Adsorption of phage T2 is inhibited due to inversion of cryptic prophage DNA by the serine recombinase PinQ
Joy Kirigo, Daniel Huelgas‐Méndez, María Tomás, Michael J Benedik, Rodolfo García‐Contreras, Thomas K Wood

TL;DR
A recombinase enzyme in E. coli cryptic prophage inhibits T2 phage infection by inverting DNA and blocking phage adsorption.
Contribution
Discovery of the first recombinase-activated phage inhibition system involving DNA inversion and receptor blocking.
Findings
PinQ serine recombinase inverts a DNA segment to form a novel protein StfE2 that blocks phage adsorption.
StfE2 prevents T2 phage Gp38 from binding to porins FadL and OmpF.
T2 phage escape mutants develop resistance by mutating the Gp38 hypervariable region.
Abstract
Recombinases catalyze site-specific integration, excision, and inversion of DNA and are often found adjacent to anti-phage system genes clustered in defense islands; however, their function in phage defense is unknown, as they are frequently dismissed as markers of prophages. Here, we characterize the physiological role of the previously uncharacterized serine recombinase PinQ (P segment inversion by Qin) of Escherichia coli cryptic prophage Qin and discover that it inhibits T2 phage infection by inverting a 1797 bp segment in a different cryptic prophage e14; this inversion leads to the formation of a novel protein from two chimeric genes, StfE2, that we find blocks phage adsorption. Modeling shows StfE2 inhibits T2 phage adsorption by preventing Gp38 binding to its primary receptors, porins FadL and OmpF. Corroborating the receptor-blocking hypothesis, T2 escape mutants evolve…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
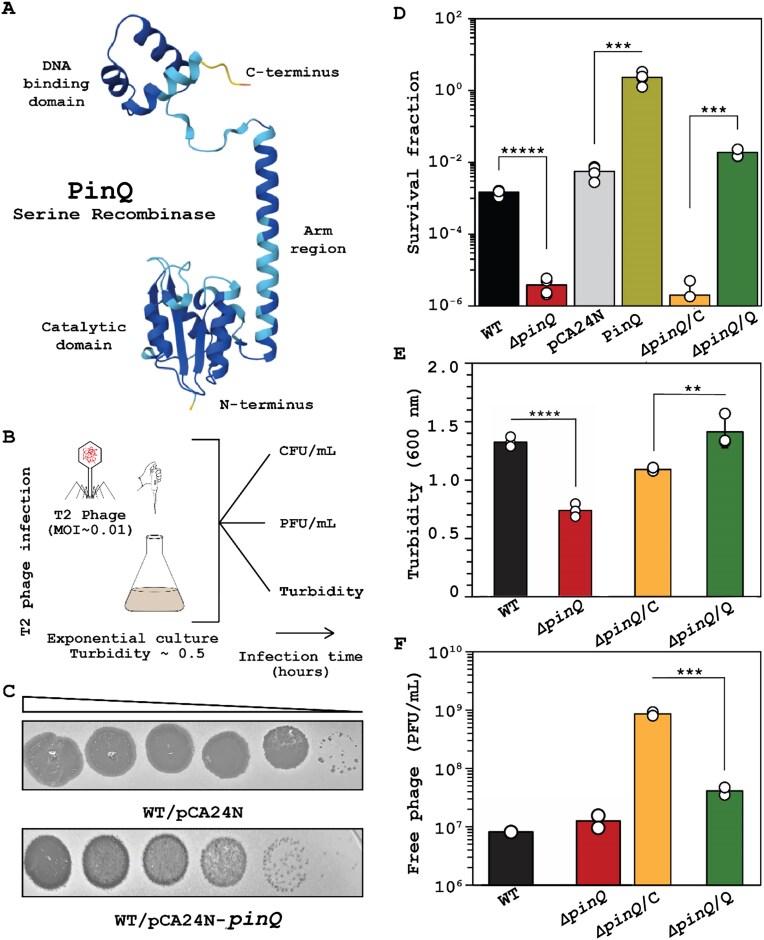 Figure 1
Figure 1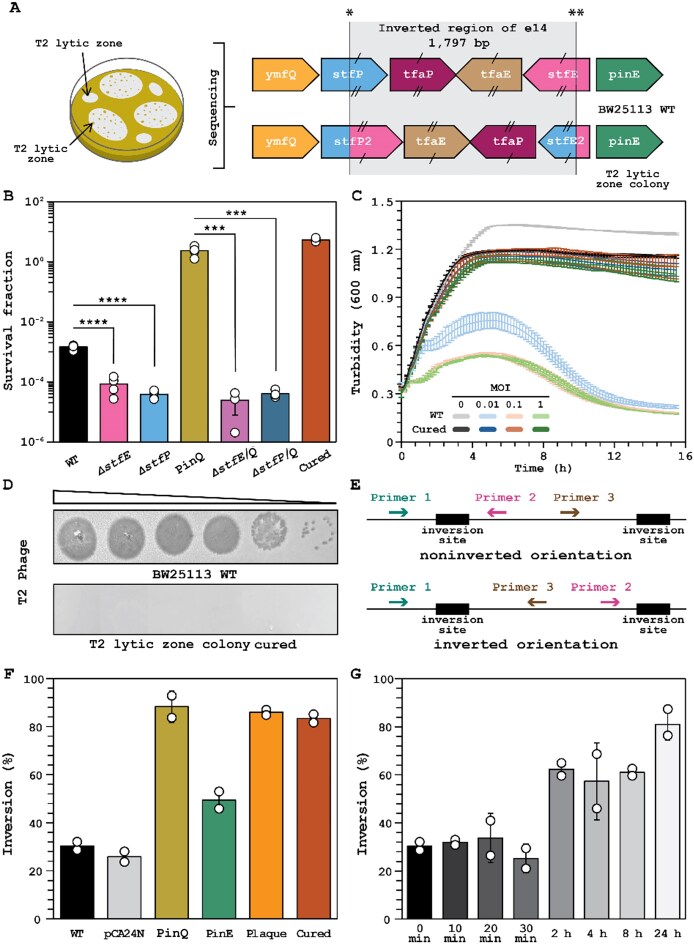 Figure 2
Figure 2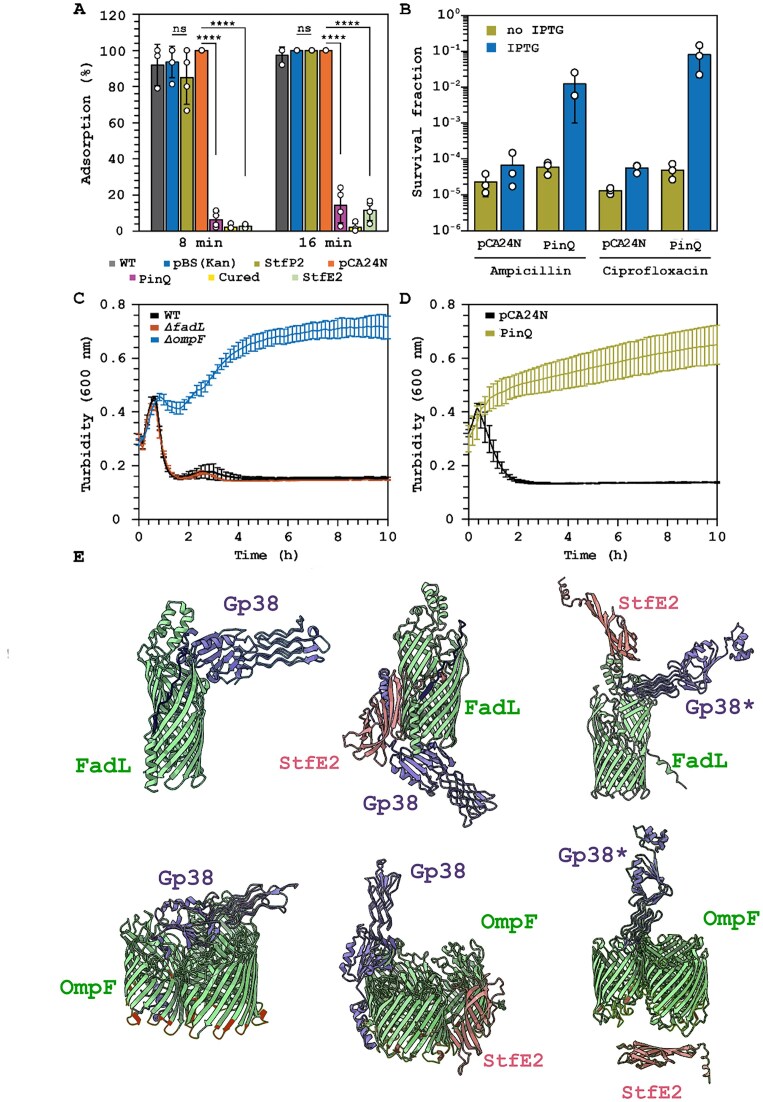 Figure 3
Figure 3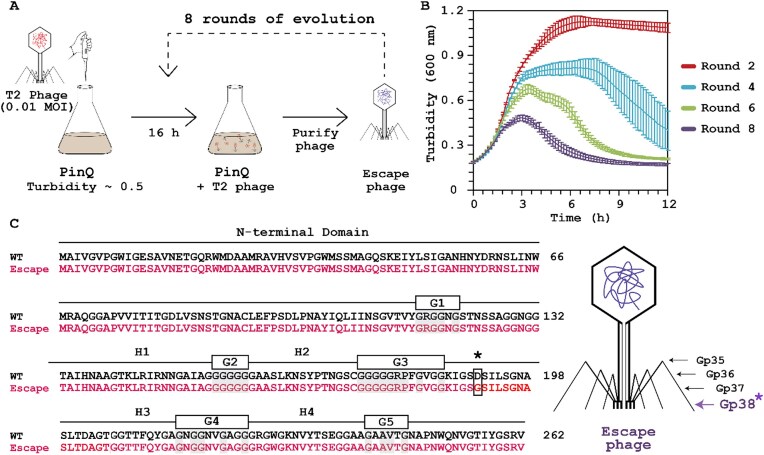 Figure 4
Figure 4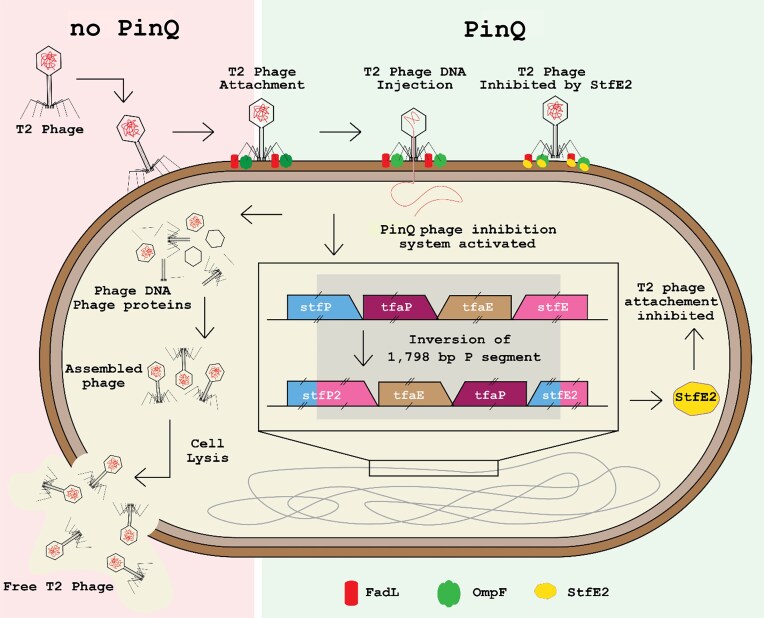 Figure 5
Figure 5| Strains/Plasmids/Phages | Features | Source |
|---|---|---|
| Strains | ||
| BW25113 |
| [ |
| BW25113 PinQ lytic zone colony 1 | pCA24N- | This study |
| BW25113 PinQ lytic zone colony 2 | pCA24N- | This study |
| BW25113 cured | BW25113 PinQ lytic zone colony 1 with pCA24N- | This study |
| BW25113 Δ |
| [ |
| BW25113 Δ |
| [ |
| BW25113 Δ |
| [ |
| BW25113 Δ |
