The Developmental Environment Mediates Adult Seminal Proteome Allocation in Male Drosophila melanogaster
Rebecca von Hellfeld, Rebecca Konietzny, Philip D. Charles, Roman Fischer, Benedikt M. Kessler, Stuart Wigby, Irem Sepil, Juliano Morimoto

TL;DR
Early life conditions in fruit flies affect adult male reproductive protein allocation, with small males transferring more specific proteins per mating.
Contribution
This study reveals how larval crowding influences seminal proteome allocation in male Drosophila, including male-specific protein transfer patterns.
Findings
Large males produced more seminal fluid proteins overall.
Small males transferred higher amounts of specific proteins per mating.
Ten specific proteins were transferred in significantly higher amounts by small males.
Abstract
Early life conditions can have long‐lasting effects on fitness. In insects, larval crowding increases intraspecific competition, shaping adult phenotypes and influencing male reproductive success in pre‐ and post‐mating competition. Although some larval crowding effects on seminal fluid protein (Sfp) allocation are known, studies often focus on a small subset of Sfps and overlook male–female interactions. A comprehensive understanding of how male and female larval environments interact to influence seminal proteome composition and transfer is still lacking. Here, we manipulated Drosophila melanogaster larval crowding (low vs. high) to generate large and small adults and mated individuals in a fully factorial design. We then measured Sfp production, composition and transfer. Large males produced relatively higher quantities of Sfps. However, small males transferred greater quantities…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
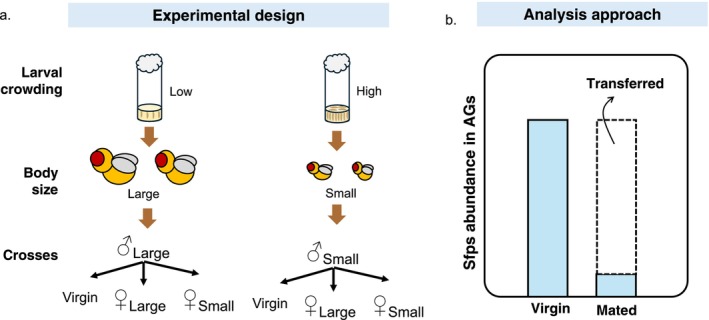 FIGURE 1
FIGURE 1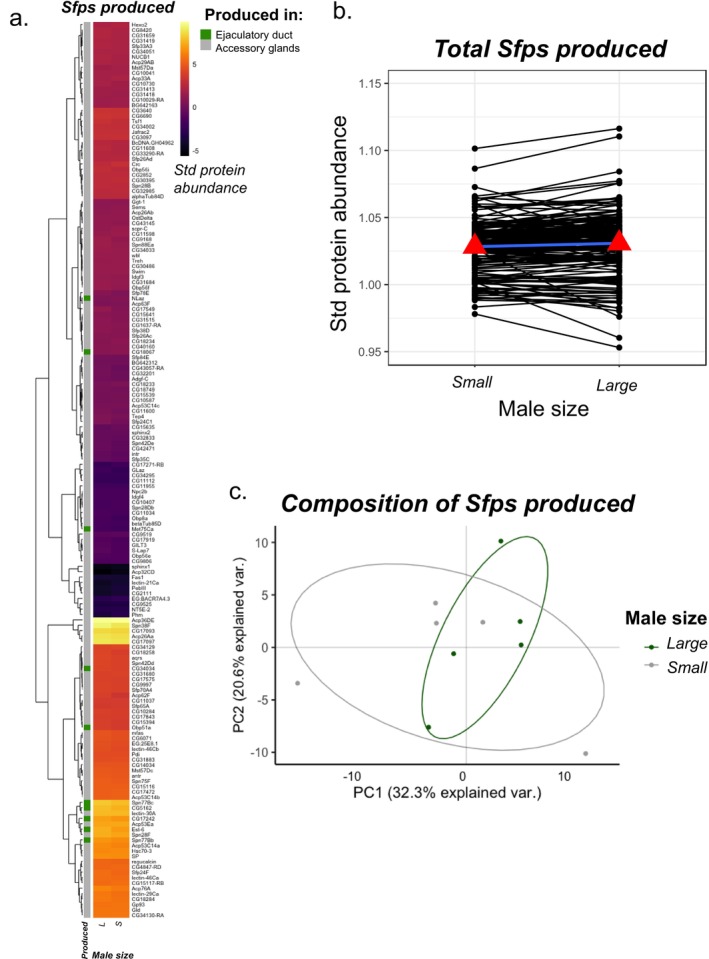 FIGURE 2
FIGURE 2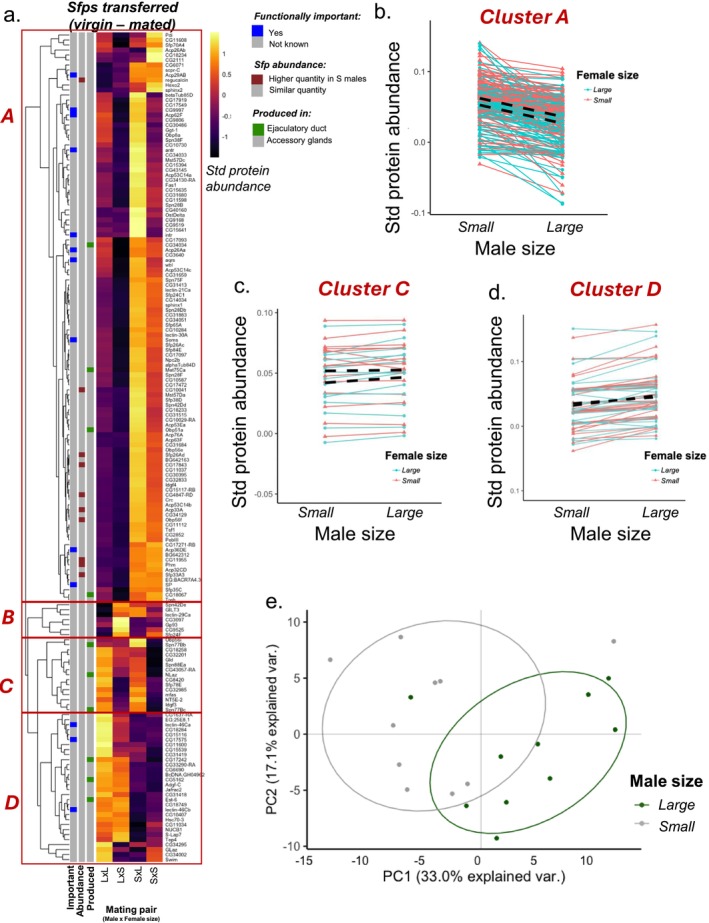 FIGURE 3
FIGURE 3- —Biotechnology and Biological Sciences Research Council10.13039/501100000268
- —CNPq‐Ciência sem Fronteiras10.13039/501100017564
- —Royal Society10.13039/501100000288
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Behavior and Reproduction · Insect and Arachnid Ecology and Behavior · Plant and animal studies
Introduction
1
Male reproduction was traditionally considered inexpensive compared with female reproduction and thus, not a major constraint on male reproductive success (Bateman 1948; Dawkins 2016). This assumption, however, has been overturned by studies showing that sperm and ejaculate resources can be limiting (Dewsbury 1982; Wedell et al. 2002). Seminal fluid proteins (Sfps) which, together with sperm, are abundant in the ejaculate and play a pivotal role in triggering post‐mating responses in the female, can be depleted faster than sperm in the short term, impacting male reproductive success in many species (Hihara 1981; Sirot et al. 2009; Perry et al. 2013; Wigby et al. 2016, 2020; Sanghvi et al. 2025). As a result, it is not surprising that mechanisms regulating Sfp investment have evolved (Cameron et al. 2007; Marcotte et al. 2007; Cornwallis and O'Connor 2009; Reinhardt et al. 2011; Perry et al. 2013). The most detailed knowledge of male Sfp allocation comes from Drosophila melanogastxer , where some studies have shown that males quantitatively and qualitatively modulate the amount of specific Sfps transferred to females (Sirot et al. 2011; Hopkins et al. 2019). Sfps in this species have a range of vital roles in reproduction (Swanson et al. 2001), for example, by governing sperm storage (Neubaum and Wolfner 1999; Tram and Wolfner 1999), supporting oogenesis and ovulation (Soller et al. 1999; Heifetz et al. 2000, 2001), reducing remating likeliness (Chen et al. 1988) and influencing hatching success (Prout and Clark 2000; Chapman 2001).
Ecological conditions can modulate the resources that animals gather and allocate to reproductive traits, making some traits more susceptible to condition‐dependent expression (Cotton et al. 2004). Male reproductive traits have long been the focus of condition‐dependent theory, and a meta‐analysis has shown that seminal fluid quantity is strongly condition‐dependent (Macartney et al. 2019). However, it is still unclear whether condition dependence can act at the molecular level to modulate the expression or allocation of individual Sfps in the males' ejaculate. An important ecological factor that can shape males' ejaculate quality as well as their ability to allocate their ejaculate is early life