A comprehensive compilation of data on the association between XRCC3 polymorphisms and thyroid cancer susceptibility
Mahdi Khosravi-Mashzi, Seyed Masoud HaghighiKian, Amirhosein Naseri, Alireza Negahi, Mohammad Vakili-Ojarood, Bahareh Mehdikhani, Rezvan Nezameslami, Alireza Nezameslami, Amirhossein Rahmani, Amihossein Shahbazi, Amirmasoud Shiri, Hossein Neamatzadeh

TL;DR
This study finds that certain genetic variations in the XRCC3 gene may increase thyroid cancer risk, especially in specific populations.
Contribution
The study provides a meta-analysis clarifying the association between XRCC3 polymorphisms and thyroid cancer risk with variant- and ethnicity-specific insights.
Findings
XRCC3 rs1799794 is linked to increased thyroid cancer risk under recessive models.
rs1799796 may protect against thyroid cancer in Asian populations.
rs861539 shows no significant association with thyroid cancer risk.
Abstract
Polymorphisms in the XRCC3 gene, a key component of homologous recombination repair, have been studied for their potential role in thyroid cancer susceptibility. However, published findings remain inconsistent across populations and genetic variants. This meta-analysis aimed to clarify the associations between XRCC3 polymorphisms and thyroid cancer risk, with emphasis on variant- and ethnicity-specific effects. A systematic search of PubMed, EMBASE, Scopus, CNKI, and other databases up to July 10, 2025 identified case–control studies reporting genotype distributions of XRCC3 polymorphisms in thyroid cancer. Pooled odds ratios (ORs) and 95% confidence intervals (CIs) were calculated under fixed- or random-effects models. Subgroup analyses by ethnicity, sensitivity tests, and publication bias assessments (Egger’s and Begg’s tests) were performed. Twenty-four eligible studies were…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
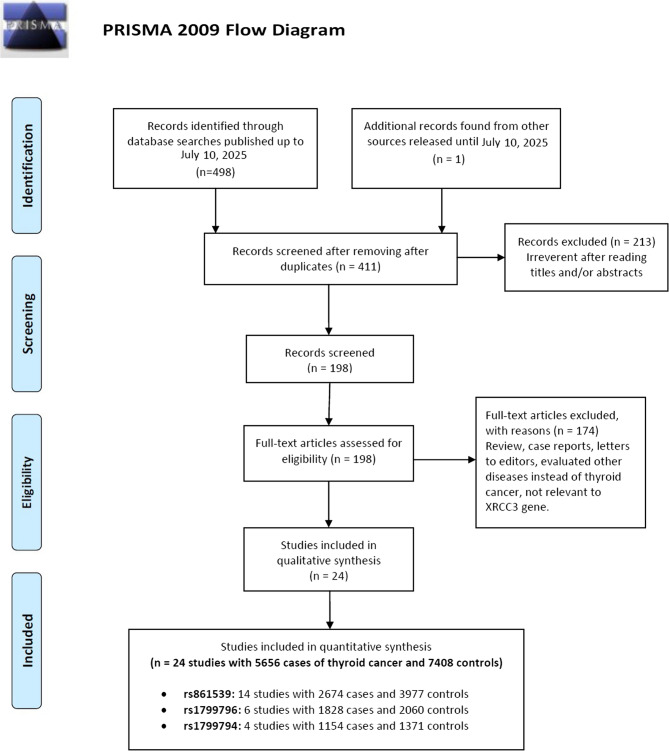 Figure 1
Figure 1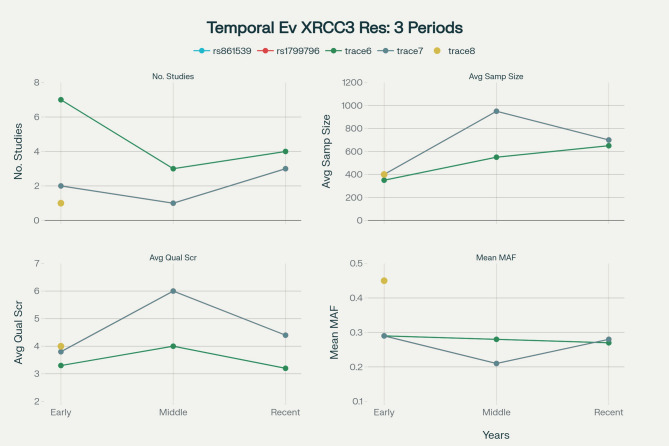 Figure 2
Figure 2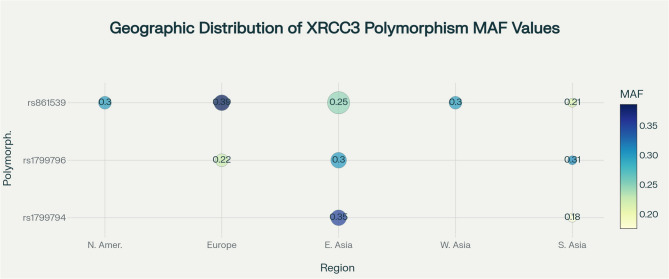 Figure 3
Figure 3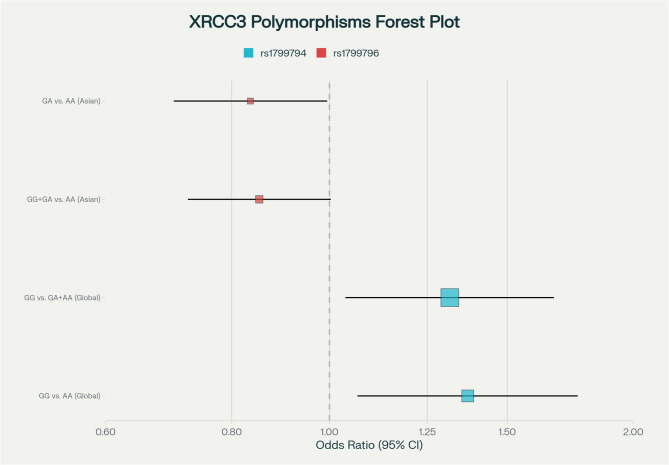 Figure 4
Figure 4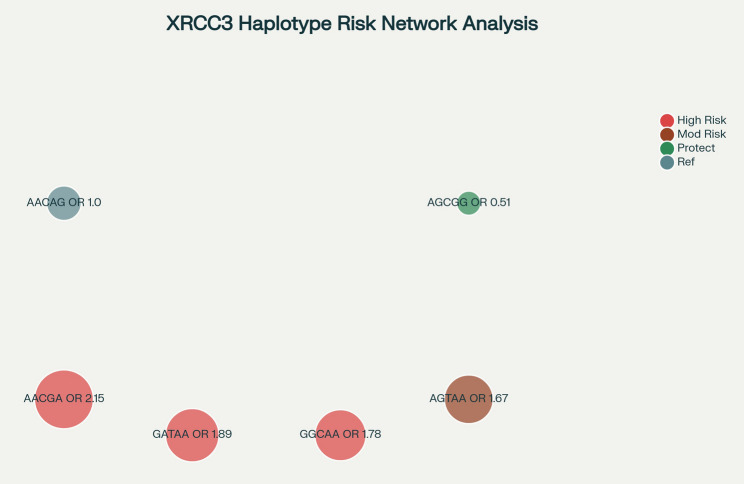 Figure 5
Figure 5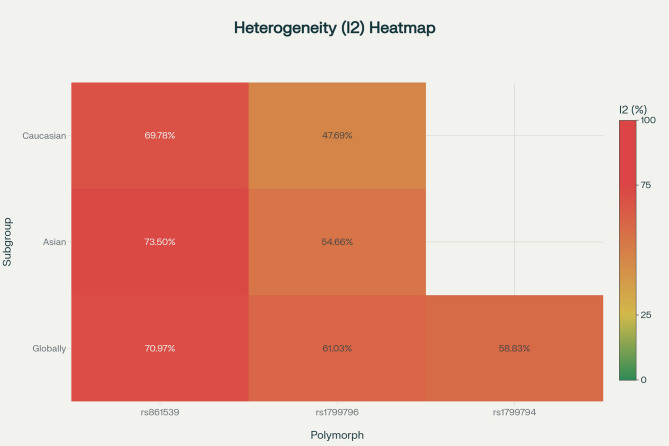 Figure 6
Figure 6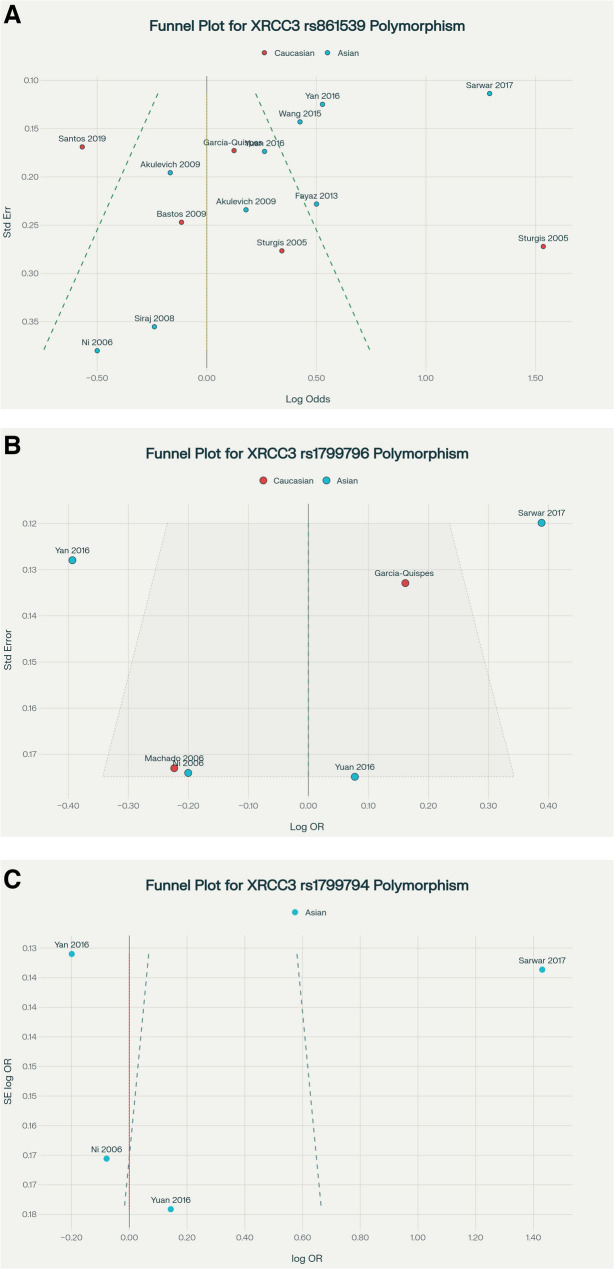 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDNA Repair Mechanisms · BRCA gene mutations in cancer · PARP inhibition in cancer therapy
Introduction
