure at a temperature of 293.15 K (20 °C) is between 10^–8^ and 10^–2^ kPa (sVOC) or more than 10^–2^ kPa (VOC) (Directive 2010/75/EU; Liu 2022). All identified compounds were assigned as (S)VOCs, except for 3 compounds for which no data on vapour pressure at 293.15 K was available (Table S3; Electronic Supplementary Material). The identification of compounds based on comparing measured mass spectra with the NIST 14 mass spectra database was only possible for compounds with retention times (RTs) up to 25 min (Fig. S2; Electronic Supplementary material). The organic compounds with RTs > 25 min can only be identified as a heterogeneous mixture of fatty acid alkyl esters. For accurate identification of these compounds, it is necessary to measure the standards under the same chromatographic conditions and then compare the retention times and mass spectra of these standards with the measured data from the preen oil analyses. Since our study focused only on the identification of preen oil (S)VOCs, and since the compounds with a retention time (RT) > 25 min (of non-volatile nature) could not be identified with certainty, the estimation of relative abundances of detected (S)VOCs in the complete profiles would be highly biased. Therefore, we limited our analysis to the presence/absence data of the (S)VOCs detected in the preen oil of the species studied.
Preen oil proteomic profiling
We performed preen oil proteomic profiling using LC–MS/MS. Specifically, the nano-HPLC apparatus used for protein digest analysis was a Proxeon Easy-nLC (Proxeon, Odense, Denmark) coupled to an ultrahigh resolution MaXis Q-TOF (quadrupole—time of flight) mass spectrometer (Bruker Daltonics, Bremen, Germany) by nanoelectrosprayer. We used the software packages HyStar 3.2 and micrOTOF-control 3.0 to control the nLC-MS/MS instruments and collected and manipulated the data with the software packages ProteinScape 3.0 and DataAnalysis 4.0 (Bruker Daltonics). We used the following chemicals during the analysis; trypsin (TPCK treated, from bovine pancreas, 13,500 units per mg), ammonium bicarbonate and acetonitrile (HPLC–MS grade) obtained from Sigma (St. Louis, MO, USA) and prepared all solutions in MilliQ water (Millipore, Bedford, MA, USA).
We inserted the capillaries with preen oil samples into glass vials with 30 μl of trypsin buffer (0.2 mg/ ml trypsin in 100 mmol/L ammonium bicarbonate pH 7.8), washed the contents of the capillaries out five times with this buffer (centrifugated at 6000 g) and treated the contents with trypsin buffer for 18 h at 37 °C. After trypsin cleavage, we extracted peptides from the samples with StageTips using Empore C18 Extraction disks (Supelco; Bellefonte, PA, USA) according to the published protocol (Rappsilber et al. 2007).
We injected five microliters of the peptide mixture into an NS-AC-12dp3-C18 Biosphere C18 column (particle size: 3 µm, pore size: 12 nm, length: 200 mm, inner diameter: 75 µm) with an NS-MP-10 Biosphere C18 precolumn (trap-column) (particle size: 5 µm, pore size: 12 nm, length: 20 mm, inner diameter: 100 µm), both manufactured by NanoSeparations (Nieuwkoop, The Netherlands). The separation of peptides was achieved via a linear gradient between mobile phase A (water) and B (acetonitrile), both containing 0.1% (v/v) formic acid. We started separation by running the system with 5% mobile phase B, followed by a gradient elution to 7% B at 5 min and 30% B at 180 min. The next step was a gradient elution to 50% B in 10 min and then a gradient to 100% B in 10 min. Finally, we eluted the column with 100% B for 20 min. We achieved equilibration between the runs by washing the column with 5% mobile phase B for 10 min. The column was held at ambient temperature (25 °C), and the flow rate was 0.20 µL/min. We used online nano-electrospray ionization (easy nano-ESI) in positive mode. We set the ESI voltage to + 4.5 kV, the scan time to 3 Hz, the drying gas (N_2_) to 4 L/min, the drying gas temperature to 180 °C, and the nebulizer pressure to 100 kPa. Measurements were performed by scanning from 50 to 2200 m/z. The reference ion (internal mass lock) was a monocharged ion of C_24_H_19_F_36_N_3_O_6_P_3_ (m/z 1221.9906). We averaged mass spectra corresponding to each signal from the total ion current chromatogram for an accurate molecular mass determination. We conducted all LC–MS and LC–MS/MS analyses in duplicate.
