Morphological and phylogenetic analyses reveal novel entomopathogenic fungi infecting scale insects and aphids in China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
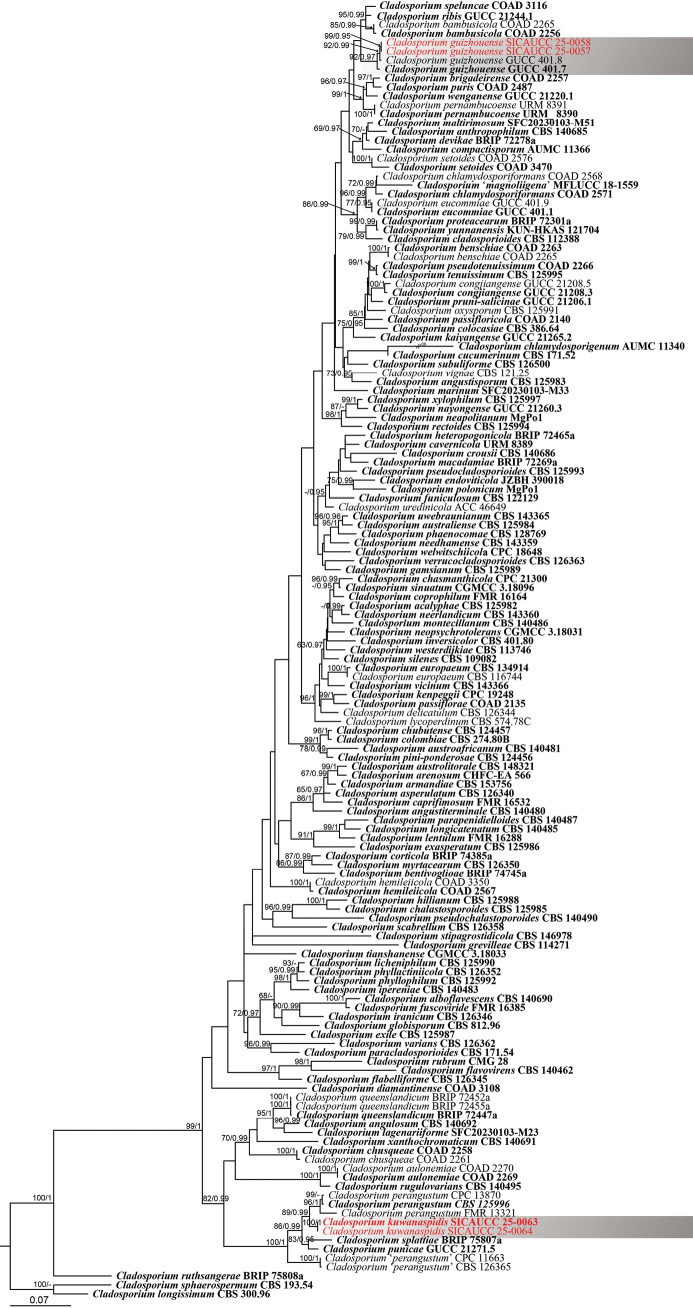 Figure 1
Figure 1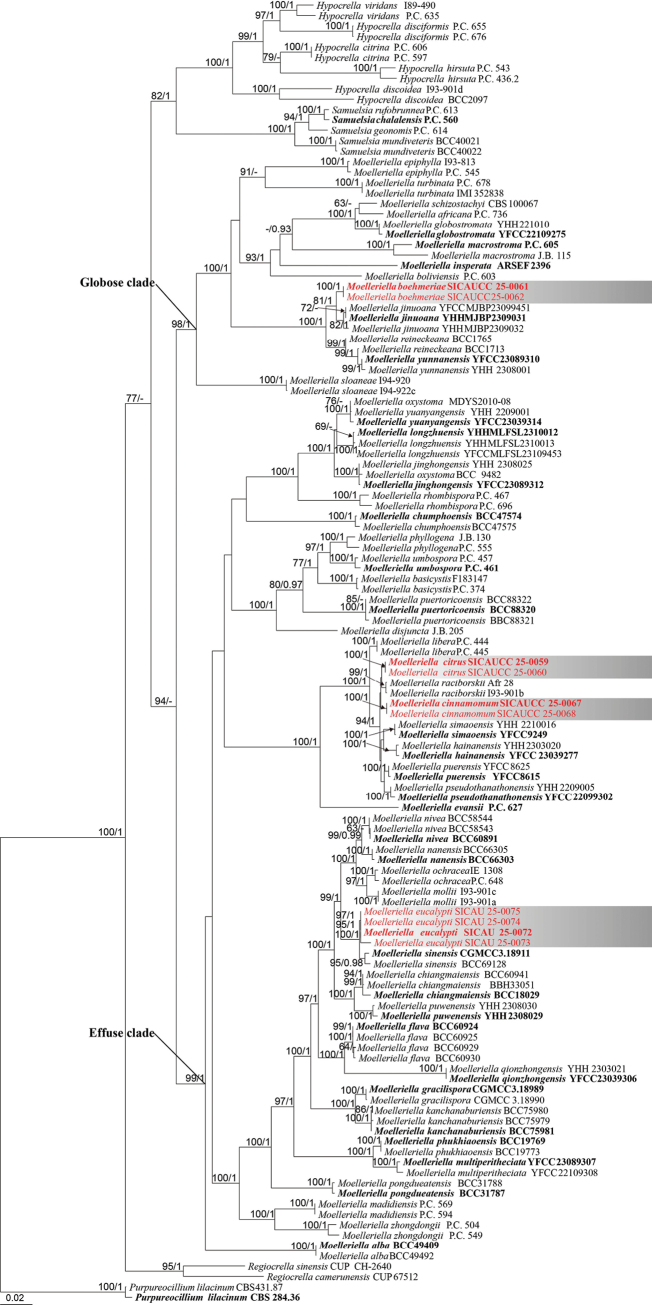 Figure 2
Figure 2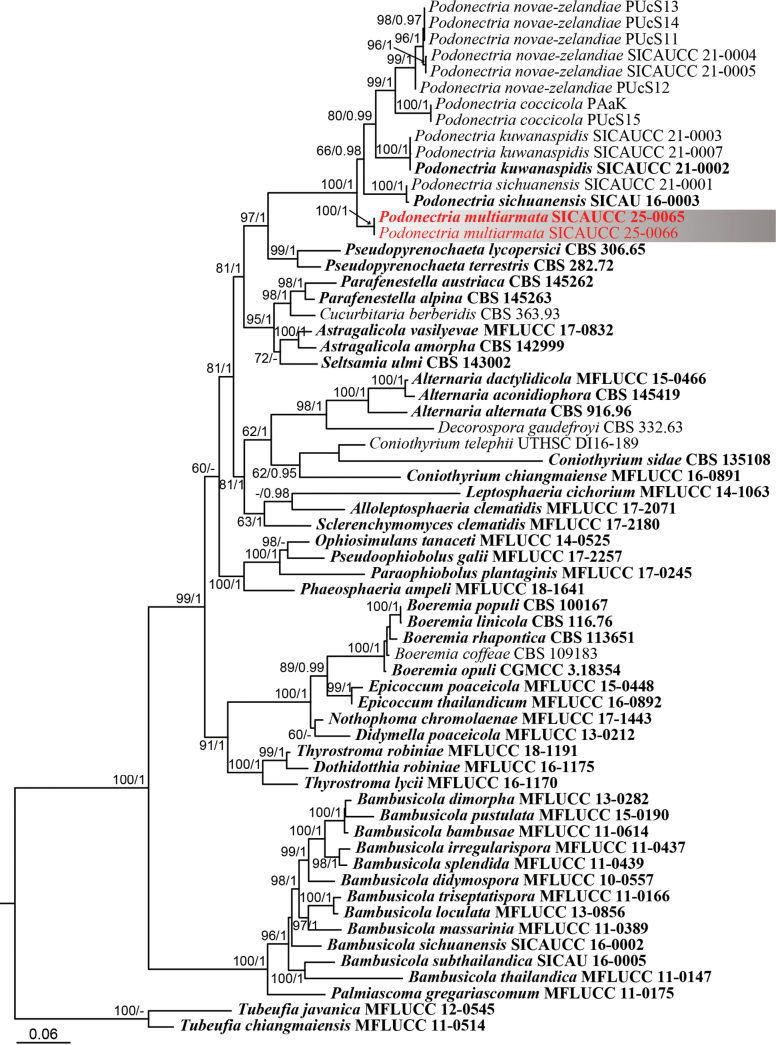 Figure 3
Figure 3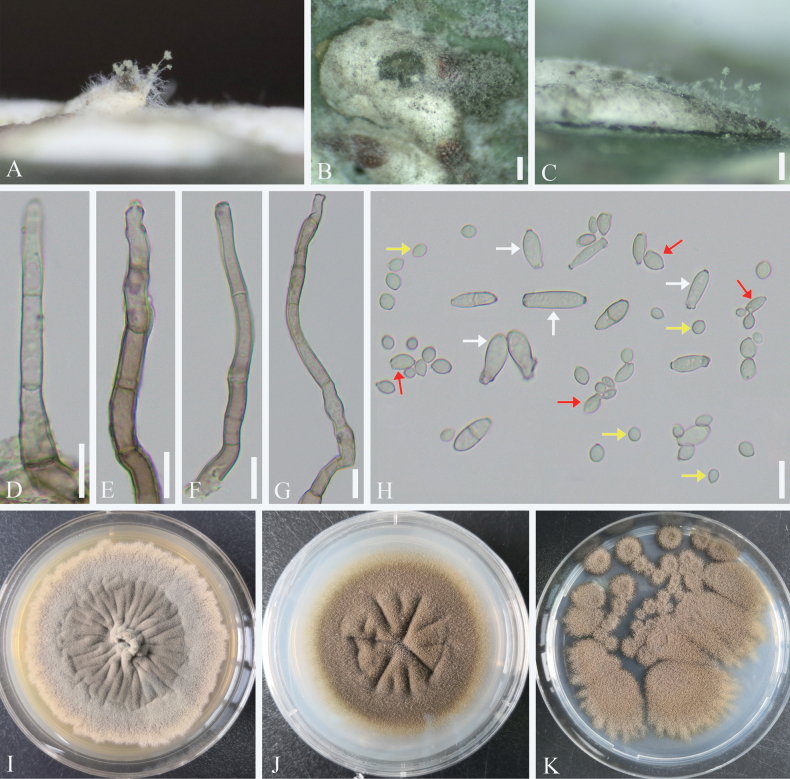 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6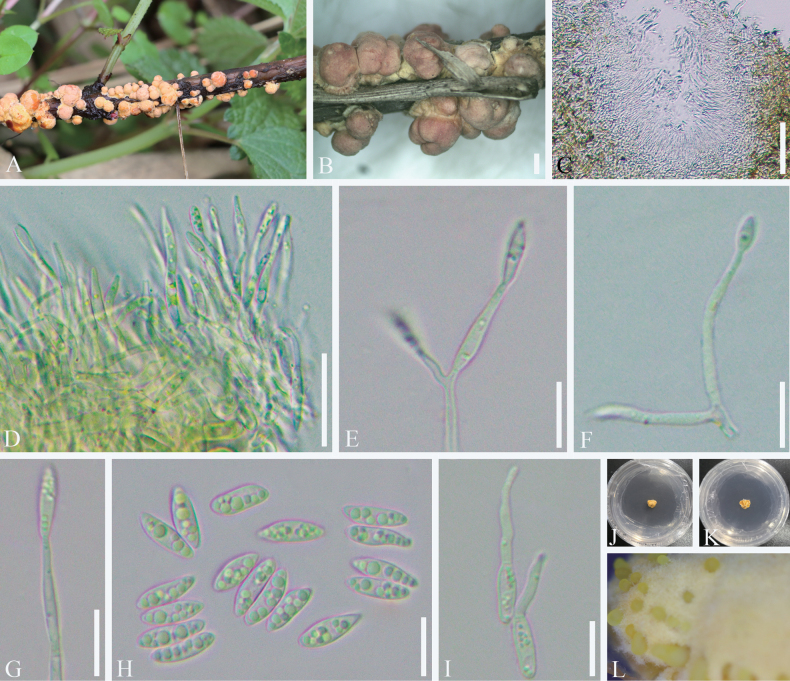 Figure 7
Figure 7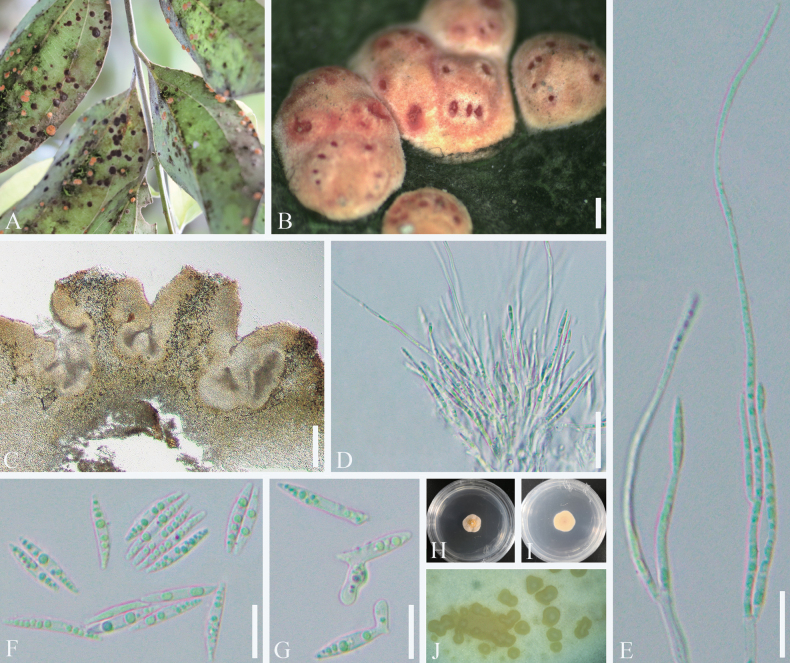 Figure 8
Figure 8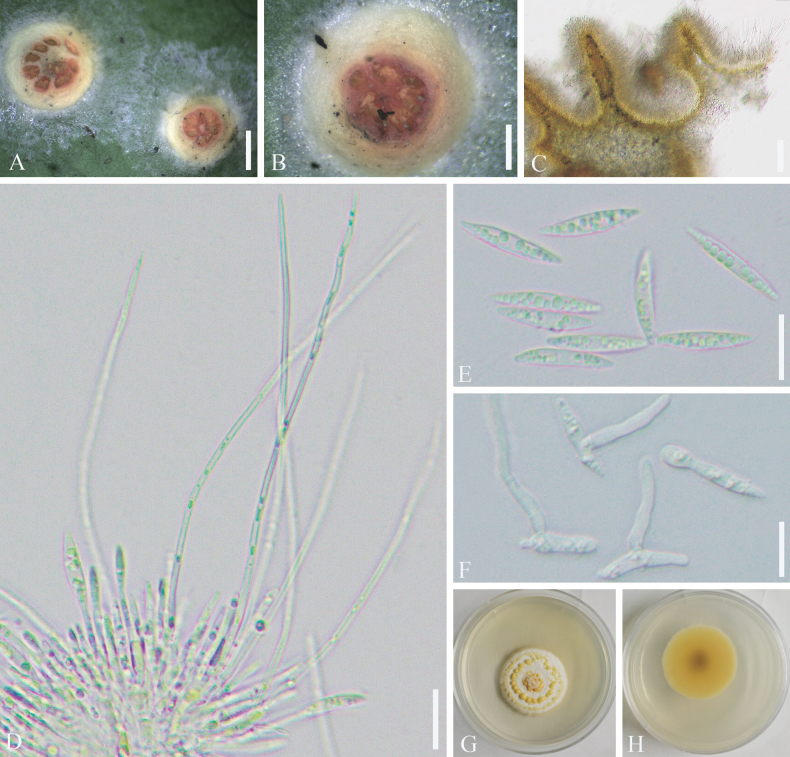 Figure 9
Figure 9 Figure 10
Figure 10| Species | Voucher information |
|
| tef1-α | References |
|---|---|---|---|---|---|
|
| CBS 125982 T |
|
|
|
|
|
| CBS 140690 T |
|
|
|
|
|
| CBS 140692 T |
|
|
|
|
|
| CBS 125983 T |
|
|
|
|
|
| CBS 140480 T |
|
|
|
|
|
| CBS 140685 T |
|
|
|
|
|
| CHFC-EA 566 T |
|
|
|
|
|
| CBS 153756 T |
|
|
|
|
|
| CBS 126340 T |
|
|
|
|
|
| COAD 2269 T |
|
|
|
|
|
| COAD 2270 |
|
|
|
|
|
| CBS 125984 T |
|
|
|
|
|
| CBS 140481 T |
|
|
|
|
|
| CBS 148321 T |
|
|
|
|
|
| COAD 2256 T |
|
|
|
|
|
| COAD 2565 |
|
|
|
|
|
| COAD 2263 T |
|
|
|
|
|
| COAD 2265 |
|
|
|
|
|
| BRIP 74745a T |
|
|
|
|
|
| COAD 2257 T |
|
|
|
|
|
| FMR 16532 T |
|
|
|
|
|
| URM 8389 T |
|
|
|
|
|
| CBS 125985 |
|
|
|
|
|
| CPC 21300 T |
|
|
|
|
|
| AUMC 11340 T |
|
|
| |
|
| COAD 2571 T |
|
|
|
|
|
| COAD 2568 |
|
|
|
|
|
| COAD 2258 T |
|
|
|
|
|
| COAD 2261 |
|
|
|
|
|
| CBS 124457 T |
|
|
|
|
|
| CBS 112388 T |
|
|
|
|
|
| AUMC 11366 T |
|
|
| |
|
| CBS 386.64 T |
|
|
|
|
|
| CBS 274.80B T |
|
|
|
|
|
| GUCC 21208.3 T |
|
|