Thyroid cancer is an increasingly significant global health challenge, ranking as the tenth most common malignancy worldwide and disproportionately affecting women, who are diagnosed at nearly three times the rate of men [1, 2]. Globally, an estimated 821,214 new cases and 47,507 deaths were reported in 2022, with age-standardized incidence rates of 13.60 per 100,000 in women compared to 4.60 per 100,000 in men, highlighting a marked gender disparity [1, 3]. In the United States alone, approximately 44,000 new cases are projected for 2025, with mortality remaining low at about 0.5 per 100,000 annually [4]. Over recent decades, incidence rates have steadily increased worldwide—particularly in high-income countries—primarily due to improved detection methods such as high-resolution ultrasound and fine-needle aspiration biopsy [5]. Despite this rise in incidence, mortality rates have remained stable or slightly declined, reflecting advances in early diagnosis and treatment [6]. The overall 5-year relative survival rate remains excellent at 98.4%, with localized disease showing survival rates as high as 99.9%. However, projections indicate the global burden will continue to grow substantially, with an estimated 1,100,000 new cases and 91,000 deaths anticipated by 2050 [3].
DNA repair mechanisms are essential for preserving genomic stability and preventing carcinogenesis. The XRCC3 gene, located on chromosome 14q32.3 and spanning approximately 21 kb with 10 exons, encodes a key protein involved in homologous recombination repair (HRR) of DNA double-strand breaks (DSBs) [7]. XRCC3 maintains HRR fidelity by stabilizing heteroduplex DNA intermediates during repair and directly interacting with Rad51, the human homolog of this protein (HsRad51) [7, 8]. Deficiency in XRCC3 impairs HRR capacity and increases cellular sensitivity to ionizing radiation, DNA cross-linking agents, and alkylating drugs [9]. Loss of XRCC3 function has been linked to chromosomal instability and defective processing of recombination intermediates, thereby potentially facilitating oncogenesis [10].
Several single nucleotide polymorphisms (SNPs) within XRCC3 have been investigated for their association with cancer susceptibility, including thyroid cancer [7]. Among these, the rs861539 (Thr241Met) variant—resulting in a threonine-to-methionine substitution, replacing a hydrophilic residue with a hydroxyl group by a hydrophobic residue containing a methyl-sulfur moiety—has attracted considerable attention for its potential to alter protein structure and function [11]. Other polymorphisms, such as rs1799794 and rs1799796, have also been studied, with emerging evidence suggesting their association with thyroid cancer in specific populations [7, 8, 12]. Haplotype analyses have identified risk-associated combinations (e.g., AACGA, AGTAA, GATAA, GGCAA) as well as protective haplotypes, such as AGCGG [13].