| [ |
| BW25113 Δ | KanS | This study |
| BW25113 Δ | KanS | This study |
| BW25113 Δ |
| [ |
| BW25113 Δ |
| [ |
| BW25113 Δ |
| [ |
| BW25113 Δ |
| [ |
| BW25113 Δ | KanS | This study |
| BW25113 Δe14 | KanS | [ |
| Plasmids | ||
| pCA24N | CmR; | [ |
| pCA24N- | CmR; | [ |
| pCA24N- | CmR; | [ |
| pCA24N- | CmR; | [ |
| pCA24N- | CmR; | [ |
| pCA24N- | CmR; | This study |
| pCP20 | ApR and CmR, FLP recombinase, temperature-sensitive replication | [ |
| pBS(Kan) | KanR; | [32] |
| pBS(Kan)- | KanR; | This study |
| Phages | ||
| T2 | TKW stock | |
| T4 | TKW stock | |
| P1 | TKW stock | |
| Bas03 |
| [ |
| Bas25 |
| [ |
| Bas26 |
| [ |
| Bas66 |
| [ |
| Bas69 |
| [ |
- —Biotechnology Endowment
- —DGAPA10.13039/501100006087
- —UNAM10.13039/501100005739
- —SECIHTI
- —Pennsylvania State University10.13039/100008321
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Bacterial Genetics and Biotechnology · RNA and protein synthesis mechanisms
Introduction
The symbiotic relationship between prophages (integrated viral genomes) and their hosts is longstanding and widespread [1]. The majority of all sequenced bacterial genomes contain one or more prophage sequences [1, 2], and viral sequences are present not only in bacterial genomes [3] but also in plant [4], animal, and human genomes [5]. In bacteria, active prophages provide reservoirs for virulence genes (e.g. Shiga toxin, botulinum toxin, diphtheria toxin) [6] and host immunity against phages (e.g. restriction systems) [7].
In contrast, evidence is mounting that cryptic prophages; i.e. prophage fossils that are no longer able to lyse the cell or make phage particles [8], play a role in bacterial physiology. For example, Escherichia coli K-12 contains nine cryptic prophages (CP4-6, DLP12, e14, rac, Qin, CP4-44, CPS-53, CPZ-55, and CP4-57) [9], and they have been shown to influence its stress response to antibiotics and oxidative stress [10, 11] as well as regulate resuscitation from stress-induced dormancy [12]. Moreover, like prophages, cryptic prophages are involved in anti-phage defense in that e14 prophage encodes phage inhibition systems Lit (inhibits T4 replication) [13] and McrA (degrades T-even phage DNA) [14], and CP4-6, rac, Qin, CP4-44, and CP4-57 contain toxin-antitoxin systems [9], whose primary physiological role is phage inhibition [15, 16].