| Yang et al. (2023) |
|
| GUCC 21208.5 |
|
|
| Yang et al. (2023) |
|
| FMR 16164 T |
|
|
|
|
|
| BRIP 74385a T |
|
|
|
|
|
| CBS 140686 T |
|
|
|
|
|
| CBS 171.52 T |
|
|
|
|
|
| CBS 126344 |
|
|
|
|
|
| BRIP 72278a T |
|
|
| |
|
| COAD 3108 T |
|
|
|
|
|
| JZBH 390018 T |
|
|
|
|
|
| GUCC 401.1 T |
|
|
|
|
|
| GUCC 401.9 |
|
|
| |
|
| CBS 134914 T |
|
|
|
|
|
| CBS 116744 |
|
|
|
|
|
| CBS 125986 |
|
|
|
|
|
| CBS 125987 |
|
|
|
|
|
| CBS 126345 |
|
|
| |
|
| CBS 140462 T |
|
|
|
|
|
| CBS 122129 T |
|
|
|
|
|
| FMR 16385 T |
|
|
|
|
|
| CBS 125989 T |
|
|
|
|
|
| CBS 812.96 T |
|
|
|
|
|
| CBS 114271 T |
|
|
|
|
|
| GUCC 401.7 T |
|
|
|
|
|
| GUCC 401.8 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| COAD 2567 T |
|
|
|
|
|
| COAD 3350 |
|
|
|
|
|
| BRIP 72465a T |
|
|
|
|
|
| CBS 125988 T |
|
|
|
|
|
| CBS 401.80 T |
|
|
|
|
|
| CBS 140483 T |
|
|
|
|
|
| CBS 126346 T |
|
|
|
|
|
| GUCC 21265.2 T |
|
|
| Yang et al. (2023) |
|
| CPC 19248 T |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| SFC20230103-M23 T |
|
|
|
|
|
| FMR 16288 T |
|
|
|
|
|
| CBS 125990 |
|
|
|
|
|
| CBS 140485 |
|
|
|
|
|
| CBS 300.96 |
|
|
| |
|
| CBS 574.78C |
|
|
| |
|
| BRIP 72269a T |
|
|
|
|
|
| MFLUCC 18-1559 T |
|
|
| |
|
| SFC20230103-M51 T |
|
|
|
|
|
| SFC20230103-M33 T |
|
|
|
|
|
| CBS 140486 T |
|
|
|
|
|
| CBS 126350 T |
|
|
| Yang et al. (2023) |
|
| GUCC 21260.3 T |
|
|
|
|
|
| MgPo1 T |
|
|
|
|
|
| CBS 143359 T |
|
|
|
|
|
| CBS 143360 T |
|
|
|
|
|
| CGMCC 3.18031 T |
|
|
|
|
|
| CBS 125991 |
|
|
|
|
|
| CBS 171.54 T |
|
|
|
|
|
| CBS 140487 T |
|
|
|
|
|
| COAD 2135 T |
|
|
|
|
|
| COAD 2140 T |
|
|
| |
|