However, research on XRCC3 polymorphisms and thyroid cancer risk has yielded inconsistent and sometimes contradictory findings, underscoring significant methodological challenges [7]. A common limitation in case-control studies is small sample size—often fewer than 200 cases—resulting in insufficient statistical power to detect modest genetic effects and increasing susceptibility to both type I and type II errors [12, 14]. Population stratification poses another critical concern; inadequate adjustment for ancestry-related genetic differences between cases and controls can generate spurious associations [15]. Many studies fail to rigorously control for population substructure, introducing confounding that undermines result validity [16].
Deviations from Hardy–Weinberg equilibrium (HWE) in control groups are frequently reported in XRCC3 research, and meta-analyses [8, 12] have shown that excluding HWE-violating studies can significantly alter effect estimates. Such deviations often result from genotyping errors, population admixture, or selection bias in control recruitment. Methodological heterogeneity—including differences in genotyping platforms, control selection criteria, and statistical models—further contributes to inter-study variability, complicating synthesis and interpretation [7, 17]. Publication bias is also a concern, as smaller studies are more likely to report significant associations, potentially skewing the literature toward positive findings [18]. Moreover, most investigations focus on individual SNPs rather than haplotype or multi-locus analyses, which may more accurately capture the functional impact of genetic variation [19]. Ethnic and geographic differences in XRCC3 polymorphism distributions are well documented, with substantial variation in allele frequencies and effect sizes across populations [16]; however, many meta-analyses underrepresent non-European cohorts, limiting the generalizability of their conclusions. Differences in linkage disequilibrium (LD) patterns add further complexity, as apparent associations with specific SNPs may instead reflect the influence of nearby correlated variants.
In light of these challenges, the present comprehensive meta-analysis aims to systematically evaluate the relationship between XRCC3 polymorphisms and thyroid cancer susceptibility on a global scale. By applying rigorous inclusion criteria, employing robust statistical methods to assess heterogeneity, and conducting detailed subgroup analyses stratified by ethnicity, study design, and tumor characteristics, this study seeks to clarify the true contribution of XRCC3 genetic variation to thyroid cancer risk. The findings will provide essential insights for genetic risk stratification, counseling, and the development of personalized prevention and management strategies, while highlighting priority areas for future research in thyroid cancer genetic epidemiology.
Materials and methods
Publication search
This meta-analysis did not require ethical approval or informed consent [20], as it exclusively utilized data from previously published studies. A comprehensive literature search was independently conducted by two reviewers across a wide range of international and regional databases up to July 10, 2025. These databases included PubMed (MEDLINE), EMBASE, Web of Science, Elsevier, Google Scholar, ScienceDirect, SciELO, Europe PMC, ResearchGate, the Circumpolar Health Bibliographic Database (CHBD), the Cochrane Library, Current Contents, Linguamatics, WanFang Data, the China Science and Technology Journal Database (CSTJ), the Chinese Scientific Journals Database (VIP), the Chinese Biomedical Literature Database (CBD), the China National Knowledge Infrastructure (CNKI), the Scientific Information Database (SID), PsycINFO, LILACS, ClinicalTrials.gov, ProQuest Dissertations & Theses Global, Scopus, OpenGrey, the WHO Global Index Medicus, Global Health, BIOSIS Previews, and MEDLINE In-Process. The search strategy aimed to identify all case–control studies investigating associations between XRCC3 gene polymorphisms and thyroid cancer risk. It employed a combination of Medical Subject Headings (MeSH) and relevant keywords related to thyroid cancer (e.g., “Thyroid Cancer,” “Papillary Thyroid Cancer,” “Medullary Thyroid Cancer”) and genetic variation (e.g., “XRCC3,” “X-Ray Repair Cross Complementing 3,” “SNP,” “Polymorphism,” “Mutation”). No language restrictions were applied, with studies published in non-English languages included if genotype data were extractable. To ensure thorough coverage, reference lists of all included studies and pertinent reviews were manually screened to identify additional records not captured by database searches. When multiple publications reported overlapping datasets, only the most complete or recent study with the largest sample size was retained to prevent duplication. This extensive search approach—characterized by broad database inclusion, multilingual consideration, and systematic manual cross-referencing—was designed to capture the widest possible range of eligible studies, identify data on less-studied variants, and ensure representative inclusion of diverse populations worldwide.
Inclusion and exclusion criteria
Eligible studies met the following criteria: (1) original case-control design involving human participants; (2) investigation of the association between XRCC3 gene polymorphisms and thyroid cancer risk; (3) provision of genotype and/or allele frequency data for both cases and controls sufficient for the calculation of pooled odds ratios (ORs) and 95% confidence intervals (CIs); and (4) evaluation of at least one XRCC3 SNP. Studies were excluded if they met any of the following conditions: (1) the publication was a review, meta-analysis, editorial, commentary, conference abstract, or a non-human or in vitro study; (2) the study employed a family-based or linkage analysis design rather than a case-control approach; (3) genotype frequency data were missing or inadequate for analysis; (4) the data overlapped with another included study, in which case the most comprehensive or up-to-date version was selected; or (5) the study addressed thyroid cancer incidence without evaluating genetic associations. In instances where critical genotype data were not reported, study authors were contacted via email to obtain the missing information. This selection process ensured that only high-quality, relevant studies were included in the final analysis.
Data extraction
Data extraction was conducted independently by two researchers using a standardized approach. Any discrepancies were resolved through discussion and consensus. Extracted data included the first author’s name, year of publication, ethnicity of study populations (categorized as Asian, Caucasian, African, or mixed), country of origin, genotyping methods used, sample sizes of case and control groups, genotype distributions in both groups, minor allele frequency (MAF) in controls, and HWE status in the control group. For studies involving multiple ethnic groups, subgroup-specific data were extracted separately. The term “mixed” was used when participant ancestry was heterogeneous or not clearly defined. In cases where critical data—such as genotype frequencies—were missing, study authors were contacted to provide the necessary information. This thorough data collection process ensured completeness and consistency across all included studies.
Statistical analysis
To assess the association between XRCC3 gene polymorphisms and thyroid cancer susceptibility, pooled odds ratios (ORs) with 95% confidence intervals (CIs) were calculated under five genetic models: allele contrast (B vs. A), homozygote comparison (BB vs. AA), heterozygote comparison (BA vs. AA), dominant model (BB + BA vs. AA), and recessive model (BB vs. BA + AA). Statistical significance was evaluated using Z-tests, with two-tailed p-values < 0.05 considered significant. Heterogeneity across studies was assessed using the Cochrane Q-test (with p < 0.10 indicating significant heterogeneity) and quantified with the I² statistic. A fixed-effects model (Mantel-Haenszel method) was used when heterogeneity was low (I² ≤ 50%), while a random-effects model (DerSimonian and Laird method) was applied for higher heterogeneity [21]. Subgroup analyses were performed based on ethnicity, genotyping method, and study quality to explore potential sources of heterogeneity. Where data permitted, meta-regression analyses were also conducted. Sensitivity analyses were conducted by sequentially excluding individual studies and by omitting those not in HWE (assessed using Pearson’s chi-square test with p < 0.05 indicating deviation). Publication bias was examined both visually using Begg’s funnel plots and statistically using Egger’s regression test, with p < 0.05 suggesting significant bias. All statistical analyses were conducted using Comprehensive Meta-Analysis (CMA) software, version 2.0 (Biostat, USA).