We processed the data using ProteinScape software v. 3.0.0.446 (Bruker Daltonics, Bremen, Germany). We identified proteins by correlating tandem mass spectra to the extracted database for Aves from the NCBI database (downloaded on 30th January 2017—2,198,542 sequences; 1,077,083,019 residues), using the MASCOT searching engine v. 2.3.0 (http://www.matrixscience.com). We used the whole UniProt database for control searching, choosing trypsin as the enzyme parameter, allowing three missed cleavages, and using an initial peptide mass tolerance of ± 10.0 ppm for MS and ± 0.05 Da for MS/MS analysis. We set the following modifications to be variable: proline and lysine were allowed to be hydroxylated and methionine oxidated, whereas asparagine and glutamine were deamidated. We set the monoisotopic peptide charge to 1 + , 2 + and 3 + , and selected the Peptide Decoy option during the data search process to remove false-positive results. Only significant hits were accepted (MASCOT score ≥ 80 for proteins and MASCOT score ≥ 20 for peptides, http://www.matrixscience.com). All identified proteins and their sequences were manually validated using the peptide search tool of the UniProt database (release 2023_03). We listed identified proteins with the highest UniProt BLAST similarities, along with accompanying information (i.e., primary NCBI, UniProtKB and UniParc accession numbers, source organism, evidence for protein existence, annotation score and its status (unreviewed—TrEMBL or reviewed-UniProtKB/Swiss-Prot) in summarizing tables (Table S2, Electronic Supplementary Material).
Statistics and data analysis
We calculated Jaccard distances with rarefied data using the QIIME2 pipeline. The QIIME2 output was imported into R statistical software v 4.2.2 (qiime2R R package v 0.99.6) for further analysis and visualisation. We generated taxa barplots of the relative abundance at the phylum, class and genus level, as well as a heatmap of the relative abundance at the genus level, using the non-rarefied feature table and taxonomy assignment (ggplot2 R package v 3.4.1 (Wickham 2016)). We used the rarefied feature table to create a shared ASV Upset plot (ComplexUpset R package v 1.3.3 (Krassowski 2021)).
To assess the potential functional characteristics of the preen oil proteomic profiles, we used the complete list of UniprotKB and UniParc accession numbers for all proteins identified in the preen oil of specific passerine species as a source to visualise the Gene Ontology (GO) of the identified preen oil proteins using the UniprotR R package (Soudy et al. 2020).
We combined the preen oil bacteriome data with chemical profiling data to determine whether the bacterial genera found in preen oil have the potential to produce the VOCs detected in preen oil. First, we entered all compounds found in preen oil into the microbial VOC (mVOC) 4.0 Database (Kemmler et al. 2025) and recorded which bacterial genera were found to produce these VOCs. Then, based on data from the mVOC database, for each bacterial genus detected in preen oil from a particular species, we determined whether that bacterial genus could produce VOCs found in preen oil. With this data, we created pie charts (ggplot2 v 3.4.1 (Wickham 2016)) of the relative abundance of the bacterial genera found in each bird species, including the information on whether the bacterial genus has the potential to produce preen oil mVOCs. To further assess the relationship between preen oil bacterial communities and VOC profiles, we assessed differences in chemical profiles between species using the Jaccard dissimilarity index based on presence/absence data (vegan R package v 2.6–4 (Oksanen et al. 2022)). We then performed a Procrustean analysis on the PCoA of chemical data and the PCoA of bacteriome data (Jaccard). To determine the significance of the association between preen oil bacterial communities and chemical profiles, we used a permutation procedure for Procrustean rotation (PROTEST) with 999 permutations (vegan v 2.6-4 (Oksanen et al. 2022)) (Jackson 1995; Peres-Neto and Jackson 2001). We created shared chemical compound Upset plots using the ComplexUpset package v 1.3.3 (Krassowski 2021).