Cryptic prophages are also hotbeds for DNA-modifying enzymes, with site-specific recombinases (integrases) that may be active and are critical for innovations in biotechnology [17] and synthetic biology [18]. CP4-6, CP4-57, CPS-53, CPZ-55, DLP12, e14, rac, and Qin prophages all contain recombinases [9], which catalyze the integration, excision, or inversion of defined DNA segments [19]. These recombinases fall into two main families, serine recombinases and tyrosine recombinases, named in reference to the conserved amino acid residue that mediates catalysis [19]. In addition, serine recombinases fall into two classes: large serine recombinases, responsible for excision, integration, deletion, or transposition; and small serine recombinases (SSRs) that function as invertases or resolvases [20].
Serine recombinases also play an important role in host and phage physiology. For example, SSR Hin catalyzes DNA inversion of a 1 kb segment that controls flagella phage variation in Salmonella typhimurium [21], increasing bacterial virulence [22]. Gin and Cin (SSRs) catalyze inversions of the 3 kb (G segment) and 4.2 kb (C segment) of phages Mu [23] and P1 [24], respectively. Inversion in both the G and C segments enables the phages to adsorb to different bacterial hosts [23, 25]. Escherichia coli K-12 also contains an SSR in cryptic prophage e14, PinE, which inverts a 1.8 kb segment containing putative tail fiber genes; however, the function of this inversion is not clear [26, 27].
Previously, while determining that some E. coli cells that survive phage infection are dormant, we found point mutations arise in pinR and pinQ, which encode uncharacterized putative SSRs, and found deleting pinQ increases the sensitivity to T2 phage 330-fold [28]. Hence, we sought to characterize the physiological role of PinQ here (note PinR is 99% identical, so we focused on PinQ as PinR is equivalent); PinQ, in cryptic prophage Qin, was named due to 38% identity to SSR PinE, but PinQ (and PinR) have not been characterized to date. Critically, this is the first report of a recombinase playing a role in anti-phage defense even though recombinases are commonly positioned near clusters of anti-phage system genes, but they have been dismissed [29]. We found that upon T2 infection, PinQ from cryptic prophage rac inverts a 1.8 kb segment in cryptic prophage e14; this inversion inhibits T2 phage by creating a new, chimeric protein (StfE2) encoded by the upstream portion of stfE^+^ and from the complementary strand of stfP^+^ after the inversion. StfE2 is found to reduce adsorption of phage T2, and modeling shows StfE2 likely inhibits T2 by blocking access to its E. coli receptor, FadL. Moreover, T2 escapes the PinQ-mediated inversion phenotype through mutation of gp38, which encodes its adhesion protein. Therefore, we discovered a new class of phage inhibition system based on DNA recombination and a chimeric protein.
Materials and methods
Bacterial strains, plasmids, phages, medium, and antibiotics
The bacterial strains (E. coli BW25113 [30] and its isogenic mutants), plasmids, and phages used in this work are described in Table 1. Cells were grown in lysogeny broth (LB, 1% tryptone, 0.5% yeast, and 1% NaCl w/v) at 37°C. Kanamycin, 50 μg/ml (Kan50), was used for preculturing knockout mutants and pBS(Kan)-based plasmids, and chloramphenicol, 30 μg/ml (Cm30), was added to strains containing pCA24N-based plasmids to maintain the vector. Single-gene knockouts of E. coli were obtained from the Keio Collection [30]. The pinQ pinE Kan^S^ was constructed using P1 transduction to transfer ΔpinQ Kan^R^ to pinE Kan^S^ [32]; kanamycin resistance was removed using FLP recombinase via pCP20 [32]. The final constructs were verified by polymerase chain reaction (PCR) (primers shown in Supplementary Table S1).
Plasmids used in this study, except for pCA24N-stfE2 and pBS(Kan)-stfP2, were obtained from the ASKA collection [31], and gene expression was induced by the addition of isopropyl β-d-1-thiogalactopyranoside (IPTG, 1 mM). pCA24N-pinQ was cured from BW25113 cells isolated from T2 lytic zones (“cured” strain) by growing in overnight LB liquid cultures with IPTG in the absence of antibiotic for the plasmid and by screening using LB and LB chloramphenicol plates. The absence of the plasmid was confirmed via PCR using pCA24N-specific primers (pCA24N_Fow and pCA24N_Rev, Supplementary Table S1). Phage lysates were stored at 4°C, and T2 phage was used at a multiplicity of infection (MOI) of ∼0.01 for all experiments unless stated otherwise.
pCA24N-stfE2 was constructed by amplifying a 422 bp fragment from the inverted region of the chromosome of PinQ-producing cells (BW25113/pCA24N-pinQ) using primers StfE2_Fow and StfE2_Rev (Supplementary Table S1) to introduce restriction sites for StuI and PstI [31]. pBS(Kan)-stfP2 was constructed by amplifying an 833 bp fragment from the inverted region of the chromosome of PinQ-producing cells (BW25113/pCA24N-pinQ) using primers StfP2_Fow and StfP2_Rev (Supplementary Table S1) to introduce restriction sites for ApaI and BamHI to facilitate cloning into pBS(Kan) [34]; pBS(Kan) was used since it has better promoter silencing with glucose than pCA24N, and construction in pCA24N led only to mutations in stfP2. Constructed plasmid sequences were verified by Plasmidsaurus.
Temporal turbidity and cell viability
T2 phage was added to exponentially growing cells (turbidity at 600 nm ∼0.5; Eppendorf Biophotometer D30), and the cells were incubated at 37°C and 250 rpm and monitored using a UV-Vis spectrometer (Sunrise Tecan). For cell viability (Supplementary Fig. S1 for deletion mutants), 1 ml aliquots were washed twice with phosphate-buffered saline (PBS, 8 g NaCl, 0.2 g KCl, 1.15 g Na_2_HPO_4_, and 0.2 g KH_2_PO_4_ in 1000 ml ddH_2_O), and enumerated using the drop assay [35].
Phage titers and plaque assay
For phage titers, 1 ml samples were centrifuged (10 min, 5000 rpm) to separate free phage from cells, and 100 μl samples of the supernatant (containing free phage) were serially diluted in phage buffer (0.1 M NaCl, 10 mM MgSO_4_, and 20 mM Tris–HCl pH 7.5). Plaques formed (PFU/ml) were enumerated on double-layer agar plates (1% tryptone, 0.5% NaCl, 1.5% agar lower layer, and 0.4% agar top layer) using the drop assay.
Sequencing genomes of cells producing PinQ that survive T2 infection
To gain insights into the mechanism of PinQ phage inhibition, PinQ was produced for 16 h via BW25113/pCA24N-pinQ. To double-layer agar plates, 100 μl of each overnight culture and 100 μl drop of T2 phage stock (6 × 10^8^ PFU/ml) were added and incubated overnight. Surviving colonies inside the lytic zones were purified from phages by streaking on LB Cm30 plates. Genomic DNA was extracted using the Qiagen DNeasy Ultraclean Microbial Kit and then sequenced using Oxford Nanopore Technology (library prep using Ligation Sequencing Kit V14 and sequencing via PromethION R10.4.1 flow cells), and the genome was assembled using Flye v2.9.1 by Plasmidsaurus after removing the sequence reads of T2 DNA. Accession numbers for all the sequences are shown in Supplementary Table S2.