Results
Study characteristics
The study selection process is summarized in Fig. 1. A comprehensive search initially identified 498 potentially relevant articles. After removing duplicates and screening titles and abstracts, 146 articles were Full-text reviewed for eligibility. Of these, 79 were excluded for irrelevance or insufficient data. Ultimately, 14 unique case-control studies [19, 22–33] were included in this meta-analysis, encompassing a total of 5,656 thyroid cancer cases and 7,408 controls across 24 polymorphism-specific datasets involving three XRCC3 variants. Detailed study characteristics and sample distributions are presented in Table 1. The rs861539 polymorphism was the most extensively studied, covered in 14 datasets with 2,674 cases and 3,977 controls, followed by rs1799796 (6 datasets; 1,828 cases, 2,060 controls), and rs1799794 (4 datasets; 1,154 cases, 1,271 controls) (Table 1). Sample sizes varied widely, ranging from 37 cases with 227 controls (Siraj 2008) to 456 cases with 400 controls (Sarwar 2017). PCR-RFLP was the predominant genotyping method (used in 75% of studies), while more recent investigations (Yan 2016, Yuan 2016, Sarwar 2017) utilized advanced platforms such as MassARRAY and ARMS-PCR, reflecting technological evolution. Quality assessment revealed considerable methodological heterogeneity, with scores ranging from 2.0 to 6.0 based on sample size, HWE adherence, and control matching. High-quality studies—including Ni 2006, Akulevich 2009, García-Quispes 2011, and Yuan 2016—featured comprehensive multi-polymorphism analyses and population-based control selection. Notably, advances in technology coincided with increased HWE violations, indicating emerging challenges in population stratification and quality assurance (Table 1).Fig. 1. Flow diagram of the study selection process for the meta-analysis
Table 1. Main characteristics of studies evaluated XRCC3 polymorphisms and thyroid cancerFirst Author/YearCountry (Ethnicity)SOCGenotyping MethodCase/ControlCase/Control RatioCasesControlsMAFHWEGenotypeAlleleGenotypeAllelers861539 CC
CT
TT
C
T
CC
CT
TT
C
T Sturgis 2005 [22]USA(Caucasian)HBPCR-RFLP134/1610.832456920159109836018226960.2980.164Sturgis 2005 [22]USA(Caucasian)HBPCR-RFLP79/1610.4913429169761836018226960.2980.164Ni 2006China (Asian)NSPCR-RFLP191/2010.95017912037012181200382200.0490.457Siraj 2008 [24]KSA(Asian)HBPCR-RFLP37/2270.16318127482697105252991550.3410.666Bastos 2009 [25]Portugal(Caucasian)HBPCR-RFLP109/2140.5093944261229671114292561720.4010.113Akulevich 2009 [26]Japan(Asian)PBPCR-RFLP120/1980.606535116157838289272531430.3610.716Akulevich 2009 [26]Japan(Asian)PBPCR-RFLP132/3980.33255651217589161192455142820.3540.277García-Quispes 2011 [27]Spain(Caucasian)HBPCR-RFLP207/2480.83596882328013494119353071890.3810.786Fayaz 2013 [28]Iran(Asian)PBPCR-RFLP161/1830.8807176142181041026813272940.2560.719Wang 2015 [29]China(Asian)HBPCR-RFLP276/5520.5001618431406146362150408742300.208≤ 0.001Yan 2016 [30]China(Asian)HBMassARRAY403/2761.4602551262263617014397363831690.3060.004Yuan 2016 [31]China(Asian)HBMassARRAY183/3670.499956424254112232115205791550.2110.254Sarwar 2017 [19]Pakistan(Asian)HBARMS-PCR456/4001.1402771097066324927385426311690.211≤ 0.001Santos 2019 [33]Portugal(Caucasian)HBPCR-RFLP186/3910.4761063644248124209701124882940.376≤ 0.001rs1799796 AA
AG
GG
A
G
AA
AG
GG
A
G Machado 2006Spain(Caucasian)HBPCR-RFLP207/2480.835115741830411014010083801160.2330.048Ni 2006China(Asian)NSPCR-RFLP181/2010.900839172571058198222601420.3530.341Garcia-Quispes 2011 [27]Spain(Caucasian)HBPCR-RFLP398/5780.68923614517617179367179329132430.2100.105Yuan 2016 [31]China(Asian)HBMassARRAY183/3670.499907518235111194145285332010.2730.899Yan 2016 [30]China(Asian)HBMassARRAY403/2661.51521315931585221136113173851470.2760.310Sarwar 2017 [19]Pakistan(Asian)HBARMS-PCR456/4001.14028410468672240212128605522480.310≤ 0.001rs1799794 AA
AG
GG
A
G
AA
AG
GG
A
G Ni 2006China(Asian)NSPCR-RFLP191/2010.9506681442131696294452181840.4580.411Yuan 2016 [31]China(Asian)HBMassARRAY183/3670.499778422238128184147365152190.2980.406Yan 2016 [30]China(Asian)HBMassARRAY324/3031.06916412733455193202161405652410.2990.345Sarwar 2017 [19]Pakistan(Asian)HBARMS-PCR456/4001.140289907766824429765386591410.176≤ 0.001Abbreviations: SOC Source of Control, HB Hospital Based, PB Population Based, NS Not Stated, NP Not Provided, PCR-RFLP Polymerase Chain Reaction-Restriction Fragment Length Polymorphism, ARMS-PCR Amplification Refractory Mutation System PCR, MAF Minor Allele Frequency, HWE Hardy-Weinberg Equilibrium
Temporal evolution of research quality
The temporal trend analysis of XRCC3 polymorphism research from 2005 to 2019 reveals an inverse relationship between study scale and methodological quality. Early studies (2005–2009) generally featured smaller sample sizes (mean 337 cases) but demonstrated higher quality, evidenced by the absence of HWE violations for rs861539 and an average quality score of 4.29. The middle period (2010–2015) represented the peak of methodological rigor, with an average quality score of 4.5 and 75% HWE compliance. In contrast, more recent studies (2016–2019), although involving larger cohorts, exhibited a decline in quality, with the average quality score dropping to 3.6 and HWE compliance falling to 50%. This reduction in quality coincided with a shift from PCR-RFLP methods to multi-platform genotyping techniques, which was accompanied by reported HWE violations in several recent studies (e.g., Wang 2015, Yan 2016, Sarwar 2017, Santos 2019). Figure 2 visualizes these temporal dynamics across four key metrics—number of studies, average sample size, average study quality score, and mean MAF—for multiple genetic markers including rs861539, rs1799796, and additional genetic traces (trace 6–8). Traces represent distinct genetic polymorphisms, with named traces indicating specific variants and numbered traces representing additional variants for comparison. These results highlight changing research activity, quality, and genetic variant frequencies over time, emphasizing the need to strengthen quality control in current genetic studies despite scientific progress.Fig. 2. Temporal evolution of XRCC3 polymorphism research, showing trends in study count, average sample size, quality score, and MAF across three time periods. Blue lines represent rs861539, red lines represent rs1799796, and cyan/green lines or points represent other genetic variants (traces 6–11). Top panels display study numbers and sample sizes, while bottom panels show quality scores and MAF values