| CBS 125996 T |
|
|
|
|
|
| CBS 126365 |
|
|
|
|
|
| CPC 11663 |
|
|
|
|
|
| CPC 13870 |
|
|
|
|
|
| FMR 13321 |
|
|
|
|
|
| URM 8390 T |
|
|
|
|
|
| URM 8391 |
|
|
|
|
|
| CBS 128769 |
|
|
|
|
|
| CBS 126352 |
|
|
| |
|
| CBS 125992 T |
|
|
|
|
|
| CBS 124456 T |
|
|
|
|
|
| MgPo1 T |
|
|
|
|
|
| BRIP 72301a T |
|
|
| |
|
| GUCC 21206.1 T |
|
|
| Yang et al. (2023) |
|
| CBS 125993 T |
|
|
|
|
|
| CBS 140490 T |
|
|
|
|
|
| COAD 2266 T |
|
|
|
|
|
| GUCC 21271.5 T |
|
|
| Yang et al. (2023) |
|
| COAD 2487 T |
|
|
|
|
|
| BRIP 72447a T |
|
|
|
|
|
| BRIP 72452a |
|
|
|
|
|
| BRIP 72455a |
|
|
|
|
|
| CBS 125994 T |
|
|
|
|
|
| GUCC 21244.1 T |
|
|
| Yang et al. (2023) |
|
| CMG 28 T |
|
|
|
|
|
| CBS 140495 T |
|
|
|
|
|
| BRIP 75808a T |
|
|
|
|
|
| CBS 126358 T |
|
|
|
|
|
| COAD 3470 T |
|
|
|
|
|
| COAD 2576 |
|
|
|
|
|
| CBS 109082 T |
|
|
|
|
|
| CGMCC 3.18096 T |
|
|
|
|
|
| COAD 3116 T |
|
|
|
|
|
| BRIP 75807a |
|
|
|
|
|
| CBS 193.54 |
|
|
|
|
|
| CBS 146978 T |
|
|
|
|
|
| CBS 126500 T |
|
|
|
|
|
| CBS 125995 T |
|
|
|
|
|
| CGMCC 3.18033 T |
|
|
| |
|
| ACC 46649 |
|
|
|
|
|
| CBS 143365 T |
|
|
|
|
|
| CBS 126362 T |
|
|
|
|
|
| CBS 126363 T |
|
|
|
|
|
| CBS 143366 T |
|
|
|
|
|
| CBS 121.25 |
|
|
|
|
|
| CPC 18648 T |
|
|
| Yang et al. (2023) |
|
| GUCC 21220.1 T |
|
|
|
|
|
| CBS 113746 T |
|
|
|
|
|
| CBS 140691 T |
|
|
|
|
|
| CBS 125997 T |
|
|
|
|
|
| KUN-HKAS 121704 T |
|
|
| |
|
| CBS 300.96 T |
|
|
|
| Species | Voucher information |
| rpb1 | tef1-α | References |
|---|---|---|---|---|---|
|
| P.C. 606 |
|
|
|
|
|
| P.C. 597 |
|
|
|
|
|
| BCC2097 |
|
|
| |
|
| I93-901d |
|
|
|
|
|
| P.C. 655 |