DNA inversion via quantitative PCR
Quantitative PCR (qPCR) was used to quantify the amount of inverted genomic DNA in overnight cultures containing pCA24N-derived plasmids that were induced with IPTG. Genomic DNA was extracted using the Qiagen DNeasy Ultraclean Microbial Kit. qPCR was performed via an Applied Biosystems StepOne real-time PCR system. Primers (Supplementary Table S1) were used to amplify a noninverted 153 bp (PinQ_1 and PinQ_2) or noninverted 831 bp (PinQ_1B and PinQ_2B) or inverted 494 bp (PinQ_1 and PinQ_3) or inverted 586 bp (PinQ_1B and PinQ_3B) segments of the E. coli chromosome of the e14 cryptic prophage (coordinates: 1 196 443–1 210 635) using the Luna Universal qPCR Master Mix. No housekeeping primers are required since inversion compares the ratio of expression of inverted versus noninverted in the same DNA sample.
Quantitative real-time reverse transcriptase polymerase chain reaction
Quantitative real-time reverse transcriptase PCR (qRT-PCR) was used to quantify gene expression after T2 phage infection (5 min, MOI 0.01). RNA was isolated by rapid cooling using ethanol/dry ice in the presence of RNAlater™ stabilization solution (Invitrogen™) and extracted using PureLink™ RNA Mini Kit (Invitrogen™). qRT-PCR was performed using iTaq Universal SYBR^®^ Green RNA-to-CT™ 1-Step kit (Bio-Rad) in an Applied Biosystems StepOne real-time PCR system. Relative quantification was calculated using double delta Ct method with rrsG, 16S ribosomal RNA, as the housekeeping gene. The primers used are shown in Supplementary Table S1.
T2 adsorption
Phage adsorption [36] was assayed by diluting overnight cultures 1:100 into 10 ml of LB (or LB Cm30 for pCA24N-based strains) and incubating until turbidity (600 nm) ∼0.5, then T2 phage was added to the cultures, which were incubated at room temperature without shaking. Samples (1 ml) were taken after 0, 8, and 16 min, centrifuged for 10 min at 5000 rpm, rinsed twice with PBS, and vortexed for 15 s at maximum (Genie 2, Fisher Scientific) to remove free and loosely attached phages. The cells were resuspended in phage buffer (0.1 M NaCl, 10 mM MgSO_4_, and 20 mM Tris–HCl pH 7.5), and PFU were determined using double-layer agar plates.
Antibiotic import through FadL and OmpF
To determine the extent of antibiotic import through the FadL and OmpF pores after production of PinQ, ampicillin (100 μg/ml, 10 minimum inhibitory concentration, MIC) or ciprofloxacin (5 μg/ml, 10 MIC) were added to exponentially growing cells after 2 h production of PinQ via pCA24N-pinQ, and the cells were cultured for 3 h. After treatment, cultures were washed twice with PBS, and cell viability (colony forming units, CFU/ml) was determined using the drop assay. Temporal turbidity (600 nm) measurements were also determined over 10 h every 10 min using a Tecan Sunrise UV-Vis spectrometer for overnight cultures diluted 1:100 into fresh LB in 96-well plates that were incubated at 37°C with shaking (250 rpm) until the exponential growth phase (turbidity ∼0.3), followed by the addition of ampicillin at varying concentrations (0, 5, 10, 20, and 50 μg/ml).
Molecular docking
Protenix [37] was used to predict protein–protein interactions. Structures for protein docking were predicted with AlphaFold3 [38]. 3D structures of Gp38, FadL, and OmpF were downloaded from the Protein Data Bank.
Escape phage mutants
To gain further insights into the mechanism of PinQ phage inhibition, T2 escape mutants were generated and sequenced. Escape mutant populations were generated by sequentially propagating T2 phage on the same E. coli host so that only the phage undergoes mutation. Overnight cultures of BW25113/pCA24N-pinQ were cultured in LB Cm30 to the exponential phase (turbidity at 600 nm ∼0.5), and after 2 h induction with IPTG, T2 phage was added, and cells were incubated for 16 h in 25 ml in 250 ml shake flasks at 37°C while shaking at 250 rpm. After each batch culture, phages were separated from bacterial debris via centrifugation (10 min at 5000 rpm), and the supernatants were filtered (0.22 μm filter). The filtered phage lysate was used to infect a fresh culture of BW25113/pCA24N-pinQ cells for 8 cycles. To determine the extent of T2 phage resistance to PinQ-activated inhibition after each cycle, the susceptibility of exponentially growing (turbidity ∼0.5 at 600 nm), PinQ-producing cells in 96-well plates (after 2 h induction with IPTG) was monitored in the presence of phage isolated from each batch culture for 12 h by scanning every 10 min with a Tecan Sunrise UV-Vis spectrometer.
Phages from the 8th sequential batch culture were sequenced after removing host DNA (via TURBO DNase, Invitrogen) and removing host RNA (via RNase A, Omega Bio-Tek). The DNase was inactivated by heat treating at 75°C for 15 min, and the phage capsid was digested with proteinase K (New England Biolabs). Phage DNA was isolated using the Norgen Biotek Phage DNA Isolation Kit. The presence of T2 phage DNA was confirmed using primers gp28_Fow and gp28_Rev for gp028 (Supplementary Table S1), and the absence of chromosomal E. coli DNA was confirmed using the rrsG gene (rrsG_Fow and rrsG_Rev, Supplementary Table S1).
The escape T2 phage DNA was sequenced using next-generation sequencing with Illumina 2 × 150 bp configuration by Genewiz. The quality checking of the raw reads from the whole genome sequencing was done with FastQC version 0.12.1. The trimming of the first 10 bp in both raw reads was done with Cutadapt version 5.1. The genome assembly was via the BV-BRC server (https://www.bv-brc.org/) [39] using metaSPAdes version 4.0.0 with Read processing set as normalized Illumina reads, Genome parameters set with an estimated genome size of 10 Mb (the assembled contigs had the genome size of phage T2 for both the wild type (WT) and the escape mutant genomes), target genome coverage set at 200, Assembly Polishing utilized with Racon iterations set at 2, Pilon iterations set at 2, Assembly thresholds used with a minimum contig length of 300, and minimum contig coverage set to 5. BLASTn was used to corroborate T2 phage identity of the assembled genome, and the genome was annotated using Pharokka version 1.7.5.
The gp38 gene was sequenced by the Sanger method from a PCR reaction using gp38-f and gp38-r primers (Supplementary Table S1) by Quintara Biosciences. GenBank files of sequenced phages were deposited under the accession codes listed in Supplementary Table S2.
Fitness assay based on growth yield in minimal medium
Growth yields were evaluated in M9 minimal medium [40] with either 0.25% palmitic acid or 0.25% glucose as the sole carbon source. Cultures were incubated for 24 h at 37°C with shaking at 250 rpm, and the initial and final CFU were determined. Experiments were performed with three independent cultures per strain for each carbon source.
Statistical analysis
Statistical analysis was performed using GraphPad Prism. All data presented are the mean ± one standard deviation, and a Student’s t-test was used to evaluate the difference between data sets (probability values (p) < 0.01 were considered significant).
Results
PinQ inhibits T2 phage infection
After finding the pinQ deletion makes E. coli more sensitive to T2 infection [28], we sought to explore the extent to which uncharacterized PinQ inhibits T2 infection. Exploring structural insights using AlphaFold [41], we found PinQ has all the structural features of an SSR [42], including a DNA binding site, arm region, and catalytic domain (Fig. 1A).