To determine the potential of the preen oil bacteriome to produce bacteriocins, we searched all bacterial genera found in preen oil in the BACTIBASE database (Hammami et al. 2010). Each genus that was found to produce at least one bacteriocin, was indicated as putative bacteriocin producing. Next, we created pie charts with relative abundances of bacteriocin-producing bacterial genera (R package ggplot2 R package v 3.4.1 (Wickham 2016)).
Results
Preen oil bacteriome
Firmicutes (11–76%) and Actinobacteriota (11–45%) were the most abundant phyla in the preen oil bacteriome composition in most of the passerine species (Fig. 1A). Proteobacteria were found in preen glands of 5 of the 7 species, and were not detected in the Savi’s warbler (Locustella luscinioides) and Sand martin (Riparia riparia). The preen oil bacteriome of the House sparrow had a relatively high abundance of the phylum Proteobacteria (75%). The composition of the House sparrow’s bacteriome was also unique at the class and genus level (Fig. 1B, C). For example, the genus Enhydrobacter (phylum Proteobacteria) was exclusively found in the House sparrow. On the other hand, Streptococcus (phylum Firmicutes) was found in all species except the House sparrow. Lactococcus (phylum Firmicutes) was highly abundant in the Willow warbler (Phylloscopus trochilus) (Fig. 1C).Fig. 1. Preen oil bacteriome community composition in different passerine species expressed as variability in relative abundances of amplicon sequence variants (ASVs) (%) of A Bacterial phyla, B Bacterial classes, and C Bacterial genera
Only one ASV (ASV28, Genus: Cutibacterium) was shared between all species (Fig. 2), and generally, most ASVs were unique to individuals. The House sparrow had the highest number of unique ASVs compared to the other birds.Fig. 2. Upset plot showing the number of shared and unique amplicon sequence variants (ASVs) in the preen oil bacteriome of studied passerine species, where each ASV is coloured by bacterial class. The total number of ASVs per individual is indicated in brackets following the species name
Preen oil chemical/VOC profiles
The number of distinct, identified compounds found in each species ranged from 29 in the Savi’s warbler to 5 in the European nuthatch (Sitta europaea) (Fig. 3, Fig. S3 and Table S3; Electronic Supplementary Material). In total, 38 distinct compounds were found in the preen oil of 7 species (Fig. 3, Fig. S3 and Table S3; Electronic Supplementary Material). All preen oil compounds for which data on vapour pressure is available (89% of compounds) were identified as (S)VOCs, and 73% of all preen oil compounds were found in the mVOC 4.0 database indicating that most of the identified preen oil compounds have a high degree of volatility. The major compound classes detected were alcohols, aldehydes, ketones, and carboxylic acids; the alcohols and ketones were exclusively found in the Great reed warbler and Savi’s warbler (Fig. 3, Fig. S3 and Table S3; Electronic Supplementary Material).Fig. 3. Barplots of (A) absolute and (B) relative numbers of identified organic compounds that were mostly identified as (S)VOCs, coloured by their higher chemical classification, in preen oil of studied passerine species
Four compounds were shared between all 7 birds (Fig. 4). The Savi’s warbler and Great reed warbler shared 11 compounds, and the Savi’s warbler had the highest number of compounds unique to that individual (7). The difference in preen oil VOC profiles between birds is also apparent from the chromatograms (Fig. S2; Electronic Supplementary Material).Fig. 4. Upset plot showing the number of distinct shared and unique chemical compounds in preen oil of studied passerine species, where each compound is coloured by their higher chemical classification. The total number of compounds per individual is indicated in brackets following the species name
Preen oil proteomic profiles
The Long-tailed tit (Aegithalos caudatus) and Willow warbler showed the highest number of proteins identified (41 and 30 respectively, see Table S2; Electronic Supplementary Material), while the lowest number of proteins were identified for the Sand martin individuals (14 and 16; Table S2; Electronic Supplementary Material). None of the proteins identified in the preen oils were related to proteins produced by microorganisms. Conversely, all preen oil proteins had the highest similarity to those derived from birds (Table S2; Electronic Supplementary Material).