|
|
|
|
|
| P.C. 676 |
|
|
|
|
|
| P.C. 436.2 |
|
|
|
|
|
| P.C. 543 |
|
|
|
|
|
| P.C. 635 |
|
|
|
|
|
| I89-490 |
|
|
|
|
|
| P.C. 736 |
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
| |
|
| P.C. 374 |
|
|
|
|
|
| F183147 |
|
|
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| P.C. 603 |
|
|
|
|
|
|
|
|
| ||
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| J.B. 205 |
|
|
| |
|
| P.C. 545 |
|
|
|
|
|
| I93-813 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| P.C. 627 T |
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
| ||
|
| YFCC 22109275 T |
|
|
|
|
|
| YHH 221010 |
|
|
|
|
|
| CGMCC 3.18989 T |
|
|
|
|
|
| CGMCC 3.18990 |
|
|
|
|
|
| YHH 2303020 |
|
|
|
|
|
| YFCC 23039277 T |
|
|
| |
|
| ARSEF 2396 T |
|
|
|
|
|
| YFCC 23089312 T |
|
|
| |
|
| YHH 2308025 |
|
|
|
|
|
| YHH MJBP2309031 T |
|
|
|
|
|
| YHH MJBP2309032 |
|
|
|
|
|
| YFCC MJBP23099451 |
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
| ||
|
| P.C. 444 |
|
|
|
|
|
| P.C. 445 |
|
|
|
|
|
| YHH MLFSL2310012 T |
|
|
|
|
|
| YHH MLFSL2310013 |
|
|
|
|
|
| YFCC-MLFSL23109453 |
|
|
| |
|
| P.C. 605 T |
|
|
|
|
|
| J.B. 115 |
|
|
|
|
|
| P.C. 569 |
|
|
|
|
|
| P.C. 594 |
|
|
|
|
|
| I93-901a |
|
|
|
|
|
| I93-901c |
|
|
|
|
|
| YFCC 23089307 T |
|
|
|
|
|
| YFCC 22109308 |
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
| IE 1308 |
|
|
|
|
|
| P.C. 648 |
|
|
|
|
|
| MDYS2010-08 |
|
|
| Yang et al. (2023) |
|
| BCC 9482 |
|
|
| Yang et al. (2023) |
|
|
|
|
| ||
|
|
|
|
| ||
|
| P.C. 555 |
|
|
|
|
|
| J.B. 130 |
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
| |
|
| YFCC 22099302 T |
|
|
| |
|
| YHH 2209005 |