condition, which has long‐lasting fitness implications for individuals (Stevens et al. 2000; McGraw et al. 2007; Monaghan 2008; Morimoto et al. 2016). In insects, crowding during larval development substantially reduces nutrient availability (Klepsatel et al. 2018), which imposes metabolic and phenotypic constraints on individuals (Than et al. 2020). Individuals experiencing crowded larval environments develop smaller body sizes than individuals from uncrowded larval environments (Monaghan 2008), which influences, in adulthood, the ability of males to compete for mating opportunities (pre‐copulatory sexual selection) and fertilisation (post‐copulatory sexual selection) (Amitin and Pitnick 2007; McGraw et al. 2007). Hence, developmental impacts on adult body size may affect how males modulate their ejaculate, considering that larval density affects accessory gland (AGs) sizes, even after accounting for body size (Bretman et al. 2016). A study that quantified two specific Sfps (sex peptide and ovulin) in D. melanogaster found that large males from uncrowded environments produced these Sfps at higher abundances but transferred them in lower abundances per mating when compared with small males from crowded larval environments (Wigby et al. 2016). This study also found evidence for higher transfer of sex peptide to large (uncrowded environments) females (Wigby et al. 2016), highlighting that early life conditions of both males and females can affect ejaculate quality and quantity, and the ability of males to differentially allocate some Sfps in response to their developmental environment (Wigby et al. 2016). However, the seminal proteome of D. melanogaster is thought to contain at least 292 Sfps (Wigby et al. 2020), but to date, only two Sfps have been studied with respect to developmental conditions. Hence, while past studies have provided some insights into the dynamics between larval density, body size, sperm and few select Sfps (McGraw et al. 2007; Wigby et al. 2016; Zeender et al. 2023), we currently lack an understanding of proteome‐wide changes in seminal fluid due to rearing condition and the social contexts in which matings occur.
To address this problem, we manipulated larval crowding of males and females and used quantitative proteomics to test how early life environment impacts the production, composition and strategic allocation of non‐sperm ejaculate components. Proteomics is the ideal technology to better understand how the seminal fluid might be affected by ecological conditions, and it has been a powerful tool to discover novel seminal fluid proteins in D. melanogaster (Findlay et al. 2008; Sepil et al. 2019). We manipulated larval crowding levels to generate distinct body sizes for both males and females: high larval density to produce small adults, and low larval density to produce large adults. We then paired large and small adult males and females in a fully factorial design to compare the abundances of Sfps produced by large and small males and transferred to large and small females. In D. melanogaster , this manipulation is known to influence physiology (Kapila, Kashyap, Gulati, et al. 2021; Kapila, Kashyap, Poddar, et al. 2021; Morimoto 2022; Morimoto, Wenzel, et al. 2023), reproduction (Morimoto et al. 2017; Narasimhan et al. 2023) and the strength of pre‐ and post‐mating sexual selection (Morimoto et al. 2016). Moreover, it also influences male reproductive anatomy (Morimoto et al. 2022) and ejaculate traits (McGraw et al. 2007; Wigby et al. 2016), which has implications for ejaculate composition and sperm competition. More broadly, larval crowding is an ecologically significant trait for D. melanogaster (Morimoto and Pietras 2020) and can provide biological insights into the responses to developmental conditions in other insect species (reviewed by Than et al. 2020). Based on the literature, we predicted that:
- Sfp production would be influenced by larval crowding, wherein large males would produce higher abundances of Sfps than small males, as was previously observed for select Sfps (Wigby et al. 2016); and
- Small males would transfer relatively higher abundances of Sfps to females, as they are expected to have fewer mating opportunities compared with large males (Wigby et al. 2016). We also hypothesised that this effect would be particularly strong when small males mate with large females because large females are expected to be perceived as more attractive compared to small females (Schlupp et al. 2001; Jerry and Brown 2017).