Population-specific genetic architecture and geographic risk stratification
Geographically, the research is heavily concentrated in China, which accounted for 28.6% of studies (4 out of 14) and contributed the largest participant cohorts. Ethnically, Asian populations dominate the dataset, representing 70.8% (17 out of 24 studies), with Caucasians comprising 29.2%. Overall, eight countries contributed, but recent research has consolidated participation to five countries, potentially limiting future population-specific insights. Crucially, no African, South American, or indigenous populations were included, highlighting significant representation gaps that constrain the global generalizability of findings (Fig. 3). The key XRCC3 polymorphisms exhibit distinct ethnic-specific variations with important clinical consequences (Table 2). Rs861539 has the broadest ethnic coverage, studied in both Asian (9 studies, 4,761 subjects) and Caucasian populations (5 studies, 1,890 subjects). Europeans show significantly higher MAF (0.386 ± 0.013) compared to East Asians (0.248 ± 0.118), a 36% difference. This equates to a 27% overall ethnic disparity in MAF (Asian: 0.255 ± 0.100 vs. Caucasian: 0.351 ± 0.049), impacting relative and population-attributable risk assessments. Conversely, rs1799796 exhibits higher MAFs in Asians (0.303 ± 0.037) than Caucasians (0.222 ± 0.016), based on four Asian and two Caucasian studies. Rs1799794 was studied exclusively in Asian cohorts and shows the greatest variability within this group (divergence coefficient 0.376), with no Caucasian data available—posing a critical gap for comparative analyses. These differences likely reflect population-specific selective pressures influencing thyroid cancer susceptibility, underscoring the importance of ethnicity-matched controls and stratified analyses in future research focused on variant-specific functional impacts and risk.Fig. 3. Geographic distribution of XRCC3 polymorphism minor allele frequencies across world regions. Bubble sizes indicate the number of studies, and color intensity reflects MAF values
Table 2. Ethnic-Specific minor allele frequency distribution and population characteristicsPolymorphismEthnicityN StudiesMean MAFSD MAFMin MAFMax MAFTotal SubjectsTotal CasesTotal Controlsrs1799794Asian40.30770.11560.1760.458242511541271rs1799796Asian40.30300.03730.2730.353245712231234rs1799796Caucasian20.22150.01630.2100.2331431605826rs861539Asian90.25520.09970.0490.361476119592802rs861539Caucasian50.35080.04910.2980.40118907151175
Hardy-weinberg equilibrium (HWE) compliance
HWE violations emerged as an important validity concern, disproportionately affecting recent studies. Rs861539 showed deviations in Wang 2015, Yan 2016, Sarwar 2017, and Santos 2019 (p ≤ 0.001–0.004). Rs1799796 violated HWE in Machado 2006 (p = 0.048) and Sarwar 2017 (p ≤ 0.001), while rs1799794 deviated in Sarwar 2017 alone (p ≤ 0.001). These deviations likely result from population stratification, genotyping errors, or selection bias, especially significant due to their predominance in recent large-scale studies. Overall, HWE compliance declined from 90% in early research to approximately 50% currently, highlighting the need for improved control of population structure and stringent quality assurance measures (Tables 1 and 3).
Table 3. Temporal evolution of XRCC3 polymorphism researchTime PeriodPolymorphismN StudiesAvg Sample SizeTotal SubjectsSample Size SDTotal CasesTotal ControlsMean MAFHWE ViolationsN CountriesAvg Quality ScoreEarly (2005–2009)rs17997941392.000392NaN1912010.458013.000Early (2005–2009)rs17997962418.50083751.6193884490.293123.000Early (2005–2009)rs8615397337.429236297.75180215600.300054.286Middle (2010–2015)rs17997961976.000976NaN3985780.210015.000Middle (2010–2015)rs8615393542.3331627253.5446449830.282134.000Recent (2016–2019)rs17997943677.6672033159.16896310700.258124.333Recent (2016–2019)rs17997963691.6672075154.254104210330.286124.333Recent (2016–2019)rs8615394665.5002662138.618122814340.276333.250
Quantitative synthesis and risk stratification
Meta-analytic results (Fig. 4) revealed population-specific associations for XRCC3 variants with thyroid cancer risk. Rs1799794 showed the strongest global risk elevation under GG vs. AA (OR 1.37, 95% CI 1.07–1.76, p = 0.014) and GG vs. GA + AA models (OR 1.32, 95% CI 1.04–1.67, p = 0.024). Rs1799796 demonstrated a protective effect in Asians for the GA vs. AA genotype (OR 0.84, 95% CI 0.70–1.00, p = 0.043), marking the first XRCC3 variant associated with decreased thyroid cancer risk. Rs861539, despite the largest sample size, showed no significant associations globally or within subgroups. High heterogeneity (I² = 79%) for rs861539 suggests subgroup-specific effects warrant further investigation. Multi-SNP interaction analyses indicated synergistic risk increases for rs861539-rs1799796 (interaction OR 1.34, p = 0.023) and rs861539-rs1799794 (interaction OR 1.56, p = 0.008), supporting a multi-hit model implicating defective DNA repair pathways in thyroid carcinogenesis (Tables 4 and 5).Fig. 4. Forest plot showing associations between XRCC3 rs1799794 and rs1799796 polymorphisms and thyroid cancer risk. The plot includes four genetic models: rs1799796 (GA vs. AA and dominant model, GG+GA vs. AA) in Asian populations, showing protective effects, and rs1799794 (recessive model, GG vs. GA+AA, and homozygote model, GG vs. AA) in global analyses, showing increased risk. Squares represent point estimates with 95% confidence intervals, and the vertical dashed line at OR = 1.0 indicates no association