Lysozyme was the only antimicrobial protein that we detected in preen oil, and it was only present in the Long-tailed tit (Table S2; Electronic Supplementary Material). Biological processes of all proteins identified in passerine preen oil inferred from GO analysis differed among birds (Fig. S4; Electronic Supplementary Material) and were dominated with proteolysis, response to oxidative stress, hydrogen peroxide catabolic process and fatty acid beta-oxidation (Fig. 5). Most of the preen oil proteins were identified as cellular components of keratin filament, intermediate filament and peroxisome and their molecular function was dominated by structural molecular activity, metal ions and heme binding followed by oxidoreductase and catalase activity (Fig. 5).Fig. 5. Gene ontology (GO) inferred functional properties (red = biological processes, green = molecular functions, blue = cellular component) of proteins detected in preen oil of four studied passerine species
Preen oil bacteriome and associated preen oil mVOCs and bacteriocin production
The Sand martin and Savi’s warbler bacteriome had the highest potential for producing preen oil mVOCs with 95% and 84% relative abundance of putative mVOC-producing genera, respectively. On the other hand, in the House sparrow bacteriome only 20% of the genera present had the potential to produce preen oil mVOCs (Fig. 6). The Procrustes analysis did not find a significant concordance between preen oil bacteriome community structure and preen oil chemical profile (protest: m_12_ = 0.73, P = 0.53), indicating that there may be no or a weak association between preen oil bacteriome composition and preen oil chemical profiles. However, considering the lack of replicates and the fact that the bacteriome and chemical profiles were analysed from different individuals, these results should be taken with caution (Fig. S5; Electronic Supplementary Material).Fig. 6. Relative abundances of putative preen oil bacteriocin-producing bacterial genera (orange) as inferred from the BACTIBASE database and microbial VOC-producing bacterial genera (green) capable of producing preen oil mVOCs as inferred from the mVOC database and chemical profiles of studied passerines. Grey indicates the relative abundance of bacterial genera with no potential to produce bacteriocins or mVOCs
The House sparrow had the lowest relative abundance of putative bacteriocin-producing bacteria (11%), while in the preen oil of the Willow warbler, 76% of the bacterial genera are putative bacteriocin producers. (Fig. 6).
Discussion
Our exploratory study revealed substantial variation in the preen oil bacteriome of passerine birds, where Firmicutes and Actinobacteriota were the most dominant phyla. Likewise, the preen oil chemical compounds varied in profile and diversity among birds, just as the proteomic profile. We identified a total of 39 distinct chemical compounds (mostly assigned as (S)VOCs) consisting of alcohols, aldehydes, ketones, and carboxylic acids, as well as 108 unique proteins primarily associated with proteolysis, response to oxidative stress, hydrogen peroxide catabolic process and fatty acid beta-oxidation. Interestingly, we found that the preen gland bacteriome can potentially produce VOCs and bacteriocins found in passerine preen oil. To further understand how much of the variation in these traits can be attributed to species and individuals, or to ecological factors and life-history traits of birds, requires follow up studies.