Materials and Methods
2
Fly Stocks
2.1
We used an outbred stock of D. melanogaster collected in Dahomey (Benin) in West Africa, which has been maintained in large outbred populations (> 1000 individuals) in controlled laboratory conditions with overlapping generations since 1970 (Fowler and Partridge 1989). All stocks were maintained, and all experiments conducted, at 25°C on a 12:12 h (h) light: dark cycle in a non‐humidified room. We used a standard sugar‐yeast‐maize‐molasses medium with excess live yeast granules for experiments, following a previously published recipe (see Appendix S1; Morimoto et al. 2017).
Manipulation of the Larval Crowding
2.2
We followed the protocol established in Morimoto et al. (2016). Briefly, we collected eggs from population cages and manipulated larval crowding levels as follows: (a) the high larval density (small bodied adults) had ~50 larvae/mL of food (~200 larvae per ~4 mL fly food in a 34 mL vial) and (b) the low larval density treatment (large bodied adults) had ~4 larvae/mL of food (~40 larvae in ~10 mL fly food in a 34 mL vial). See Morimoto et al. (2016), and Morimoto, McDonald, and Wigby (2023) for full details. These densities also correspond to normal and high larval densities observed in wild populations in D. melanogaster (Morimoto and Pietras 2020). We will herein refer to the body sizes of adult flies as ‘large’ or ‘small’ to indicate individuals reared in low and high larval densities, respectively. Virgin males and females were collected within 6 h of eclosion and kept in groups of 15–20 individuals within same‐sex and larval density treatment vials for 5–7 days before the onset of experiments. At this age, protein accumulation was maximum; hence, any difference in Sfp abundances would not be due to differences in Sfp production rates between small and large males. We needed unmated males to infer Sfp production by that age and newly mated males to infer the quantities of Sfps being lost during mating. We used the same larval manipulation as in Wigby et al. (2016), whereby they reported the following adult body sizes (mean ± SE): males: small (0.60 ± 0.06) and large (0.87 ± 0.03); females: small (0.814 ± 0.08) and large (1.60 ± 0.06).
Male Ejaculate Profile
2.3
To measure male ejaculate investment in response to the developmental environment, we conducted a fully factorial experiment where large and small males were given the opportunity to mate with large and small females, in all combinations (Figure 1). Mating pairs (N = 15 per combination) were allowed to interact for 2 h, until mating was observed, after which males that successfully mated were transferred to cryotubes immediately after the end of mating and snap frozen with liquid nitrogen and kept at −80°C until dissection. Mating pairs (N = 3) that failed to mate during the 2 h interaction were excluded. The experiment was replicated independently 5 times (N total = 300 [15 mating pairs × 4 mating combinations × 5 replicates]). We also kept a sample of large and small body size virgin males as non‐mating controls (N = 100 [10 males × 2 larval treatments × 5 replicates]). Except for the lack of opportunity to mate, virgin males were treated exactly as the experimental counterparts. We randomly selected five males from each treatment (N = 30 [5 males × 6 treatments]) for dissection. Each sample was kept in ice for 2 min, after which males were placed on a glass slide containing phosphate‐buffered saline (DPBS, pH 7.2, Sigma Aldrich Cat‐No. PC5119). With thin tweezers, AGs and ejaculatory duct were dissected under a stereoscope. They were then placed in empty 1.5 mL Eppendorf tubes and frozen at −80°C for proteomic analysis. AGs and ejaculatory ducts of five males from the same treatment group and replicate were then pooled for proteomic analysis (N = 30 [6 treatments × 5 replicates]). The pooling of the AGs and ejaculatory ducts of five males from the same treatment and replicate introduces variation between samples due to differences among experiments (replicates), individuals and dissections. However, we assumed that this between‐replicate variation would be less pronounced than the variation between treatments, allowing us to test our predictions and detect differences caused by larval crowding.
Schematic representation of the experimental design. (a) We manipulated larval crowding which resulted in adult flies (both males and females) of large (low crowding) and small (high crowding) body sizes. We then crossed males and females in a fully factorial design. Virgin males of both sizes were maintained to ascertain Sfps production. (b) Sfps production was measured as the (standardised) Sfps abundance in AGs and ejaculatory ducts of virgin males. Sfps transfer was measured as the difference between the Sfps abundance in virgin males' AGs and ejaculatory ducts minus the Sfps abundance remaining in the AGs and ejaculatory ducts of mated males.
Proteomics Sample Preparation
2.4
Before proteomic analysis, we macerated the samples with a clean pestle, washed them with 25 μL Pierce RIPA buffer and digested the samples using the standard gel‐aided sample preparation (GASP) protocol (as described in Hopkins et al. 2019; Sepil et al. 2019, 2020). First, we reduced the samples by adding 50 mM DTT and waiting for 10–20 min. Then, we added an equal volume of 40% acrylamide/Bis solution (37.5:1. National Diagnostics) and waited for 30 min to facilitate cysteine alkylation to propionamide. Next, 5 μL of TEMED and 5 μL of 10% APS were added to trigger acrylamide polymerisation. We shredded the resulting gel plug by centrifugation through a Spin‐X filter insert without membrane (CLS9301, Sigma/Corning) and fixed the gel pieces in 40% ethanol/5% acetic acid. This was followed by two rounds of buffer exchange with 1.5 M urea, 0.5 M thiourea and 50 mM ammonium bicarbonate before removal with acetonitrile. The resulting immobilised proteins were left overnight for trypsin digestion (Promega) and the peptides were extracted with two rounds of acetonitrile replacements the following morning. We dried the peptides using Sola SPE columns (Thermo) and finally resuspended them in 2% ACN, 0.1% FA buffer ready for liquid chromatography‐mass spectrometry (LC–MS/MS) analysis. One hundred percent of the dissected material was injected. No protein concentration measurement or sample normalisation was performed due to the low protein amounts involved, as this would have consumed the sample.
LC–MS/MS
2.5
The samples were analysed on the LC–MS/MS platform Dionex Ultimate 3000 and Q‐Exactive mass spectrometers (Thermo) as previously described in Hopkins et al. (2019) and Sepil et al. (2019, 2020). First, the peptides were loaded in 0.1% TFA in 2% ACN onto a trap column (PepMAP C18, 300 μm × 5 mm, 5 μm particle, Thermo), followed by peptide separation on an easy spray column (PepMAP C18, 75 μm × 500 mm, 2 μm particle, Thermo) with a gradient from 2% to 35% ACN in 0.1% formic acid and 5% DMSO. MS spectra were obtained in profile mode with a resolution of 70,000 and an ion target of 3 × 10^−6^. The 15 most intense features were selected for subsequent MS/MS analysis at a resolution of 17,500, a maximum acquisition time of 128 milliseconds, an AGC target of 1 × 10^−5^, an isolation width of 1.6th and a dynamic exclusion of 27 s.