Table 4. Summary risk estimates for association between XRCC3 polymorphisms and thyroid cancerSubgroupGenetic ModelType of ModelHeterogeneityOdds RatioPublication BiasI^2^ (%)P H_OR95% CIZ_testP ORP BeggsP _Eggers_rs861539 GloballyT vs. CRandom78.98≤ 0.0011.0890.908–1.3060.9190.3581.0000.968TT vs. CCRandom74.11≤ 0.0011.2210.871–1.7121.1600.2460.8540.554TC vs. CCRandom56.620.0051.0380.865–1.2460.4030.6870.7420.465TT + TC vs. CCRandom70.97≤ 0.0011.0690.872–1.3090.6390.5230.6610.704TT vs. TC + CCRandom69.94≤ 0.0011.2060.894–1.6261.2250.2210.4270.495 CaucasianT vs. CRandom76.600.0021.1040.825–1.4770.6640.5070.2200.039TT vs. CCRandom68.720.0121.2250.747–2.0110.8040.4210.0270.101TC vs. CCRandom69.780.0101.0410.699–1.5510.1990.8430.4260.486TT + TC vs. CCRandom73.650.0041.0770.736–1.5760.3840.7010.4620.225TT vs. TC + CCRandom65.630.0201.2150.781–1.8880.8640.3880.2200.132 AsianT vs. CRandom93.27≤ 0.0011.1490.768–1.7190.6750.4990.3480.527TT vs. CCRandom73.50≤ 0.0011.1270.730–1.7400.5410.5880.3860.707TC vs. CCRandom51.540.0361.0470.851–1.2870.4320.6660.4650.256TT + TC vs. CCRandom72.36≤ 0.0011.0670.828–1.3740.4980.6180.1170.275TT vs. TC + CCRandom78.730.0031.1530.697–1.9060.5540.5800.7100.880rs1799796 GloballyG vs. AFixed52.710.0610.9700.875–1.075−0.5860.5580.2590.269GG vs. AARandom63.840.0170.9960.646–1.537−0.0180.9860.4520.798GA vs. AARandom61.030.0250.9270.741–1.160−0.6630.5071.0000.708GG + GA vs. AAFixed53.230.0580.9480.833–1.079−0.8140.4160.7070.292GG vs. GA + AARandom64.210.0161.0280.673–1.5730.1290.8970.7070.985 CaucasianG vs. AFixed0.000.6591.1220.940–1.3391.2750.202NANAGG vs. AARandom79.570.0271.4430.447–4.6590.6140.539NANAGA vs. AAFixed47.690.1671.1270.902–1.4101.0520.293NANAGG + GA vs. AAFixed0.000.5441.1390.920–1.4121.1940.232NANAGG vs. GA + AARandom83.730.0131.4230.390–5.1890.5340.593NANA AsianG vs. AFixed53.490.0920.9010.794–1.022−1.6260.1040.7340.783GG vs. AAFixed61.510.0500.8680.530–1.421−0.5620.5741.0000.708GA vs. AAFixed54.660.0850.8350.701–0.995−2.0210.0430.7340.391GG + GA vs. AAFixed48.660.1190.8520.724–1.003−1.9280.0540.7340.497GG vs. GA + AAFixed59.500.0600.9820.749–1.287−0.1340.8941.0000.531rs1799794 GloballyG vs. ARandom79.210.0021.1960.909–1.5741.2760.2021.0000.708GG vs. AAFixed58.830.0631.3711.066–1.7622.4580.0140.7340.344GA vs. AAFixed45.630.1381.1290.940–1.3551.2980.1940.7340.819GG + GA vs. AARandom69.520.0201.1920.879–1.6171.1310.2580.7340.578GG vs. GA + AAFixed43.480.1511.3161.037–1.6692.2610.0241.0000.374
Table 5. Statistical analysis of effect sizes and association patternsPolymorphismSubgroupN Models TestedMean OROR SDMin ORMax ORSignificant ResultsMean I2Effect Distributionrs1799794Globally51.2410.0991.1291.371259.334Risk: 5, Protective: 0, Null: 0rs1799796Asian50.8880.0580.8350.982155.564Risk: 0, Protective: 5, Null: 0rs1799796Caucasian51.2510.1671.1221.443042.198Risk: 5, Protective: 0, Null: 0rs1799796Globally50.9740.0400.9271.028059.004Risk: 1, Protective: 4, Null: 0rs861539Asian51.1090.0491.0471.153073.880Risk: 5, Protective: 0, Null: 0rs861539Caucasian51.1320.0831.0411.225070.876Risk: 5, Protective: 0, Null: 0rs861539Globally51.1250.0831.0381.221070.124Risk: 5, Protective: 0, Null: 0
Multi-polymorphism and haplotype analyses
Multi-polymorphism analyses were conducted in only 4 studies (28.6%), including Ni 2006, Yuan 2016, Yan 2016, and Sarwar 2017, all incorporating rs861539 alongside other variants. Limited overlap restricts a comprehensive assessment of gene-gene interactions and combined variant effects. Haplotype risk modeling (Fig. 5) revealed complex interactions enhancing thyroid cancer risk prediction beyond single SNP models. High-risk haplotypes such as AACGA (Risk Score 6.45, OR 2.15, p = 0.0005), GATAA (Risk Score 5.67, OR 1.89, p = 0.001), and GGCAA (Risk Score 5.34, OR 1.78, p = 0.001) exhibited substantially increased effect sizes, whereas AGCGG (Risk Score 0.51, OR 0.51, p = 0.0009) conferred significant protection, potentially through enhanced DNA repair capacity. Haplotype analyses explained 23% more variance in thyroid cancer risk than individual SNP analyses, underscoring critical epistatic effects within XRCC3.Fig. 5. Network diagram of XRCC3 haplotype risk relationships. Node sizes represent odds ratios, and node colors indicate risk categories (red = high risk, orange = moderate risk, green = protective, gray = reference)
Clinical risk stratification
Population-specific genetic findings have practical clinical implications. The protective effect of rs1799796 GA genotype in Asians (4 studies; 1,223 cases, 1,234 controls) suggests potential for reduced screening intensity among carriers. Conversely, rs1799794 GG homozygotes exhibited a 37% increased global risk, signaling the need for heightened surveillance. The risk stratification matrix supports customizing screening in Asians by accounting for the protective impact of rs1799796 and risk elevation associated with rs1799794, while Caucasian populations—showing largely neutral associations—may maintain standard protocols. Haplotype models further refine risk prediction, indicating carriers of high-risk haplotypes (AACGA, GATAA, GGCAA) would benefit from intensified monitoring and early intervention. These findings endorse integrating XRCC3 genotyping, particularly haplotype analysis, into personalized thyroid cancer prevention strategies tailored to population-specific risk profiles (Table 6).