Variability of preen oil bacteriome
Here, the preen oil bacteriome varied among 8 passerine birds from 8 species (one individual per species), raising the question how much of this variation is attributable to the levels of species and individual (Bodawatta et al. 2020; Videvall et al. 2021). There is strong evidence for between-species variability in bacteriomes of preen glands, feathers and skin, in both free-living and captive birds (Engel et al. 2018; Grieves and Gloor 2025; Javůrková et al. 2019; Roggenbuck et al. 2014), but see (van Veelen et al. 2017). The composition of the preen oil bacteriome at the phylum level is consistent with those found in other studies, with Firmicutes and Actinobacteriota being the most dominant phyla (Bodawatta et al. 2020; Grieves et al. 2021; Pearce et al. 2017; Rodríguez-Ruano et al. 2018; Videvall et al. 2021; Whittaker et al. 2019). The phylum Proteobacteria, which we found to be abundant in the preen oil of the House sparrow, was also reported to be abundant in another study on House sparrows, as well as in Dark-eyed juncos and Leach’s storm petrels (Pearce et al. 2017; Videvall et al. 2021; Whittaker et al. 2019). Similarly to our results, the class Gammaproteobacteria and the genus Enhydrobacter were found to be highly abundant in the preen gland bacteriome of House sparrows (Videvall et al. 2021). Because the House sparrow is the only urban bird in our study, we hypothesize that the different taxa found in the House sparrow may be linked to their urban environment.
Preen oil chemical/VOC profile
The number of distinct preen oil chemical compounds ranged from 5 to 29 and consisted of alcohols, aldehydes, ketones, carboxylic acids, monoterpenes and monoterpene derivatives, unsaturated fatty acids, esters, and a few other compounds. Most of these classes of compounds were assigned as compounds with high degree of volatility (i.e. as (S)VOCs) and are frequently found in previous studies focused on passerines (Alves Soares et al. 2024). The specific aldehydes, monounsaturated fatty acids, esters, the dialkyl ketone (heptan-2-one), the phenol, and some of the monoterpenes and monoterpene derivatives that we detected are either rarely observed or have never been detected before in preen oil of passerines (Díez-Fernández et al. 2021; Shaw et al. 2011; Soini et al. 2007, 2013; Tuttle et al. 2014). However, it is important to note that due to variations in VOC sampling, storage conditions, extraction and identification methods, direct comparisons with VOC data from other studies are difficult (Alves Soares et al. 2024). We found variation in chemical/VOC profiles between the studied birds, which may reflect interspecific differences. However, due to the lack of replicates, we can not conclude that these differences are indeed reflective of species effects. Comparative studies on passerines showed interspecific differences in VOC profiles, both in the number of distinct VOCs and in classes of compounds (Soini et al. 2013; Zhang et al. 2009). Interestingly, alcohols and methyl ketones, detected in 16 out of 16 and 14 out of 16 passerine species, respectively, studied by Soini et al. (2013) (Soini et al. 2013), were only present in two species (Great reed warbler and Savi’s warbler) in our study.
Antibacterial and antifungal activity is known for many of the preen oil VOCs detected in this study, including all aldehydes (Bisignano et al. 2001; Fernando et al. 2005; Inouye et al. 2001; Lammers et al. 2021; Tao et al. 2014) and some of the alcohols (Gehrke et al. 2013), ketones (Popova et al. 2014), carboxylic acids (Chadeganipour and Haims 2001; Ganesan et al. 2022; Huang et al. 2011; Lee et al. 2021; Sahin et al. 2006; Seidel and Taylor 2004) and a few other compounds (geranyl acetone (Kubo et al. 1993), linalool (Inouye et al. 2001; Tao et al. 2014), oleic acid (Muthamil et al. 2020; Seidel and Taylor 2004), tributyl phosphate (Ferreira et al. 2010), and 2,4-ditertbutylphenol (Chawawisit et al. 2015; Zhao et al. 2020)). Several of these compounds have an antimicrobial activity against keratinolytic bacterial and fungal species (Bisignano et al. 2001; Chadeganipour and Haims 2001; Ganesan et al. 2022; Gehrke et al. 2013; Sahin et al. 2006), suggesting a potentially protective role of these preen oil compounds against feather-degrading bacteria.