Processing of MS Data
2.6
We imported RAW files into Progenesis QIP (version 3.0.6039.34628) using default settings from previous projects and exported MS/MS spectra as MGF files using the 200 most intense peaks without deconvolution for searching (Hopkins et al. 2019; Sepil et al. 2019, 2020). We chose the D. melanogaster UniProt reference proteome as our search target. We used the search engine Mascot 2.5.1 with the following parameters: 10 ppm precursor mass accuracy, 0.05 Da fragment mass accuracy, oxidation (M), deamidation (N, Q) and propionamide (K) as variable modifications, propionamide (C) as a fixed modification, and two missed cleavage sites. Finally, we applied 1% FDR at the peptide level using the target‐decoy method inherent to Mascot and an ion score cutoff of 20. The resulting data was imported into Progenesis and the Top3 method was used for protein quantification. The peptide intensity‐based quantification data was further cleaned, normalised and processed as described in the following paragraph.
Statistical Analysis
2.7
All analyses were performed using R version 3.0.2 (R Core Team 2012). First, we removed all the proteins that had less than two unique peptides to avoid incorrect protein assignments. Although this is a conservative approach and considerably reduces the total number of proteins analysed, it helps ensure that we do not identify false positives due to peptides shared between proteins. Then, we normalised the data as described by Keilhauer et al. (2015)—a method that we have used previously for normalisation of seminal proteome data (Sepil et al. 2019, 2020). The intensity data were initially log transformed [log2(x + 1)] and for each protein, a standard deviation of their intensity profile was calculated. Then, each protein was ranked according to the standard deviation of their intensity profile, and the bottom 90% of the proteins were marked as the ‘background proteome’. This was used to median‐centre the distribution of each sample.
We paired treatment samples by replicates to obtain the differences in Sfps and performed a paired t‐test across the estimated differences to ascertain statistical significance for each Sfps. Here, our prediction was that Sfps would be in significantly higher quantities in virgin males, and paired t‐tests are ideal for testing this prediction. We applied the Benjamini‐Hochberg procedure to account for multiple testing (Tables S1–S3 for adjusted and unadjusted p‐values). The log2 fold change between the means of the paired male samples and the negative log10 of multiple test corrected p‐values were plotted against each other to create volcano plots. Volcano plots were used to visualise the spread of the data. It was also used to check if seminal fluid proteins were significantly depleted following mating, as has been predicted and observed in previous projects (Sepil et al. 2019, 2020). Proteins were described as Sfps based on the list published by Wigby et al. (2020).
The normalisation process and the assignment of the background proteome rely on the assumption that most of the detected proteins have similar intensities between samples (Keilhauer et al. 2015). This assumption can make biological sense for accessory gland proteomics, as 13% of the detected proteins are known Sfps, and we expect considerable quantitative differences for these compared to structural gland proteins, even though the size of males differs markedly between treatments. Patlar et al. (2025) recently showed that the size of the accessory gland is similar between large and small males in a fruit fly larval‐density manipulation experiment (but see also Morimoto et al. 2022 and references therein). Moreover, in our dataset, for about 90% of the proteins (the background proteome), the standard deviation of their intensity profile was lower than 1.0 [log2(x + 1)], which agrees with this inherent assumption. We also re‐ran some of the analyses described below using the unnormalised data to make sure our interpretation of the results is not an artefact of the normalisation process. This additional analysis is reported in Appendix S2, and confirms that the results are largely the same in both sets of analyses.
We focused the rest of our analyses on previously identified Sfps (Wigby et al. 2020). The heatmaps were made using a Pearson correlation distance metric and plotted using the ‘pheatmap’ package (Kolde 2019), and the data were mean‐centred (standardised) for each protein for better visualisation. Major clusters, if found, were determined by visual inspection. The line plots for the clusters were made using the ‘ggplot2’ package (Wickham 2016), and again used the mean‐centred data. Male size‐related compositional changes in the seminal fluid proteome and male and female size‐related compositional changes in the transferred seminal fluid proteome were assessed using principal component analyses and linear mixed effect models (Bates et al. 2015; Kuznetsova et al. 2017). We used linear mixed‐effects models in all our further analyses because the data were normally distributed and included the random effects of protein name and/or replicate. We applied multiple testing corrections when individual proteins were assessed.
For Sfp production, the initial model included male size as a fixed effect and replicate number as a random effect. For Sfp transfer, the initial model included male size, female size and their interaction as fixed effects, and replicate number as a random effect to control for repeated measurements.
Male size‐related Sfp abundance differences were analysed for all Sfps together using a linear mixed‐effect model. Male and female size‐related transferred Sfp abundance differences were analysed for each Sfp cluster separately again using linear mixed‐effect models. We inferred the abundance of Sfps transferred to the female by subtracting the Sfp abundance of newly mated males from virgin males within the same treatment and replicate as done previously (Sepil et al. 2020). Here, for Sfp production, the initial model included male size as a fixed effect, and protein name and replicate number as random effects. For transferred Sfp abundance differences, the initial model included male size, female size and their interaction as fixed effects and protein name and replicate number as random effects. Model selection was performed by backward stepwise elimination; nonsignificant (p > 0.05) variables were eliminated from the model to arrive at the minimal adequate model. The ‘Database for Visualization and Integrated Discovery’ (DAVID) was used for gene ontology (GO) enrichment analysis (Huang et al. 2009b).
Male size‐related Sfp abundance differences and male and female size‐related transferred Sfp abundance differences were also analysed for each Sfp separately using linear mixed‐effect models. For testing the effect of male size on virgin male Sfp abundances (Sfp production), the initial model included male size as a fixed effect and replicate number as a random effect. For transferred Sfp abundance differences, the initial model included male size, female size and their interaction as fixed effects and replicate number as a random effect. Model selection was performed by backward stepwise elimination, and the resulting p values were corrected for multiple testing using the Benjamini–Hochberg procedure. Sfps that were transferred in significantly different abundances as a response to male size are annotated on the Sfp transfer heatmap, together with Sfps produced in the ejaculatory duct (as opposed to the accessory glands) and Sfps that are known to be functionally important (sex peptide network proteins, ovulin network proteins or proteins linked to sperm competition performance). Finally, we checked whether the functionally important Sfps were over‐represented in any of the clusters using proportion tests.