PinQ recombinase defends against T2 infection. (A) PinQ is a putative SSR based on its predicted structure. (B) Experimental setup for investigating T2 phage inhibition (0.01 MOI) via plaque, CFU, turbidity, and PFU assays. (C) Plaque assay with cells producing PinQ from BW25113/pCA24N-pinQ. Ten-fold serial dilution of T2 phage. The images shown are for one representative of two independent cultures. (D) Cell survival (full temporal data in Supplementary Fig. S2A for WT and ΔpinQ), turbidity (E, full temporal data in Supplementary Fig. S2B), and T2 free phage (F, full temporal data in Supplementary Fig. S2C) with T2 (0.01 MOI) after 1 h. Bars and error bars are the mean and standard deviation of four independent cultures. Dots are individual data points. *** P< .005, **** P< .0005, ***** P< .00005. Note: WT is E. coli BW25113, pCA24N is WT/pCA24N, PinQ is WT/pCA24N-pinQ, ΔpinQ/C is ΔpinQ/pCA24N, and ΔpinQ/Q is ΔpinQ/pCA24N-pinQ. IPTG was used at 1 mM for producing PinQ from pCA24N-pinQ in panels (C–F).
Next, we directly investigated the role of PinQ on T2 phage inhibition by assaying plaque formation, host survival, temporal cell viability, temporal cell turbidity, and temporal phage production (experimental design shown in Fig. 1B). We found producing PinQ from a plasmid reduces T2 plaque formation by ∼10-fold compared to the empty plasmid (Fig. 1C). Corroborating this result, in the absence of PinQ (ΔpinQ), there was a 400-fold decrease in cell survival after 1 h of infection with T2 phage treatment relative to the WT (Fig. 1D). We used 0.01 MOI for all experiments to avoid premature cell population collapse. This phenotype was completely complemented since production of PinQ (Δ*pinQ/*pCA24N-pinQ) increases cell survival (400-fold increase) relative to the empty plasmid (Fig. 1D).
In agreement with the increase in plaques and reduced survival for the ΔpinQ mutant, temporal cell viability over 5 h without PinQ in the presence of T2 is significantly reduced (100-fold, Supplementary Fig. S2A). Without PinQ, there is also a significant decrease in turbidity during T2 infection (Fig. 1E; Supplementary Fig. S2B); this phenotype was also complemented since producing PinQ in the pinQ mutant leads to no observed decrease in turbidity during T2 treatment (Fig. 1E; Supplementary Fig. S2B). As expected, temporal free phage production in the pinQ mutant is consistently 10-fold higher compared to WT strain over 5 h (Fig. 1F; Supplementary Fig. S2C), and producing PinQ in the pinQ mutant led to a 20-fold decrease in free phage, complementing the phenotype. Together, these six sets of phage infection studies conclusively demonstrate that activation of PinQ inhibits T2 infection.
PinQ inhibits T2 phage infection by inverting the P segment of e14 cryptic prophage
To determine the mechanism of PinQ-related T2 anti-phage defense, we hypothesized that since producing PinQ during T2 infection inhibited T2 phage infection (Fig. 1C–F), producing PinQ, a putative serine recombinase, must lead to genetic changes that may be discerned by sequencing surviving/growing cells from inside T2 lytic zones formed on a lawn of PinQ-producing cells (Fig. 2A). Therefore, whole genome sequencing was performed for surviving cells from two separate colonies in T2 lytic zones. Strikingly, we found an inversion of a 1797 bp of cryptic prophage e14 known as the P segment [43] for both sets of surviving cells (Fig. 2A; Supplementary Table S3). As expected for an SSR, this 1797 bp fragment is flanked by the two inverted repeats required for DNA inversion [44] (5′-TTGGTTTGGGAGAAGG-3′ and 5′-CCTTCTCCCAAA CCAA-3′) (Supplementary Fig. S3). This inversion occurs within the genes stfP^+^ and stfE^+^ in e14 (Fig. 2A; Supplementary Fig. S3); note these two genes are in the opposite orientation, so the inversion, which occurs within the coding region of both, generates two complete chimeric genes that we call stfP2^+^ (begins with stfP^+^ and ends with stfE^+^) and stfE2^+^ (begins with stfE^+^ and ends with stfP^+^), encoding chimeric proteins StfP2 and StfE2 (Supplementary Fig. S4). Hence, cells that survive T2 infection have an inversion of the 1797 bp P segment in e14.
PinQ inhibits T2 phage infection by inverting the e14 P segment. (A) LHS: schematic of the experiment for isolating colonies within T2 lytic zones. RHS: sequencing of the lytic zone survivors revealed the inverted 1797 bp region (P segment) of e14 cryptic prophage, which produces chimeric genes stfP2+ and stfE2+. (B) Cell survival after contact with T2 phage (0.01 MOI) for 1 h. (C) Turbidity with T2 (0, 0.01, 0.1, and 1 MOI). Data points are the mean and standard deviation of three independent cultures. (D) Plaque assay. Ten-fold serial dilution of T2 phage WT. Images shown are for one representative of two independent cultures. (E) qPCR experiment schematic for determining the extent of inversion. (F) Inversion of DNA isolated from overnight cultures based on qPCR (cf., PCR results in Supplementary Fig. S6). Bars and error bars are the mean and standard deviation of two independent cultures with two replicates for each. (G) Inversion of WT during T2 (0.01 MOI) phage infection based on qPCR. Bars and error bars are the mean and standard deviation of four independent cultures. *** P< .005, **** P< .0005. Dots are individual data points. Extent of inversion was determined using primer pair PinQ_1–PinQ_2 (noninverted) and PinQ_1–PinQ_3 (inverted) (Supplementary Table S1). Note: WT is E. coli BW25113, pCA24N is WT/pCA24N, PinQ is WT/pCA24N-pinQ, PinE is WT/pCA24N-pinE, Lytic zone is the PinQ-producing cells isolated from the T2 lytic zone, and cured are cells derived from the PinQ lytic zone colony cured of pCA24N-pinQ. Inversion occurs at the inverted repeat sites * (5′-TTGGTTTGGGAGAAGG-3′) and ** (5′-CCTTCTCCCAAACCAA-3′). IPTG was used at 1 mM for producing PinQ from pCA24N-pinQ in panels (B) and (F).
To verify that the sequenced cells from colonies that formed in the T2 lytic zone are resistant to T2 phage, host survival, temporal turbidity, and phage production were assayed. We found the cured strain (lytic zone colony cured of the pCA24N-pinQ vector) had a 3600-fold increase in cell survival relative to WT (Fig. 2B). In agreement with this result, the cured strain showed no collapse in growth in the presence of T2 even at 1 MOI (Fig. 2C). Strikingly, unlike the WT, no T2 plaques were seen at any T2 phage concentration for the cured strain (Fig. 2D). Hence, the cells that survived with the DNA inversion in e14 cryptic prophage are resistant to T2 infection. Corroborating the importance of the inversion, we found that preventing inversion either by deleting stfP or deleting stfE, since each mutation removes one of the inverted repeats, there was a 40- and 20-fold decrease in cell survival, respectively, after 1 h of infection with T2 phage relative to the WT (Fig. 2B). Hence, the inversion is necessary for anti-phage defense.
Similarly, PinQ production in the stfP and stfE mutants (ΔstfP/pCA24N-pinQ and ΔstfE/pCA24N-pinQ) had no impact relative to the stfP and stfE mutants alone, and these stfP and stfE deletions in the presence of PinQ heterologous expression show a 60 000- and 90 000-fold decrease in survival, respectively, relative to WT with PinQ expression (BW25113/pCA24N-pinQ) (Fig. 2B). Hence, PinQ is effective only when inversion is possible. As an additional control, we found that producing PinQ in the delta e14 strain (that lacks the P segment) also has no effect on T2 inhibition (Supplementary Fig. S5).