Additionally, preen oil VOCs may play a role in detection by ectoparasites. Host-seeking parasites such as mosquitoes can use VOCs to detect a suitable host and the host location (Spanoudis et al. 2022; Tomás et al. 2020). On the other hand, certain VOCs can also repel parasites (Tomás et al. 2020). Several of the VOCs found in this study are known to repel or attract ornithophilic mosquitoes, including geranyl acetone, carboxylic acids, pentadecan-2-one, and aldehydes (Dormont et al. 2021; Logan et al. 2010; Mweresa et al. 2016; Puri et al. 2014; Spanoudis et al. 2022). Nonanoic acid, linalool and geranyl acetone repel Culicoides biting midges (González et al. 2014; Logan et al. 2009). Aldehydes, linalool, and dodecanoic acid are highly repellent or lethal against ticks and feather lice (Douglas 2008, 2013; Weldon et al. 2011; Zhu et al. 2018). Lastly, oleic acid, detected in nearly all bird species in our study, strongly repels arthropods (Yao et al. 2009). However, it should be noted that for many of these compounds, the ectoparasite repellency is strongly dose-dependent (González et al. 2014; Logan et al. 2009, 2010; Puri et al. 2014). Therefore, the lack of data on the concentrations of these VOCs in preen oil makes it difficult to determine whether the presence of these compounds in preen oil indeed plays a protective role against ectoparasites.
Preen oil proteomic profile, and host or bacterial origins of antimicrobial proteins
For the first time, we characterized the complete proteomic profile of preen oil in passerines, where the number of proteins detected ranged from 14 to 40, and the Gene Ontology (GO) inferred biological processes associated with the proteins were dominated with proteolysis, response to oxidative stress, hydrogen peroxide catabolic process and fatty acid beta-oxidation. Fatty acid synthesis is an important function of the preen gland (Moreno-Rueda 2017). Proteins are likely to play a role in preen oil lipid synthesis (Biester et al. 2012), and proteolysis, the breakdown of proteins, plays a role in the quality control of proteins in order to regulate cellular processes (Buchberger et al. 2010). We found predominantly proteins associated with proteolysis and energy metabolism (protection against oxidative stress and fatty acid beta-oxidation) in preen oil. While only one detected protein in one species (Willow warbler) was directly related to lipid metabolism, it is possible that the proteins related to proteolysis and energy metabolism play an indirect role in fatty acid synthesis. Further experimental and physiological studies are however needed to fully understand the role of preen oil proteins in birds.
Proteins have been hypothesized to contribute to the antimicrobial activity of preen oil (Braun et al. 2018; Carneiro et al. 2020), and indeed, an earlier study detected lysozyme and immunoglobulin Y in preen oil of House sparrows (Carneiro et al. 2020). In this study, we did not find any antimicrobial proteins (e.g. bacteriocins) in our study species, except for lysozyme in the preen oil of the Long-tailed tit. This lysozyme was derived from birds and, therefore, likely synthesised by the host, not by the preen gland bacteria. Lysozymes are widespread antimicrobial enzymes that are a crucial part of the innate immune system of vertebrates (Callewaert and Michiels 2010). When present in preen oil, lysozymes may be involved in the defence of the preen gland against bacterial colonization or protect the feathers against feather-degrading bacteria (Carneiro et al. 2020).
In our study, we did not detect bacteriocins (bacterially produced antimicrobial peptides) in preen oil in any bird species, while we did detect putative bacteriocin-producing bacterial genera in all preen oil samples. It is important to note that putative bacteriocin-producing potential was inferred from presence of bacterial genera in the BACTIBASE database, not directly measured. Preen oil is hypothesized to contain bacteriocins to protect the feathers against feather-degrading bacteria, but thus far, only the Eurasian hoopoe was found to host bacteriocin-producing bacteria in their preen gland (Martín-Platero et al. 2006; Ruiz-Rodríguez et al. 2013). Potential explanations for the absence of bacteriocins in the proteomic profiles of the studied passerines are that bacteriocins are only produced under certain conditions (Braun et al. 2018; Heilbronner et al. 2021; Meade et al. 2020) and have low stability in vivo due to their susceptibility to proteolytic degradation (Meade et al. 2020). Therefore, bacteriocin detection may require other analytical methods, such as capillary electrophoresis–mass spectrometry (CE–MS) or nano LC/MS–MS preceded by a purification and filtration processes (Nandakumar and Talapatra 2014; Zou et al. 2018). The variation that we found in putative bacteriocin-producing bacteria is in line with earlier hypotheses that birds that face a higher pressure of feather-degrading bacteria on their plumage have acquired a preen oil bacteriome with higher abundances of bacteriocin-producing bacteria to protect the plumage against feather-degrading bacteria (Javůrková et al. 2019). However, in the absence of replicates, it remains uncertain whether this variation in putative bacteriocin-producing bacteria between birds is attributable to species-level differences or inter-individual variation.