Results
3
Large Males Produce More Sfps Than Small Males but Have Similar Seminal Proteome Profiles
3.1
From the 30 samples in which five male AG and ejaculatory duct were pooled, we found a total of 2050 proteins. Of these, 1274 had at least two unique peptides and were thus kept in the final dataset. We detected 167 known Sfps out of a total of 292 using the most recent list from Wigby et al. (2020). Of the 167 known Sfps, 135 were significantly more abundant in virgin samples than in mated samples (for each protein, p < 0.033; 0.166 < log2 fold change < 2.729; Data S1 and S2, Figure S1). Only eight Sfps were significantly more abundant in mated versus virgin males (for each protein p < 0.049; −0.21 < log2 fold change < −0.901). From here on, we focused our analyses on the 167 known Sfps to ascertain if their production and transfer were affected by the size of the male, by the size of its female mate or by their interaction.
We first assessed the abundance of Sfps in virgin males in response to our larval crowding manipulation, which resulted in distinct body sizes. The heatmap shows no obvious pattern of abundance change within the Sfp proteome in relation to male size and no clear clustering among the Sfps (Figure 2a). However, there is a small but statistically significant change between treatments, whereby Sfp abundances were higher in large versus small males overall (linear mixed effects model: χ12 = 8.334, p = 0.004; Figure 2b). We investigated whether this effect was driven by some Sfps more than others by testing for significant changes in individual Sfps. However, we found no evidence for male‐size related abundance differences for any of the 167 Sfps in isolation, after multiple test correction (Table S1). These results suggest that large males produce more Sfps than small males, but this effect is not primarily driven by large effects of a small number of Sfps and instead, is driven by relatively small effects in many Sfps.
Condition‐dependent Sfps production. Seminal fluid protein production increases in large males compared to small males. (a) Heatmap showing the abundance of 167 Sfps identified in accessory gland and ejaculatory duct samples of small and large males. Each cell gives the across‐replicate mean for that Sfp in each treatment (n = 5 replicates per treatment). None of the Sfps changed in abundance as a response to male size after multiple test correction. Row annotation provides information relating to whether the Sfp is produced in the accessory glands or ejaculatory duct (green annotation). Pearson correlation was used as the distance metric for the hierarchical clustering. (b) Line plots showing the change in standardised Sfp abundance with male size. The mean standardised abundance of Sfps in small and large males are depicted with red triangles and joined by a blue line. Large males have a statistically significant increase in Sfp abundances compared to small males. (c) Principal component analyses of the seminal fluid proteome in male reproductive tissues. The composition of the seminal fluid proteome is similar between small and large males. Green points represent small male virgin samples (5 replicates) and grey points represent large male virgin samples (5 replicates). Ellipses denote 80% normal probability.
Next, we investigated whether the composition of the seminal proteome differed between large and small males, but we did not find any evidence for it (Figure 2c). While larval crowding impacted the abundances of the Sfps, these changes were consistent across Sfps, hence, the overall composition remained similar. PC1 explained 32.3% of the variation in the data, but this was not associated with the size of the male (linear mixed effects model: χ12 = 0.768, p = 0.381). PC2 explained 20.6% of the variation in the data, but again this was not associated with the size of the male (linear mixed effects model: χ12 = 0.279, p = 0.598). We repeated the above analysis with unnormalised data to ascertain whether the patterns were not an artefact of the normalisation process, but the results remained largely unchanged (see Appendix S2). Together, these results suggest that although larval crowding strongly influences male size, this translates into minor but widespread differences in Sfp abundances before mating.
Seminal Protein Transfer Responds to Male and Female Developmental Environment
3.2
Although large and small males have similar Sfp profiles, they may not necessarily transfer similar amounts to females at mating, especially if the females also vary in size. To test this, we investigated how the abundance of transferred Sfps changed between our mating treatment groups. We performed a hierarchical clustering analysis and the resulting heatmap revealed that there was clustering among the Sfps in their response to male and female size. We identified four higher‐order clusters and analysed them separately to identify distinct patterns of abundance change within the transferred Sfp proteome (Figure 3a). We also checked whether the distribution of functionally important Sfps among the clusters was biased towards any of them.
Seminal fluid protein transfer responds to male size. (a) Heatmap showing the abundance of 167 Sfps transferred to females during mating (n = 5 replicates per treatment). Large males were mated to large females (LxL) or small females (LxS) and small males were mated to large females (SxL) or small females (SxS). Each cell gives the across‐replicate mean for that Sfp in each treatment combination. Row annotation provides information relating to whether the Sfp is produced in the accessory glands or ejaculatory duct (green annotation) and whether the Sfps are known to be functionally important (blue annotation). 14 Sfps (Acp33A, CG10041, CG11955, CG17843, CG4847‐RD, Obp56f, Phm, Sfp26Ad, Acp53Ea, CG31413, CG34129, PebIII, Sfp26Ac, Adgf‐C) were transferred in significantly higher abundances by small males compared to large males, and these Sfps are highlighted using red annotation. Pearson correlation was used as the distance metric for the hierarchical clustering. (b–d) Line plots showing the standardised abundance of Sfps transferred as a response to male and female size for Sfps in cluster A (b), cluster C (c) and cluster D (d). The average change in Sfp abundance for large and small females as a response to male size is depicted with dashed black lines. (e) Principal component analyses of the seminal fluid proteome transferred to females during mating. The composition of the seminal fluid proteome transferred is different for small and large males. Green points represent large male samples (5 replicates mated to small females and 5 replicates mated to large females—10 in total) and grey points represent small male samples (5 replicates mated to small females and 5 replicates mated to large females—10 in total). Ellipses denote 80% normal probability.
Cluster A was the largest with 114 Sfps. This group of Sfps responded significantly to male size and female size, but there was no interaction between the two (Figure 3b; male size only, linear mixed effects models: χ12 = 338.15, p < 0.0001; female size only, χ12 = 51.346, p < 0.0001; interaction, χ12 = 0.08, p = 0.776). Small males transferred a higher abundance of these Sfps compared to large males. Males also transferred a higher abundance of these Sfps to large females compared to small females. Ten of 13 functionally important Sfps were found in Cluster A (sex peptide network proteins: antr, intr, CG9997, aqrs, Sems, SP; ovulin network proteins: Acp26Aa; sperm competition performance proteins: Acp62F, Acp36DE, Acp29AB). However, this ratio was not more than expected based on chance (proportion test: χ12 = 0.114; p = 0.367).
Cluster B was the smallest cluster with eight Sfps (Figure S2). The Sfps in Cluster B did not respond to male size, female size or their interaction. The variation in the abundances that were transferred to the female could not be explained by male or female body size (linear mixed effects models: male size: χ12 = 1.072, p = 0.3; female size, χ12 = 2.404, p = 0.121; interaction, χ12 = 2.204, p = 0.137). None of the functionally important Sfps were found in Cluster B, yet this was not different than expected based on chance (proportion test: χ12 = 0.011; p = 0.543).