|
|
|
|
|
| YFCC 8615 T |
|
|
|
|
|
| YFCC 8625 |
|
|
|
|
|
|
|
|
| ||
|
|
|
|
| ||
|
|
|
|
| ||
|
| YHH 2308029 T |
|
|
|
|
|
| YHH 2308030 |
|
|
|
|
|
| YHH 2303021 |
|
|
|
|
|
| YFCC 23039306 T |
|
|
| |
|
| Afr 28 |
|
|
|
|
|
| I93-901b |
|
|
|
|
|
| BCC1713 |
|
|
| |
|
| BCC1765 |
|
|
| |
|
| P.C. 467 |
|
|
|
|
|
| P.C. 696 |
|
|
|
|
|
| CBS 100067 |
|
|
|
|
|
| YFCC 9249 T |
|
|
| Yang et al. (2023) |
|
| YHH 2210016 |
|
|
| Yang et al. (2023) |
|
| CGMCC 3.18911 T |
|
|
| |
|
|
|
|
|
| |
|
| I94-920 |
|
|
|
|
|
| I94-922c |
|
|
|
|
|
| P.C. 678 |
|
|
|
|
|
| IMI 352838 |
|
|
|
|
|
| P.C. 457 |
|
|
|
|
|
| P.C. 461 T |
|
|
|
|
|
| YFCC 23039314 T |
|
|
| |
|
| YHH 2209001 |
|
|
| |
|
| YFCC 23089310 T |
|
|
| |
|
| YHH 2308001 |
|
|
|
|
|
| P.C. 504 |
|
|
|
|
|
| P.C. 549 |
|
|
|
|
|
| CBS 284.36 T |
|
|
|
|
|
| CBS 431.87 |
|
|
|
|
|
| CUP 67512 |
|
|
|
|
|
| CUP CH-2640 |
|
|
|
|
|
| P.C. 560 T |
|
|
|
|
|
| P.C. 614 |
|
|
|
|
|
|
|
|
| ||
|
|
|
|
| ||
|
| P.C. 613 |
|
|
|
| Species | Voucher information |
|
|
| tef1-α | rpb2 | References |
|---|---|---|---|---|---|---|---|
|
| MFLUCC 17-2071 T |
|
|
|
|
|
|
|
| CBS 145419 T |
|
|
|
| ||
|
| CBS 916.96 T |
|
|
|
|
|
|
|
| MFLUCC 15-0466 T |
|
|
|
|
| |
|
| CBS 142999 T |
|
|
|
|
| |
|
| MFLUCC 17-0832 T |
|
|
|
|
|
|
|
| MFLUCC 11-0614 T |
|
|
|
|
| |
|
| MFLUCC 10-0557 T |
|
|
|
|
|
|
|
| MFLUCC 13-0282 T |
|
|
|
|
| |
|
| MFLUCC 11-0437 T |
|
|
|
|
| |
|
| MFLUCC 13-0856 T |
|
|
|
|
|
|
|
| MFLUCC 11-0389 T |
|
|
|
|
| |
|
| MFLUCC 15-0190 T |
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
| MFLUCC 11-0439 T |
|
|
|
|
| |
|
|
|
|
|
|
|
| |
|