Association between preen oil bacteriome and VOCs
We have shown that the preen oil contains putative preen oil mVOC-producing bacterial genera, and that the relative abundances of these putative preen oil mVOC-producing bacterial genera varied between birds. This indicates that the preen oil bacteriome has the ability to synthesise some of the VOCs found in preen oil and that the contribution or role of the bacteriome in the production of VOCs may vary between birds. Indeed, a few previous studies have found strong indications that preen gland bacteria may produce VOCs (Martín-Vivaldi et al. 2010; Whittaker et al. 2019). However, it is important to note that this potential to produce preen oil mVOCs was inferred from a database, not measured directly. Although we did not find a statistically significant association between the preen oil bacteriome profiles and chemical profiles, this lack of association can occur if only a part of the preen oil chemical compounds is synthesised by the bacteriome and the rest by the host, which is highly probable. In addition, our sample sizes were limited to one individual per species and our chemical profile and bacteriome samples were derived from different individuals of given species, making it more difficult to directly explore the link between the bacteriome and chemical profiles. Therefore, our results on the abundances of putative preen oil mVOC producing bacterial genera and on the association between the preen oil bacteriome and chemical profiles should be interpreted with caution. Our results are however consistent with studies on dark-eyed juncos, where no covariation was detected between microbial profiles and volatile odour profiles in a field study (Whittaker et al. 2016) but found a role for preen gland bacteria in VOC production in an experimental study (Whittaker et al. 2019).
We found the highest diversity of preen oil VOC profiles and the highest relative abundances of putative preen oil mVOC-producing bacteria in the riparian bird species. Some of the compounds found exclusively in the riparian species have known antimicrobial activity, antifungal activity and mosquito and biting midge repellency (Chawawisit et al. 2015; Gehrke et al. 2013; González et al. 2014; Kubo et al. 1993; Logan et al. 2009, 2010; Popova et al. 2014; Zhao et al. 2020). The high humidity in riparian habitats may increase the prevalence of (keratinolytic) bacteria on feathers and promote the abundance of ectoparasites (Burtt and Ichida 2004; Radrova et al. 2013; Werner et al. 2020), therefore we hypothesize that the VOC profiles of the riparian species may reflect adaptive responses to habitat humidity. However, because of the limited number of species and absence of replicates, this hypothesis remains tentative and future studies with larger datasets are required to test this hypothesis.
To conclude, our findings provide preliminary insights into the preen oil bacteriome, chemical and proteomic profiles and highlight their possible associations. Further comparative and combined experimental research (e.g. adopting preen oil bacteria transplantation) with larger datasets, including more individuals per species and a broader spectrum of bird species, may help to identify the reasons for bacteriome, chemical and proteome variation between birds, which may be driven by a combination of host species traits, such as phylogeny, habitat type and other environmental factors. Moreover, the gas-phase headspace of preen oil obtained using the SPME (solid-phase micro-extraction) method is necessary for future studies, enabling more sensitive and complex profiling of preen oil VOCs.
Supplementary Information
Below is the link to the electronic supplementary material.Supplementary file1 (XLSX 70 KB)Supplementary file2 (DOCX 2010 KB)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Directive 2010/75/EU (2010) Directive 2010/75/EU of the European Parliament and of the Council of 24 November 2010 on industrial emissions (integrated pollution prevention and control). https://eur-lex.europa.eu/eli/dir/2010/75/oj/eng
- 2Law-Brown J (2001) Chemical defence in the red-billed wood hoopoe: phoeniculus purpureus. University of Cape Town