Cluster C had 15 Sfps and these were transferred in higher abundances to large females compared to small females (Figure 3c; linear mixed effects model: female size, χ12 = 16.319, p < 0.0001). Male size or the interaction between male and female size did not impact the abundance of the Cluster C Sfps transferred (linear mixed effects model: male size, χ12 = 1.712, p = 0.19; interaction, χ12 = 1.039, p = 0.307). None of the functionally important Sfps were found in Cluster C. Again, this was not different than expected based on chance (proportion test: χ12 = 0.369; p = 0.728).
Finally, Cluster D was the second largest cluster with 30 Sfps. These Sfps were transferred in higher abundances when the males were large as opposed to small (Figure 3d; male size, χ12 = 30.848, p = 0.0001). The transfer of these was not impacted by female size or the interaction between male and female size (linear mixed effects model: female size, χ12 = 0.011, p = 0.915; interaction, χ12 = 2.068, p = 0.15). Three of 13 functionally important Sfps were found in Cluster D (sex peptide network proteins: lectin‐46Ca, lectin‐46Cb, CG17575). However, this ratio was not more than expected based on chance (proportion test: χ12 = 0.007; p = 0.465).
For each cluster, we also conducted a DAVID (Huang et al. 2009b, 2009a) analysis to see if these Sfps were enriched in any function within all the Sfps we identified, but did not detect any enriched classes.
The hierarchical clustering analysis and the analyses of the individual clusters revealed that transferred Sfps do not show a uniform response to changes in male and female size. Instead, we found distinct patterns of transfer between protein groups. This suggests that the composition of the seminal fluid proteome transferred during mating might be different depending on male or female size. We tested this with a PCA (Figure 3e). PC1 explained 33% of the variation in the data. For PC1, we observed separate clustering based on male size, indicating that the composition of the seminal fluid proteome transferred is different between large and small males (linear mixed effects model: χ12 = 8.212, p = 0.004). However, the composition of the seminal proteome transferred did not respond to female size or the interaction between male and female size (linear mixed effects model: female size, χ12 = 2.043, p = 0.152; interaction, χ12 = 0.05, p = 0.822). PC2 explained 17.1% of the variation in the data, but these were not associated with the size of the male, size of the female or their interaction (linear mixed effects model: male size, χ12 = 2.114, p = 0.149; female size, χ12 = 0.431, p = 0.511; interaction, χ12 = 0.002, p = 0.962). Hence, male size was the primary driver of compositional change in Sfps profiles transferred to females, whereas neither female size nor the interaction between female and male size had an effect.
Finally, we tested whether individual Sfps contributed to this compositional change, to try and identify the key Sfps underpinning compositional effects. After correcting for multiple tests, we found that small males transferred 10 Sfps in significantly higher abundances compared to large males (Tables S2 and S3). These Sfps are Acp33A, CG10041, CG11955, CG17843, CG4847‐RD, Obp56f, Phm, Regucalcin, Sfp26Ad and Sfp33A3 and are all within Cluster A. However, none of the other 135 Sfps were transferred in significantly different abundances based on the interaction between male and female size after multiple test correction (Table S2). Re‐analysing the data with raw unnormalised data did not change the results qualitatively (see Appendix S2). Together, these results suggest that during mating, males have fine control over the quantities of Sfps transferred to females, and small males strategically allocate several Sfps in higher abundance compared to large males, and this leads to a compositional change in the seminal fluid proteome transferred.
Discussion
4
Holometabolous insects are sensitive to the ecological conditions experienced during their larval development, which can have long‐lasting effects. Ecological conditions such as resource limitation or landscape fragmentation can lead to larval crowding. Crowding is a major source of ecological stress to the insect larvae because crowding reduces nutrient availability and increases food toxicity, both contributing to longer developmental times, lower larval survival and lower adult body mass and reproductive potential. Despite this, we lack the molecular understanding of how individuals plastically respond to varying larval crowding, especially in relation to reproductive traits. Our data reveal differential allocation and composition of the seminal proteome based on larval crowding in D. melanogaster , driven primarily by males but also partly by females. We found that large (low larval density) adult males have a small but significant increase in seminal proteome production compared to small (high larval density) males, but that they are compositionally similar. In fact, on an individual basis, no Sfps were observed to be produced significantly more by small males. This finding provides only weak support for the hypothesis that small males have limited ejaculate resources compared with large males (prediction 1) (Wigby et al. 2016). However, our results broadly support the hypothesis that small males, who are likely weaker pre‐mating competitors, can maximise their fitness by investing more Sfps in each mating event, whereas large males, which have more opportunities for mating than small males, could benefit from being more conservative at Sfp allocation to avoid ejaculate depletion (prediction 2) (Wedell et al. 2002; Linklater et al. 2007; Sirot et al. 2009). However, Sfp allocation at mating was not uniform, suggesting that males invest subsets of Sfps differentially.
We had expected to observe an increase in Sfp production for large males, broadly consistent with previous observations (Wigby et al. 2016) and built on the assumption that males with overall more resources would invest more in reproduction (prediction 1) (Dewsbury 1982; Grafen 1990; Cotton et al. 2006; De Nardo et al. 2021). Our data supported this prediction: large males produced overall a greater abundance of Sfps than small males (Figure 2c). However, the composition of the produced seminal proteome remained unchanged, suggesting that although larval crowding strongly influences male size, this translates into minor but widespread and consistent differences in Sfp abundances before mating. This lack of compositional change might suggest that the fitness value of individual Sfps does not change with male size—i.e., Sfps that benefit large males benefit small males in the same way. However, the non‐uniform changes to ‘transferred’ Sfps (discussed further below), perhaps argue against this idea. Alternatively, there may be developmental constraints which limit plasticity in seminal proteome composition, especially if large changes in the abundance of individual or groups of Sfps cause a wider reduction in function or effective transfer of the Sfps during mating. This was observed in a previous study where the accumulation of Sfps in the absence of mating led to a change in the seminal proteome composition in old males, yet despite having higher abundances of most Sfps, they were transferred in lower quantities by old unmated males compared to young males (Sepil et al. 2020). Moreover, large reductions in a single Sfp, sex peptide, are known to generate widespread changes in the seminal proteome composition and structure, potentially altering many aspects of ejaculate function (Wainwright et al. 2021).