To confirm PinQ inverts the e14 DNA and to quantify the extent of inversion, qPCR was performed using a three-primer design shown in Fig. 2E. We found the inversion was 30 ± 2% in the WT and that the empty plasmid pCA24N does not significantly affect inversion (26 ± 3%) (Fig. 2F; Supplementary Table S4); these inversion rates were confirmed using PCR (Supplementary Fig. S6A) that similarly demonstrated in a WT culture the inversion loop is primarily in one orientation, with only a small fraction being inverted. As expected, inactivation of PinQ reduces inversion (Supplementary Table S4). In contrast, producing PinQ leads to e14 DNA inversion (88 ± 6%) (Fig. 2F; Supplementary Fig. S6; Supplementary Table S4) and most cells in the population are in the inverted state. PinE, an SSR that is part of cryptic prophage e14 that inverts this same segment of DNA and has 38% identity with PinQ, led to 49 ± 5% inversion (Fig. 2F; Supplementary Fig. S6; Supplementary Table S4); like PinQ, the physiological role of PinE is unknown [43]. Moreover, sequenced cells that survived T2 infection were isolated from lytic zones and were producing PinQ had 86 ± 2% inversion (Fig. 2F; Supplementary Fig. S6, Supplementary Table S4). After curing the sequenced lytic zone strain of the pCA24N-pinQ plasmid, the inversion percent is 83 ± 3% (Fig. 2F; Supplementary Fig. S6, Supplementary Table S4). Hence, PinQ is active and inverts e14 DNA in one direction more than PinE. We also found the ΔpinE mutation causes a 40-fold decrease in cell survival, and the ΔpinQ ΔpinE double mutations result in a 1000-fold reduction in cell survival with T2 (Supplementary Fig. S1). In addition, the number of surviving colonies in the T2 lytic zone was assayed for the pinQ and pinE pinQ mutants, and, as expected, the number of colonies was lower in the pinQ mutant (57 ± 34) compared to the WT (120 ± 36) and even lower in the pinE pinQ double mutant (31 ± 21, Supplementary Fig. S7). Hence, PinE plays a significantly smaller role than PinQ in T2 anti-phage defense. Notably, pinE and the pinE pinQ mutant have the P segment locked in the non-inverted orientation (Supplementary Fig. S6B and Supplementary Table S4).
Supporting these results, we found P segment inversions in 33% of 562 E. coli genome sequences deposited in the core_nt database of GenBank; for example, E. coli MG1655 has been sequenced in the inverted and noninverted orientations. Furthermore, using the complete cryptic prophage sequences for e14, Qin, rac, and CPS-53 as a query, since e14, rac, and Qin have Pin elements and CPS-53 has a somewhat-related integrase, we also used BLAST to probe the database of non-redundant E. coli genomes and found only inversions in e14.
P segment inverts during T2 infection and reverts in the absence of phage infection
To determine whether the e14 inversion has physiological relevance for surviving T2 phage infection, temporal e14 inversion was measured for the WT strain during T2 infection. We found the WT inverts its e14 DNA significantly from 30% to 62 ± 4% within 4 h and inversion reaches a maximum after 24 h (81 ± 7%) (Fig. 2G). Hence, the P segment of e14 is inverted in the WT strain with native levels of PinQ (i.e. no overproduction) during T2 phage infection, and this inversion constitutes a significant anti-phage defense system.
To determine whether the P segment remains in the inverted orientation after T2 infection, we tested inversion in the absence of T2 phage using the cured strain (that no longer produces PinQ from a pCA24N-pinQ), which was derived from the strain producing PinQ isolated from the T2 lytic zone. We found immediate inversion of the cured strain from 78 ± 3% to 59.25 ± 0.01% in the absence of T2 phage after one regrowth cycle (Supplementary Fig. S8). Hence, the P segment inversion rapidly reverts when the stress of phage infection is removed.
T2 induces pinQ+ expression
To determine whether PinQ expression is induced by T2 phage infection, we used qRT-PCR and found pinQ^+^ is induced 14 ± 2-fold relative to no phage (Supplementary Fig. S9). Due to the high similarity between pinR^+^ and pinQ^+^, and no downstream genes to work as proxies of these genes, we could not decouple the expression of pinR^+^ and pinQ^+^, so this is measure of their combined expression. However, pinE is only induced 4 ± 2-fold, indicating pinR^+^/pinQ^+^ are more active than pinE^+^.
PinQ-mediated T2 phage inhibition decreases phage adsorption
So far, we have shown that PinQ is active and rapidly inverts the E. coli e14 P segment to prevent T2 infection, leading to complete resistance. Since many prophages defend their host from rival phages [45], and since preventing attachment is the most prevalent anti-phage defense [46–48], we explored whether P segment inversion by PinQ affects T2 adsorption. Remarkably, we found production of PinQ significantly decreases T2 phage adsorption to 6 ± 4% at 8 min and to 10 ± 10% adsorption at 16 min (Fig. 3A). Corroborating these results, the cured strain had 2 ± 1% and 2 ± 2% adsorption at 8 min and 16 min, respectively (Fig. 3A). In comparison, E. coli WT strain and empty plasmid (BW25113/pCA24N) had nearly 100% adsorption of T2 phage after 8 min. Hence, PinQ reduces T2 phage adsorption.
StfE2 inhibits T2 adsorption and PinQ reduces porin-related antibiotic import. (A) T2 phage adsorption on E. coli strains. *** P< .005, **** P< .0005. Bars and error bars are the mean and standard deviation of four independent cultures. Dots are individual data points. (B) Fold change in cell survival after 3 h treatment with ampicillin (100 μg/ml, 10× MIC) and ciprofloxacin (5 μg/mL, 10× MIC). Bars and error bars are the mean and standard deviation of four independent cultures. Dots are individual data points. Temporal turbidity (600 nm) during ampicillin (C, 20 μg/ml) and (D, 50 μg/ml) treatment for exponentially growing cells. Data points are the mean and standard deviation of four independent cultures. (E) StfE2 blocks T2 Gp38 adhesion protein from binding receptors FadL and OmpF. Molecular docking analysis shows interactions of StfE2 with T2 phage adhesion protein Gp38 and T2 receptors, FadL and OmpF. Gp38 (D190G) in the T2 escape phage is less inhibited by StfE2. WT is E. coli BW25113, pCA24N is WT/pCA24N, pBS(Kan) is WT/pBS(Kan), PinQ is WT/pCA24N-pinQ, StfE2 is WT/pCA24N-stfE2, StfP2 is WT/pBS(Kan)-stfP2, and cured are cells derived from the PinQ lytic zone colony cured of pCA24N-pinQ. IPTG was used at 1 mM for producing PinQ from pCA24N-pinQ in panels (A, B, and D). IPTG was used at 0.1 mM for producing StfE2 and StfP in panel (A).*
To determine if the decrease in T2 phage adsorption was due to a decrease in T2 receptor binding, we explored whether the PinQ-mediated inversion alters the two T2 receptors, FadL (long-chain fatty acid outer membrane channel) and OmpF (outer membrane porin F) [49] by determining whether transport through the pores formed by FadL and OmpF is affected. Since OmpF imports β-lactam antibiotics [50] and quinolones [51], we reasoned that if the PinQ inversion affects the T2 receptor OmpF, ampicillin and ciprofloxacin resistance would be increased by PinQ. In agreement with our hypothesis, we found cells producing PinQ survive both 10 MIC ampicillin treatment (100 μg/ml) and 10 MIC ciprofloxacin treatment (5 μg/ml) 100-fold and 1000-fold, respectively, relative to cells with the empty plasmid (Fig. 3B). Hence, PinQ reduces the effect of both β-lactam and quinolone antibiotics.