| MFLUCC 11-0147 T |
|
|
|
|
|
|
|
| MFLUCC 11-0166 T |
|
|
|
| ||
|
| CBS 109183 |
|
|
|
|
| |
|
| CBS 116.76 T |
|
|
|
| Jayawardena et al. | |
|
| CGMCC 3.18354 T |
|
|
| (2019) | ||
|
| CBS 100167 T |
|
|
|
| ||
|
| CBS 113651 T |
|
|
| |||
|
| MFLUCC 16-0891 T |
|
|
|
|
| |
|
| CBS 135108 T |
|
|
|
|
| |
|
| UTHSC:DI16-189 |
|
|
|
| ||
|
| CBS 363.93 |
|
|
| |||
|
| CBS 332.63 |
|
|
| |||
|
| MFLUCC 13-0212 T |
|
|
|
| ||
|
| MFLUCC 16-1175 T |
|
|
|
|
|
|
|
| MFLUCC 15-0448 T |
|
|
|
| ||
|
| MFLUCC 16-0892 T |
|
|
|
| ||
|
| MFLUCC 14-1063 T |
|
|
| |||
|
| MFLUCC 17-1443 T |
|
|
|
| ||
|
| MFLUCC 14-0525 T |
|
|
|
| ||
|
| MFLUCC 11-0175 T |
|
|
|
|
| |
|
| CBS 145263 T |
|
|
|
|
| |
|
| CBS 145262 T |
|
|
|
|
| |
|
| MFLUCC 17-0245 T |
|
|
|
|
| |
|
| MFLUCC 18-1641 T |
|
|
|
|
| |
|
| PAaK |
|
|
| |||
|
| PUcS15 |
|
|
| |||
|
|
|
|
|
|
| Xu et al. (2021) | |
|
|
|
|
|
|
| Xu et al. (2021) | |
|
|
|
|
|
|
| Xu et al. (2021) | |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| PUcS14 |
|
|
| |||
|
| PUcS13 |
|
|
| |||
|
| PUcS12 |
|
|
| |||
|
| PUcS11 |
|
|
| |||
|
|
|
|
|
|
| Xu et al. (2021) | |
|
|
|
|
|
|
| Xu et al. (2021) | |
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
| Xu et al. (2021) | |
|
| MFLUCC 17-2257 T |
|
|
|
|
| |
|
| CBS 306.65 T |
|
|
|
| ||
|
| CBS 282.72 T |
|
|
|
| ||
|
| MFLUCC 17-2180 T |
|
|
|
|
|
|
|
| CBS 143002 T |
|
|
|
|
|
|
|
| MFLUCC 16-1170 T |
|
|
|
|
|
|
|
| MFLUCC 18-1191 T |
|
|
|
|
|
|
|
| MFLUCC 11-0514 T |
|
|
|
|
| |
|
| MFLUCC 12-0545 T |
|
|
|
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEntomopathogenic Microorganisms in Pest Control · Forest Insect Ecology and Management · Plant Pathogens and Fungal Diseases