Adult male AG size, but not testis size, is affected by both larval crowding and the presence of adult males during larval development (Bretman et al. 2016). AG volumes negatively correlate with increasing larval crowding both in absolute and relative terms, accounting for body size differences (Morimoto et al. 2022), although recent studies suggest nuanced interactions with nutrition (Patlar et al. 2025). AG size has been linked directly to increased production and transfer of sex peptide (Wigby et al. 2009), and to increased pre‐ and post‐copulatory success (Bangham et al. 2002; Wigby et al. 2009). Our proteomics data do not allow us to measure AG volume, but our results suggest that the known differences in AG volume derived from larval crowding likely associate with the abundance of the seminal proteome. Our recent study using ecologically relevant larval densities and microcomputing tomography has shown that AG volume tends to decrease for males that experienced high larval crowding. This agrees with sexual selection theory that male AG investment is expected to decrease when the intensity of post‐copulatory sexual selection is higher than a given threshold (Morimoto et al. 2022). However, our past work did not enable us to correlate changes in AG volume with Sfps composition. Future studies combining proteomics with microcomputing tomography will allow us to better ascertain the relationship between AG volume, Sfp yields and compositions directly.
When we assessed the fold‐change difference in the abundance of Sfps in the reproductive tract of virgin and mated males, as a measure of Sfp transfer to females, we confirmed prediction 2 that small males transfer higher abundances of Sfps to females. This is in partial agreement with previous literature. Wigby et al. (2016) used ELISA to quantify the production and transfer of two Sfps—sex peptide (SP) and ovulin—in a fully factorial experiment that manipulated larval crowding levels similar to our present design. In their study, they found that large males produced higher abundances of SP and ovulin but also reported a non‐significant trend for large (low crowding) males to transfer proportionally lower abundances of SP in each mating. Supporting this, our proteomic analysis confirmed that large males tend to produce higher abundances of Sfps overall, although we did not have evidence of significantly higher production of SP and ovulin in large vs small males. Wigby et al. (2016) also reported that males tend to transfer proportionally a higher abundance of SP to larger females, although they did not compare the transfer of ovulin across females from different sizes. Our results did not show evidence that large females receive higher abundances of Sfps consistently and the composition of the seminal fluid transferred did not vary as a response to female size. However, Sfps in two of the Clusters A and C had a trend of being transferred in higher abundances to large females. Notably, both SP and ovulin are Sfps from Cluster A. Using ELISA, Wigby et al. (2016) reported males transferred more SP to large females, in agreement with our findings of Cluster A to which SP belongs.
One possible explanation for some of the discrepancies is the sensitivity of the methods used to measure Sfps. The ELISA approach by Wigby et al. (2016) was highly sensitive to their target Sfps at the expense of global resolution of the seminal proteome. Conversely, our approach was focused on understanding the response of the entire seminal proteome to larval crowding effects. As a result, Wigby et al. (2016) approach likely captured relatively smaller changes in the production and transfer of their target Sfps than our method could capture. However, it is important to note that in our study, Sfps transferred were not homogeneous across the seminal fluid proteome, suggesting that males fine‐tune and strategically control the transfer of specific Sfps. Specifically, Sfps from Clusters A and C, but not Clusters B and D, were transferred in higher abundances to large females, whereas Sfps of Cluster A were transferred in higher abundances by small males compared to large males, but the reverse was true for Sfps in Cluster D. Taken together, the results strengthen the idea that males have control of specific components of their ejaculate and allocate them strategically. Cluster A—the largest cluster—supported the prediction of small males transferring Sfps in a higher abundance compared to large males, and of males transferring a higher abundance to large females (Figure 3). The 10 Sfps transferred in higher abundances by small males compared to large males were: Acp33A, CG10041, CG11955, CG17843, CG4847‐RD, Obp56f, Phm, Regucalcin, Sfp26Ad, Sfp33A3. None of these proteins has known or clearly defined reproductive functions. However, Acp33A is associated with sperm competitiveness in D. melanogaster : both ‘offence’ (the displacement of previous male's sperm) and ‘defence’ (protection of stored sperm against displacement) (Fiumera et al. 2005; Ravi Ram and Wolfner 2007), and hence might be boosted by small males in anticipation of future or past sperm competition. In particular, the high larval density experienced by small males could be an indicator of high population density, and hence high sperm competition risk (Bretman et al. 2016), signalling the need to boost the transfer of sperm competition‐related Sfps. The role of Obp56f is unclear, but it does not seem to be required for female fecundity or the induction of female refractoriness after mating (Brown et al. 2023). Regucalcin is a Ca^2+^ binding protein which is transferred more by males that mate frequently than by males at their first mating (Sepil et al. 2020); however, its function as an Sfp also remains unknown. Other Sfps with known functions in sperm competition, such as Acp36DE, Acp62F, Acp29AB and SP, were not found to be transferred in significantly higher quantities by small males, yet all these Sfps were, like Acp33A, within Cluster A. Hence, the trend of small males transferring a higher quantity than large males still holds. The multiple test correction we applied is highly stringent/conservative and might have limited our ability to detect important quantitative differences between individual Sfps. Therefore, we used the clustering approach to complement our analyses.
Once transferred to females, Sfps trigger a wide range of pathways which modulate female immunity, nutrition, vision and overall behaviour (Griffith 2008; Yapici et al. 2008; Häsemeyer et al. 2009; Rubinstein and Wolfner 2013; Sun and Spradling 2013; Peng et al. 2024), through a wide range of communication modes (Kortsmit et al. 2024; Zelinger et al. 2024). We did not directly measure Sfps in the female reproductive tract and therefore, it will be important for future studies to ascertain whether the reproductive tract of small and large females provide an additional selective barrier that exacerbates or homogenises the response to strategic ejaculate investments. This is because we know that larval crowding plays a major role in shaping female seminal receptacle morphology (Amitin and Pitnick 2007) and that Sfps interact with and within the seminal receptacle to facilitate sperm storage and release and to trigger a wide range of post‐mating behaviours (Guan et al. 2023). We also note that because males were frozen soon after mating, our data does not include mating‐induced changes in Sfp expression that might affect subsequent matings. Understanding the dynamics and replenishment of Sfp expression, and its influence on fertilisation, is an essential future step to better understanding proteomic changes in the ejaculate.