To determine whether the PinQ effect was mediated by OmpF or FadL, we tested which pore was primarily responsible for transporting ampicillin and found deleting ompF protects cells from ampicillin at 2 MIC (20 μg/ml) relative to WT and the ΔfadL mutant (Fig. 3C). Similarly, producing PinQ protects the cell from ampicillin at 5 MIC (50 μg/ml) (Fig. 3D). These results suggest indirectly that the PinQ anti-phage defense mechanism is tied to blocking at least the OmpF T2 phage receptor.
To determine whether PinQ-medicated decrease in T2 phage adsorption is due to repression of fadL and ompF, we tested relative gene expression after phage infection and found 13 ± 2- and 4 ± 1-fold induction of fadL^+^ and ompF^+^, respectively (Supplementary Fig. S9). These results show that in T2 phage adsorption inhibition is not due to a decrease in gene expression of T2 receptors.
StfE2 blocks T2 adsorption
Since the PinQ-mediated P segment e14 inversion affects four genes (tfaP^+^, tfaE^+^, stfP2^+^, or stfE2^+^) of the inverted P segment (Fig. 2A), we sought to determine which proteins encoded by these four genes are responsible for T2 phage inhibition. To avoid polar effects from gene knockouts, we produced each of the four proteins (rather than studying deletions) during T2 infection and found producing the putative tail fiber TfaE (BW25113/pCA24N-tfaE) leads to no survival and producing the putative tail fiber TfaP (BW25113/pCA24N-tfaP) leads to a 20 000-fold reduction in host survival upon T2 infection (Supplementary Fig. S10A). Confirming these results, producing TfaP and TfaE also reduced turbidity during T2 infection (Supplementary Fig. S10B). Moreover, TfaP and TfaE production causes only a slight decrease in the growth rate (Supplementary Fig. S10C); hence, the increased cell death is not related to cell growth. Hence, TfaP and TfaE do not provide T2 anti-phage defense and are different from tail-type proteins in mobile genetic elements that inhibit phages by blocking tail assembly [52].
In contrast, producing StfE2 dramatically reduced T2 phage adsorption with 3 ± 1% phage adsorption at 8 min and 12 ± 6% adsorption at 16 min (Fig. 3A). In addition, producing StfP2 had no effect on T2 adsorption (Fig. 3A). Therefore, PinQ inhibits T2 phage infection by inverting the P segment, which leads to StfE2 production and blocks T2 adsorption.
To further investigate the biological role of StfP, StfP2, StfE, and StfE2, we tested these proteins against representative phages from the BASEL Collection for the families Drexlerviridae (Bas03), Siphoviridae (Bas25), Demerecvidae (Bas26, note T5 belongs to this family), Autographivirae (Bas66, note T3 and T7 belong to this family), and Schitoviridae (Bas69) [33] based on their ability to form large plaques with BW25113. Based on survival after 1 h, we found StfE2 was most effective as it provided protection from all six phages at 0.01 MOI with protection against not only T2 phage (723-fold) but also against phages Bas03 (37-fold, family: Drexlerviridae) and Bas66 (5164-fold, T3-type, family: Autographivirae) (Supplementary Table S5). In addition, StfP2 provided protection against both T2 (107-fold) and Bas69 (290-fold) but was 7-fold less effective than StfE2 for T2 at 1 h (Supplementary Table S5) and 100-fold less effective at 2 h. StfP and StfE were not effective and were sometimes deleterious. Therefore, inversion of the P segment leads to broad phage inhibition primarily by StfE2.
Bioinformatic analyses were then used to explore further how the newly formed by inversion StfE2 protein may affect T2 adsorption. StfE2 residues 47 to 125 include the C-terminal gp53-like domain that is related to pyocins like the R1 pyocin of the prophage LESB58 of Pseudomonas aeruginosa PAO1 and is related to a region of the putative cell-binding protein Gp53 from the myophage AP22 that infects Acinetobacter baumanii [53, 54]. Therefore, we hypothesized that StfE-2 may block the binding of T2 to its receptor; Gp38 is the primary T2 adhesin that binds to receptors to allow for its adsorption [55]. Molecular docking analysis of StfE2, OmpF, and Gp38 shows that StfE2 may alter the binding of Gp38 to OmpF (Fig. 3E). Molecular docking analysis (Fig. 3E) of StfE2, FadL, and Gp38 shows that StfE2 may also alter drastically the interaction of Gp38 with FadL. These results suggest that StfE2 may directly inhibit T2 adsorption to E. coli.
Fitness cost of the P segment inversion
Because of the reversion seen (Supplementary Fig. S8), we investigated whether there was fitness cost to inverting the P segment by testing growth of the cured strain from the T2 lytic zone, with high P segment inversion, using minimal palmitic acid (0.25 wt%) medium, since palmitic acid is a compound likely transported by FadL [56], one of the T2 phage receptors that is modified by StfE2, and found there is a 39 ± 56-fold reduction in growth for the fadL positive control and a 32 ± 21-fold reduction in growth in the cured strain (Supplementary Fig. S11). In contrast, all three strains grew similarly in glucose-minimal medium, so there is no inherent growth defect on non-FadL substrates like glucose. Hence, there is a clear growth disadvantage to the inversion during non-phage conditions.
T2 phage escapes PinQ inhibition by mutating gp38
To glean further insights into the PinQ phage inhibition system, T2 phage was evolved while keeping the production of PinQ unvarying (Fig. 4A). Evolved phages from each cycle were tested for their ability to lyse cells producing PinQ (Fig. 4B). We found that after two rounds of evolution, the PinQ phage inhibition system was increasingly less effective with a collapse in turbidity observed at 3 h by round eight. We then sequenced the T2 genome after eight rounds and compared it to the initial T2 phage (round 0). The T2 escape mutant after eight rounds had point mutations in α-gt (encoding an alpha-glucosyltransferase), in regA (encoding an endoribonuclease translational repressor of early genes), and in rnlA (encoding an RNA ligase and tail fiber attachment catalyst gene) (Supplementary Table S6). Critically, there was also a single bp change in gp38 in ∼53% of the T2 escape population (based on Sanger sequencing of gp38), which encodes the tail fiber protein for host specificity; i.e. for recognizing FadL and OmpF, and results in the single aa substitution D190G in hypervariable region 3 (Gp38*) (Fig. 4C). Hence, T2 phage escapes primarily by mutating the gene responsible for phage adsorption, which corroborates the changes in adsorption we found upon activating PinQ. Fittingly, docking-based analysis revealed the Gp38 substitution D190G renders StfE2 less effective in blocking Gp38* from binding both OmpF and FadL in the presence of StfE2 (Fig. 3E).
T2 escapes by altering cell adhesion. (A) Experimental setup for evolving T2 phage against PinQ anti-phage defense. T2 phage (0.01 MOI) is added to a fresh, exponentially growing culture that produces PinQ continuously (BW25113/pCA24N-pinQ with 1 mM IPTG). After 16 h, T2 phage is purified and added to fresh PinQ-producing culture that was used for the original round of evolution for 8 evolutions; i.e. T2 phage is allowed to evolve while the host is fixed. (B) Temporal turbidity as a function of phage infection time for T2 (0.01 MOI) phage from each round of phage evolution. Data points are the mean and standard deviation of three independent cultures. (C) Comparison of Gp38 adhesin sequences of T2 WT phage (round 0) and T2 escape phage (round 8). The * in amino acid 190 indicates the substitution D190G in the Gp38 adhesion gene in the round 8 escape mutant relative to T2 WT phage, and text above the DNA sequences indicate the conserved adhesin’s segments: four hypervariable segments (H1–H4) and five glycine-rich motifs (G1–G5).