Introduction
To ensure environmental safety, fungal agents are considered key biological regulators in the application of insecticides for pest control (Roy and Cottrell 2008; Sharma and Sharma 2021). The initial recommendation to use microbial insect pathogens for controlling insects was proposed by LeConte (1874) and Pasteur (1874). Subsequently, the first successful application of Metarhizium anisopliae was conducted in Russia against Bothynoderes punctiventris (Coleoptera: Curculionidae) by Krassilstschik (1888). This achievement built upon the earlier identification of the microbial agent, initially named Entomophthora anisopliae, which targeted Anisoplia austriaca (Coleoptera: Scarabaeidae).
To date, more than 1,000 species of entomopathogenic fungi have been documented across over 100 genera (Hywel-Jones 1993; Vega et al. 2012; Hawksworth and Lücking 2017; Corallo et al. 2019; Wu et al. 2024). Among these species, such as Akanthomyces Lebert, Beauveria Vuill., Cordyceps Fr. (=Isaria Pers.), Lecanicillium W. Gams & Zare, and Metarhizium Sorokin, are widely recognized for their potential in biological control. Notably, species like A. aculeatus, B. bassiana, B. brongniartii, C. fumosorosea, L. lecanii (≡Verticillium lecanii), L. muscarium, L. longisporum, and M. anisopliae are commonly employed in pest management strategies (Faria and Wraight 2007; McKinnon et al. 2017; Corallo et al. 2019; Stone and Bidochka 2020; Nicoletti and Becchimanzi 2020; Nishi et al. 2021; Khonsanit et al. 2024). For example, 171 products have been developed as biocontrol