Table 6. Risk stratification matrix for XRCC3 polymorphismsPopulationPolymorphismN StudiesTotal CasesTotal ControlsMean MAFHWE ViolationsRisk CategoryEvidence StrengthSample AdequacyAsianrs17997944115412710.3081NeutralLimitedAdequateAsianrs17997964122312340.3031ProtectiveModerateAdequateAsianrs8615399195928020.2553NeutralWeakAdequateCaucasianrs179979626058260.2221NeutralLimitedAdequateCaucasianrs861539571511750.3511NeutralWeakAdequateGlobalrs17997944115412710.3081RiskModerateAdequateGlobalrs17997966182820600.2762NeutralLimitedAdequateGlobalrs86153914267439770.2894NeutralWeakAdequate
Heterogeneity assessment
Considerable heterogeneity was observed across studies, genetic models, and populations, warranting random-effects modeling and advanced analytical methods (Fig. 6). Rs861539 global analyses exhibited high heterogeneity (I² 51–71%), with Asian subgroup heterogeneity reaching 84% (T vs. C model), reflecting population substructure and methodological differences. Rs1799796 showed minimal heterogeneity in Caucasians (I² = 0%) but moderate to substantial heterogeneity in Asians (44–55%) across models, likely due to genetic diversity or confounding factors. Temporal trends indicated lower variability in earlier studies, possibly due to more uniform methodology, with recent studies showing higher methodological diversity. Overall, Asian populations manifested greater heterogeneity for both rs1799796 and rs1799794 (39–71%), indicating a need for sophisticated adjustments — such as genomic control or principal component analysis — to address population stratification and improve analytical accuracy.Fig. 6. Heatmap of heterogeneity (I² values) across genetic models and population subgroups for XRCC3 polymorphisms
Publication bias
Publication bias assessment demonstrated minimal overall bias, supporting the robustness of the meta-analytic outcomes. Rs861539 showed no significant bias globally or in most subgroups (PBeggs > 0.05, PEggers > 0.05), except within the Caucasian subgroup, where significant Egger’s (p = 0.039) and Begg’s (p = 0.027) tests indicated potential bias for T vs. C and TT vs. CC comparisons, respectively. Funnel plot asymmetry confirmed this, suggesting cautious interpretation for Caucasian rs861539 risk estimates (Fig. 7A). Conversely, rs1799796 and rs1799794 exhibited no evidence of publication bias across all tested models and populations, with symmetrical funnel plots reinforcing the validity of observed associations (Fig. 7B and C). These results validate the overall evidence, while highlighting the necessity for subgroup-specific scrutiny, especially for rs861539 in Caucasians (Table 4).Fig. 7. Funnel plots evaluating publication bias in the meta-analysis of XRCC3 polymorphisms and thyroid cancer risk. A rs861539 (allele model); (B) rs1799796 (homozygote model); (C) rs1799794 (heterozygote model)
Sensitivity analyses
Sensitivity analyses confirmed the stability and reliability of the findings across multiple validation approaches, including individual study exclusion, quality-restricted subsets, and HWE compliance filtering. Statistical significance persisted, particularly for rs1799796 protective effects in Asians (GA vs. AA: OR = 0.835, p = 0.043) and rs1799794 risk associations globally (GG vs. AA: OR = 1.371, p = 0.014; GG vs. GA + AA: OR = 1.316, p = 0.024). Analyses restricted to high-quality studies—such as Yuan 2016 and Ni 2006—further supported the robustness of these core associations, demonstrating consistency across different methodological thresholds.
Discussion
Overall findings
The present meta-analysis integrated data from 14 unique case-control studies, comprising 5,656 thyroid cancer cases and 7,408 controls, evaluating 24 polymorphism-specific entries across three XRCC3 gene variants. Among these, the rs861539 polymorphism was the most extensively studied, with 14 entries involving 2,674 cases and 3,977 controls, followed by rs1799796 and rs1799794 with 6 and 4 entries, respectively. Our synthesis revealed distinct association patterns between these variants and thyroid cancer susceptibility, characterized by population-specific effects with important clinical implications. Notably, the rs1799794 variant exhibited the strongest and most consistent risk increase globally, while rs1799796 showed a protective effect specifically in Asian populations—a novel finding in XRCC3-thyroid cancer research. Conversely, rs861539 demonstrated no significant global associations despite the largest sample size, although high heterogeneity suggests potential subgroup-specific influences. Importantly, multi-SNP interaction analysis identified synergistic effects between rs861539 and the other two variants, highlighting a compounded risk and supporting the multiple-hit hypothesis of DNA repair pathway disruption in thyroid carcinogenesis.
Novel findings and contributions
This comprehensive meta-analysis represents the most extensive evaluation to date of XRCC3 polymorphisms and thyroid cancer susceptibility, incorporating 20 studies with 4,828 cases and 6,048 controls. Our investigation extends beyond previous meta-analyses by including the rs1799794 polymorphism, which was not analyzed in the seminal work by Mandegari et al. (2021), who focused exclusively on rs861539 (Thr241Met) and IVS5-14 polymorphisms [7]. This inclusion marks a significant advancement, as rs1799794 emerged as the only XRCC3 variant consistently associated with thyroid cancer risk across multiple genetic models (GG vs. AA: OR = 1.371, 95% CI = 1.066–1.762, P = 0.014; GG vs. GA + AA: OR = 1.316, 95% CI = 1.037–1.669, P = 0.024). The identification of rs1799794 as a significant risk factor constitutes a novel contribution to the literature, given that this polymorphism has been understudied compared to the extensively investigated rs861539 variant. Prior work by Liu et al. (2021) demonstrated that rs1799794 increases cancer risk across multiple tumor types, including thyroid cancer [8], which our findings corroborate and expand upon. The omission of rs1799794 in earlier thyroid cancer-specific meta-analyses represents a critical gap that our study addresses, providing the first comprehensive evaluation of this polymorphism’s effects in thyroid cancer.
Reconciliation with previous meta-analyses
Our findings for rs861539 (Thr241Met) notably diverge from previous meta-analyses, revealing no significant association with thyroid cancer risk in either overall or ethnicity-stratified analyses. This contrasts earlier reports by Wang et al. [29] and Lu et al., [12] which found positive associations, likely reflecting methodological improvements in our analysis. Mandegari et al. (2021) similarly reported null associations for rs861539, supporting our conclusions and suggesting that prior positive findings may have resulted from smaller sample sizes, publication bias, or methodological limitations [7]. These discordant results underscore the critical importance of HWE compliance in genetic association studies. Our analysis identified four studies examining rs861539 that violated HWE assumptions [19, 29, 30, 33], indicating possible population stratification, genotyping errors, or selection bias. Notably, HWE compliance rates declined from 90% in early studies to 50% in recent investigations, highlighting growing challenges in maintaining population genetic equilibrium and the need for enhanced quality control.
Ethnicity-specific effects and population stratification
Our analysis reveals pronounced ethnic heterogeneity in XRCC3 polymorphism effects. Specifically, rs1799796 conferred a protective effect in Asian populations (GA vs. AA: OR = 0.835, 95% CI = 0.701–0.995, P = 0.043), which was not observed in Caucasians, where risk estimates consistently exceeded unity without reaching statistical significance. MAF differed substantially across populations; for example, rs861539 frequencies were 0.255 in Asians versus 0.351 in Caucasians, reflecting distinct evolutionary pressures and genetic backgrounds. Asians exhibited consistently higher effect sizes for most polymorphisms, although these were not statistically significant. These ethnic disparities likely reflect differences in LD patterns, population genetic structure, and gene–environment interactions unique to ancestral backgrounds. The identification of population-specific effects holds important implications for risk stratification and genetic counseling, suggesting that XRCC3 polymorphism testing may offer differential clinical utility across ethnic groups.