Likewise, it will be important to understand how condition‐dependent ejaculate investment and any putative interactions between Sfps and the female reproductive tract, translate into fitness differences between large and small males mating with large or small females. For D. melanogaster , the larval density does not affect male remating intervals, whilst females from low‐density environments remate more rapidly (Amitin and Pitnick 2007). A recent study found no differences in mate preference, female mating latency, nor female–male interactions in individuals which experienced diluted larval diets, a treatment which mimics in some ways the developmental effects observed in crowded cultures (e.g., slower growth, delayed development) (De Nardo et al. 2021). However, here it was observed that small males transferred more sperm to non‐virgin females, displaced a larger volume of previously deposited sperm, and achieved higher paternity share per mating than large males (De Nardo et al. 2021). This suggests that smaller males may compensate for reduced mating successes by investing more sperm in each mating event. Our study supports the idea of overall higher ejaculate investment by small males, because we found that small males transferred higher abundances of Sfps to females overall, albeit this trend not being homogeneous across the seminal fluid proteome. Another important point to mention is that our proteomics approach allows us to compare relative abundances for a large pool of proteins for which no specific antibodies exist, as is the case for most Sfps. However, our approach is indirect and only provides relative quantification of protein abundances. Future research that builds on our findings by measuring Sfps in absolute quantities, for both production and transfer to females, will provide scope for deeper mechanistic insights.
Our manipulation of larval crowding to generate phenocopied small and large body sizes likely influenced how males and females interact. Schang et al. (2023) recently showed that phenocopied males might exhibit exaggerated responses compared to natural body size phenotypes, including effects on mating and offspring siring. Therefore, we cannot completely rule out that our manipulations led to distinctive patterns of Sfp production and transfer that would not typically be observed in natural body size phenotypes. However, several additional factors prevent us from making direct comparisons between our phenocopies and those of Schang et al. (2023). For example, flies in our study were derived from a ‘Dahomey’ stock with overlapping generations, whereas those in Schang et al. (2023) were derived from the ‘Ives’ stock with non‐overlapping generations. As a result, both the genetic background and life cycle regimes differed, likely influencing the traits expressed in our phenocopies. It is also worth noting that the direction of the effect sizes was the same between large and small natural and phenocopied males in Schang et al. (2023) (see their Figures 1, 2, 3), with one exception. Hence, even if our larval crowding manipulation and body size phenocopies affected the magnitude of our observed effects, it is unlikely that they altered our interpretation and conclusions.
Conclusions
5
Evidence supporting plasticity in sperm and non‐sperm production and allocation patterns is ubiquitous (Simmons 2002; Wedell et al. 2002; Hodgson and Hosken 2006; Cameron et al. 2007; Birkhead et al. 2009; Wigby et al. 2009; Sirot et al. 2011; Perry et al. 2013). Using label‐free quantitative proteomics, our study shows evidence for condition‐dependent ejaculate allocation which supports broad predictions from condition‐dependent sexual selection theory in relation to reproductive investment. These findings advance our understanding of how early‐life ecological conditions can modulate ejaculate allocation in adulthood. It will be important in future to understand the trade‐offs associated with the seminal strategies revealed here, and how it relates to sperm allocation, other life history traits and ultimately fitness. It is likely that larval density effects on male reproductive traits are common among insects. Understanding the impacts of larval crowding on ejaculate proteomics could be useful for applications that involve sterile males' release in biocontrol programmes (Sirot et al. 2011; Wei et al. 2016). Optimising developmental conditions to maximise male competitiveness could contribute to the success of such programmes. Overall, our findings advance our knowledge of the seminal proteome, which could lead to further advancements in understanding this in other organisms including humans (Rowe and Houle 1996; Wigby et al. 2020).
Author Contributions
R.v.H. led the writing of the original draft of the manuscript and contributed to data visualisation. R.K., P.D.C., R.F. and B.M.K. conducted the sample preparations and proteomics runs and contributed to the data analysis of the proteomics data and revision of the manuscript. I.S. co‐led the writing of the original draft of the manuscript and data analysis and made plots for data visualisation. S.W. and J.M. designed the experiment and contributed to writing the original draft of the manuscript and data visualisation. J.M. ran the experiment, collected and dissected the samples for proteomics, and co‐led the writing of the original draft of the manuscript. I.S. and J.M. had shared oversight of the project.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Data S1: mec70101‐sup‐0001‐DataS1.xlsx.
Data S2: mec70101‐sup‐0002‐DataS2.xlsx.
Appendix S1 and S2: mec70101‐sup‐0003‐Appendix.docx.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Amitin, E. G. , and S. Pitnick . 2007. “Influence of Developmental Environment on Male‐ and Female‐Mediated Sperm Precedence in Drosophila melanogaster .” Journal of Evolutionary Biology 20, no. 1: 381–391. 10.1111/j.1420-9101.2006.01184.x.17210031 · doi ↗ · pubmed ↗
- 2Bangham, J. , T. Chapman , and L. Partridge . 2002. “Effects of Body Size, Accessory Gland and Testis Size on Pre‐ and Postcopulatory Success in Drosophila melanogaster .” Animal Behaviour 64, no. 6: 915–921. 10.1006/anbe.2002.1976. · doi ↗
- 3Bateman, A. J. 1948. “Intra‐Sexual Selection in Drosophila .” Heredity 2, no. 3: 349–368. 10.1038/hdy.1948.21.18103134 · doi ↗ · pubmed ↗
- 4Bates, D. , M. Mächler , B. Bolker , and S. Walker . 2015. “Fitting Linear Mixed‐Effects Models Using lme 4.” Journal of Statistical Software 67, no. 1: 1–48. 10.18637/jss.v 067.i 01. · doi ↗
- 5Birkhead, T. R. , D. J. Hosken , and S. Pitnick , eds. 2009. Sperm Biology – An Evolutionary Perspective. Elsevier. 10.1016/B 978-0-12-372568-4.X 0001-8. · doi ↗
- 6Bretman, A. , C. Fricke , J. D. Westmancoat , and T. Chapman . 2016. “Effect of Competitive Cues on Reproductive Morphology and Behavioral Plasticity in Male Fruitflies.” Behavioral Ecology 27, no. 2: 452–461. 10.1093/beheco/arv 170.27004011 PMC 4797378 · doi ↗ · pubmed ↗
- 7Brown, N. C. , B. Gordon , C. E. Mc Donough‐Goldstein , et al. 2023. “The Seminal Odorant Binding Protein Obp 56g Is Required for Mating Plug Formation and Male Fertility in Drosophila melanogaster .” e Life 12: e 86409. 10.7554/e Life.86409.38126735 PMC 10834028 · doi ↗ · pubmed ↗
- 8Cameron, E. , T. Day , and L. Rowe . 2007. “Sperm Competition and the Evolution of Ejaculate Composition.” American Naturalist 169, no. 6: E 158–E 172. 10.1086/516718.17479456 · doi ↗ · pubmed ↗