PinQ recombinases are widespread
To investigate whether PinQ-type recombinases are widespread, we searched UniProt [57] for proteins with >90% identity with PinQ. The E. coli K-12 genome contains PinQ with high similarity (99%) to PinQ in Qin cryptic prophage. Similarly, PinR/Q is present in 39 genomes of bacteria (e.g. Escherichia spp. and Shigella spp.), viruses (e.g. Escherichia phages: 2H10, mEp460_ev081, and Tritos), and trematodes (e.g. Clonorchis sinensis, Chinese liver fluke). This suggests that PinQ-type recombinases are widespread, and some may be involved in anti-phage defense by inverting DNA as we found here.
Discussion
Our results demonstrate that E. coli cryptic prophage rac encodes an active serine recombinase PinQ that mediates T2 phage inhibition based on these lines of evidence: PinQ (i) increases host survival upon T2 challenge, (ii) reduces the spread of phages due to their lower level of adsorption, and (iii) inverts the 1797 bp P segment in cryptic prophage e14 within 2 h of T2 phage infection, showing the importance of PinQ at physiological levels, and leads to production of inversion-generated StfE2, which decreases T2 adsorption. Moreover, the inversion of the P segment reverts in the absence of T2 phage, and T2 phage induces pinQ^+^; hence, PinQ directly controls production of StfE2 through inversion of the e14 P segment. This phage inhibition system is comparable to that of other active anti-phage defense systems [15, 58–62] because PinQ-mediated inversion completely inhibits T2 phage infection (Fig. 2D).
Remarkably, instead of inverting the segment of DNA next to the recombinase, PinQ in Qin cryptic prophage inverts the segment of another cryptic prophage, e14, perhaps to make use of a protein captured 4.5 million years ago in Qin [10] rather than creating a separate regulator for stfE2^+^. Alternately, since we found the host is protected from several phages by StfP2 and StfE2 after the inversion (Supplementary Table S5), the host uses inversion to regulate two proteins simultaneously as well as to silence StfE2 and StfP2, in the absence of phage, due to their toxicity (Supplementary Fig. S10D). Moreover, like redundancy in ribosomal loci, perhaps the genetic architecture of inversion is used so that a single region of DNA flanked by inverted repeats may be controlled by several different serine recombinases (PinR, PinE, and PinQ all invert the same e14 P segment with PinQ/PinR dominant) in case of mutation in any one of them; in this way, the cell has a robust mechanism to compete against its worst stress, phages. Hence, our data provide additional evidence that cryptic prophages play an important role in bacterial physiology [10–12] through their recombinases and that the cell fine-tunes its response to phage infection by simultaneously combining resources from distinct cryptic prophages. Similar phenomenon was observed in Salmonella LT2, where Fin, a DNA invertase located in prophage Fels-2, controls the inversion of the H segment next to Hin [63]. Similarly, PinE inverts the phase determinant region of Salmonella typhimurium [64] and G segment of phage Mu [65].
The P segment inversion is also observed in nature in numerous sequenced E. coli strains. For example, CV601 (NCBI Accession: CP067994) contains the four inversion genes (JJT18_12940 matches stfE2, JJT18_12945 matches tfaE, JJT18_12950 matches tfaP, and JJT18_12955 matches stfP2) next to pinE that match the sequence of the inverted form of the P segment of the e14 prophage found in the BW25113 strain isolated here from T2 lytic zones. Critically, the P segment inversion is reversible (Supplementary Fig. S8) since there is a fitness cost to the inversion in K-12 (e.g. slower growth on FadL-mediated substrates such as palmitic acid; Supplementary Fig. S11).
T2 contains six long tail fibers for recognizing primary host receptors and six short tail fibers for recognizing secondary receptors to trigger injection of phage DNA into the host (Fig. 4C) [66]. Of the long tail fibers, adhesin Gp38 determines the host receptor affinity of T-even phages [67], and for T2, it recognizes FadL and OmpF [49]. Gp38 has conserved motifs (i.e. N-terminal domain, four hypervariable segments, five conserved glycine-rich motifs, and Cfin region) needed to mediate adsorption [68]. The T2 phage that escaped the PinQ anti-phage defense has a substitution in the hypervariable region 3 (H3 segment) of Gp38; however, it still contains all the conserved motifs of a Gp38 adhesin. Small substitutions in Gp38 adhesins can change host range function significantly; for example, T2 and SV76 phage Gp38 adhesins differ by 2 amino acids but have two different receptors (T2: OmpF and FadL [49]; SV76: OmpF and FhuA [69]), which suggests that the single aa substitution of the escape mutant identified in this work would cause a large change in the ability of StfE2 to inhibit T2 adsorption. Collectively, we have identified a new paradigm for phage inhibition via ubiquitous recombinases (Fig. 5).
Schematic of the PinQ phage inhibition system. LHS: Without PinQ defense, T2 lyses cells. Free T2 phage lands on the surface of WT cell, and the adhesion protein Gp38 recognizes specific T2 phage receptors, OmpF (green) and FadL (red); the T2 lytic cycle then proceeds. RHS: PinQ-mediated inhibition system. PinQ inverts the 1797 bp P segment of e14 cryptic prophage within the genes stfP+ and stfE+, and produces two new complete genes: stfP2 (begins with stfP+ and ends with reverse complement of stfE+) and stfE2+ (begins with stfE+ and ends with the reverse complement of stfP+). This inversion produces StfE2, which interacts with OmpF and FadL at the same location where Gp38 binds, inhibiting T2 phage adsorption.
Supplementary Material
gkaf1041_Supplemental_File
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Casjens S Prophages and bacterial genomics: what have we learned so far?. Mol Microbiol. 2003; 49:277–300.10.1046/j.1365-2958.2003.03580.x.12886937 · doi ↗ · pubmed ↗
- 2Bobay LM, Touchon M, Rocha EP Pervasive domestication of defective prophages by bacteria. Proc Natl Acad Sci USA. 2014; 111:12127–32.10.1073/pnas.1405336111.25092302 PMC 4143005 · doi ↗ · pubmed ↗
- 3Casjens S, Palmer N, van Vugt R et al. A bacterial genome in flux: the twelve linear and nine circular extrachromosomal DN As in an infectious isolate of the Lyme disease spirochete Borrelia burgdorferi. Mol Microbiol. 2000; 35:490–516.10.1046/j.1365-2958.2000.01698.x.10672174 · doi ↗ · pubmed ↗
- 4Harper G, Hull R, Lockhart B et al. Viral sequences integrated into plant genomes. Annu Rev Phytopathol. 2002; 40:119–36.10.1146/annurev.phyto.40.120301.105642.12147756 · doi ↗ · pubmed ↗
- 5Lander ES, Linton LM, Birren B et al. Initial sequencing and analysis of the human genome. Nature. 2001; 409:860–921.10.1038/35057062.11237011 · doi ↗ · pubmed ↗
- 6Wagner PL, Waldor MK Bacteriophage control of bacterial virulence. Infect Immun. 2002; 70:3985–93.10.1128/iai.70.8.3985-3993.2002.12117903 PMC 128183 · doi ↗ · pubmed ↗
- 7Bair CL, Black LW A type IV modification dependent restriction nuclease that targets glucosylated hydroxymethyl cytosine modified DN As. J Mol Biol. 2007; 366:768–78.10.1016/j.jmb.2006.11.051.17188297 PMC 1855630 · doi ↗ · pubmed ↗
- 8Campbell AM de Bruijn FJ, Lupski JR, Weinstock GM Bacterial Genomes: Physical Structure and Analysis. 1998; Boston, MA Springer US 23–9.10.1007/978-1-4615-6369-3. · doi ↗