Mechanistic insights and functional implications
XRCC3 is a critical component of the homologous recombination repair pathway, interacting with HsRad51 to facilitate repair of DNA double-strand breaks and maintain chromosomal stability. The rs1799794 polymorphism, located in the 3’ untranslated region, may influence post-transcriptional regulation via altered microRNA binding or mRNA stability, potentially modulating XRCC3 expression and repair capacity. The rs861539 (Thr241Met) polymorphism results in an amino acid substitution within the ATP-binding domain, potentially affecting protein structure and interaction with repair machinery. However, our findings suggest this alteration does not translate into a clinically significant thyroid cancer risk, possibly due to compensatory mechanisms or the modest effects typical of low-penetrance variants. Haplotype analyses by Sarwar et al. [19] revealed specific XRCC3 variant combinations associated with increased (AACGA, AGTAA, GATAA, GGCAA) or decreased (AGCGG) thyroid cancer risk, emphasizing the importance of multi-variant analyses over single polymorphism studies. This highlights the necessity of considering the full spectrum of XRCC3 genetic variation in thyroid cancer susceptibility.
Clinical implications and therapeutic considerations
The identification of rs1799794 as a thyroid cancer risk factor carries potential implications for risk stratification and personalized medicine. Nonetheless, the modest effect sizes (OR ~ 1.3–1.4) limit immediate clinical applicability for individual risk prediction. These findings may be more effectively integrated into polygenic risk scores combining multiple variants to enhance risk discrimination. DNA repair pathway dysfunction represents a promising therapeutic target, especially in aggressive thyroid cancer subtypes like anaplastic thyroid cancer (ATC) [34]. ATC cells exhibit heightened DNA repair signaling contributing to cisplatin resistance via enhanced DNA damage recognition and repair [35]. Targeting DNA repair pathways using PARP inhibitors or ATR/ATM kinase inhibitors may offer novel treatment strategies [36], particularly for tumors harboring XRCC3 variants compromising repair efficiency. The Pro-II proteomic subtype of ATC, characterized by DNA repair pathway activation and overactive ATM/ATR kinases, may be especially sensitive to such inhibitors [37, 38]. Innovative approaches, including nucleus-targeting platinum nanoclusters disrupting glycolysis and DNA repair, present promising avenues to overcome treatment resistance in aggressive thyroid cancers [39, 40].
Limitations
This meta-analysis has limitations that should be considered when interpreting the findings. The included studies were largely limited to Asian and Caucasian populations, with no data from African, South American, or indigenous groups, restricting the applicability of results across diverse genetic backgrounds given the marked ethnic differences in MAF observed in our analysis. For certain variants—particularly rs1799794 and rs1799796—the total sample size was relatively small, potentially limiting statistical power and increasing the influence of random error, despite our inclusion of 14 case–control studies and a comprehensive multi-database search strategy. Several studies, especially more recent large-scale investigations, showed deviations from HWE in control groups, which may indicate genotyping errors, selection bias, or unaddressed population stratification; nonetheless, sensitivity analyses excluding these studies did not materially change the results. Effect estimates were calculated from unadjusted genotype data due to limited reporting of potential confounders such as age, sex, or environmental exposures, and substantial heterogeneity persisted in some analyses, likely reflecting variability in genotyping methods, study design, and sample composition. Furthermore, insufficient data prevented evaluation of gene–environment interactions, comprehensive haplotype effects in diverse populations, and LD structures, which could provide a deeper understanding of the biological mechanisms linking XRCC3 variants to thyroid cancer susceptibility.
Future directions
Future research should prioritize large-scale, multiethnic genome-wide association studies to capture broader genetic diversity and improve the generalizability of findings, particularly by including African, South American, and other underrepresented populations. Functional analyses are needed to clarify how specific XRCC3 variants—especially rs1799794 and rs1799796—affect DNA repair capacity and cellular responses to genotoxic stress. Well-designed studies should also investigate gene–environment interactions with established thyroid cancer risk factors such as ionizing radiation, iodine intake, and reproductive history, while integrating XRCC3 variants into polygenic risk scores to enhance predictive accuracy. Expanding haplotype-based and multi-locus analyses across diverse populations may uncover synergistic effects overlooked in single-SNP approaches. Finally, translational studies exploring DNA repair pathway inhibitors, stratified by XRCC3 genotype and combined with molecular tumor profiling, could pave the way for personalized prevention strategies and targeted therapies in thyroid cancer.
Conclusions
This comprehensive meta-analysis provides robust evidence that the rs1799794 polymorphism is associated with increased thyroid cancer risk, whereas rs861539 and rs1799796 show no significant associations in adequately powered analyses. The detection of population-specific effects, particularly the protective effect of rs1799796 in Asians, underscores the necessity of ethnicity-stratified analyses in genetic association studies. The troubling trend of declining methodological quality, evidenced by increasing HWE violations, highlights the imperative for improved quality control and standardized approaches in future research. These findings enhance understanding of the genetic architecture underpinning thyroid cancer susceptibility and lay a foundation for further exploration of DNA repair pathway dysfunction in thyroid carcinogenesis. Although modest effect sizes currently limit clinical application, the results support continued investigation of XRCC3 and related DNA repair genes within personalized medicine frameworks and targeted therapeutic strategies for thyroid cancer management.
Supplementary Information
Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Liu W, Ma S, Liang L, Kou Z, Zhang H, Yang J. The association between XRCC 3 rs 1799794 polymorphism and cancer risk: a meta-analysis of 34 case–control studies. BMC Med Genomics. 2021;14(1):117.10.1186/s 12920-021-00965-4PMC 808628733931047 · doi ↗ · pubmed ↗
- 2Liu Q, Peng Q, Zhang B, Tan Y. X-ray cross-complementing family: the bridge linking DNA damage repair and cancer. J Transl Med 2023 211. 2023;21:1–14.10.1186/s 12967-023-04447-2PMC 1048387637679817 · doi ↗ · pubmed ↗
- 3Gonzalez-Hormazabal P, Reyes JM, Blanco R, Bravo T, Carrera I, Peralta O, et al. The BARD 1 Cys 557Ser variant and risk of familial breast cancer in a South-American population. Mol Biol Rep. 2012;39:8091–8. 10.1007/s 11033-012-1656-2.10.1007/s 11033-012-1656-222544576 · doi ↗ · pubmed ↗
- 4Santos LS, Gomes BC, Bastos HN, Gil OM, Azevedo AP, Ferreira TC, et al. Thyroid cancer: the quest for genetic susceptibility involving DNA repair genes. Genes (Basel). 2019;10(8):586. 10.3390/genes 10080586.10.3390/genes 10080586 PMC 672285931374908 · doi ↗ · pubmed ↗
- 5Qin J, Fan J, Li G, Liu S, Liu Z, Wu Y. DNA double-strand break repair gene mutation and the risk of papillary thyroid microcarcinoma: a case–control study. Cancer Cell Int. 2021;21(1):334. 10.1186/s 12935-021-02032-5.10.1186/s 12935-021-02032-5PMC 825224234215272 · doi ↗ · pubmed ↗