agents, primarily based on B. bassiana, B. brongniartii, C. fumosorosea, and M. anisopliae (Faria and Wraight 2007). Beauveria bassiana, marketed as “Mycotrol”, has been utilized as a commercial mycoinsecticide to target a broad spectrum of insect pests in North America (Wraight et al. 2021). Similarly, the commercial product “Green Muscle,” derived from M. acridum (≡M. anisopliae var. acridum), has proven effective against locust and grasshopper pests in Africa (Niassy and Diarra 2011). In addition, entomopathogenic fungi exhibit diverse nutritional behaviors—functioning as biotrophs, necrotrophs, or hemibiotrophs (Vega et al. 2009)—and demonstrate additional ecological roles, including rhizosphere colonization, fungal endophytism, plant disease antagonism, and enhancement of plant growth or stress resistance (Kabaluk and Ericsson 2007; Kim et al. 2008; Ownley et al. 2008; Vega 2008; Pava-Ripoll et al. 2011; Akutse et al. 2013; Jaber and Enkerli 2017; Jaber and Ownley 2017; Mantzoukas and Eliopoulos 2020; Sharma and Sharma 2021; Muola et al. 2023).
Insect-associated fungi have been widely reported across the extensive regions south of the Yellow River basin, with most species displaying distribution patterns characteristic of tropical, subtropical, and temperate zones (Liang et al. 2016; Chen et al. 2021; Wei et al. 2022; Xiao et al. 2023; Chuang et al. 2024; Wang et al. 2024, 2025), but have been rarely documented in the Sichuan region. The development of entomogenous fungi is closely linked to vegetation and insect populations, as well as climatic and soil conditions (Hajek and Leger 1994; Quesada-Moraga et al. 2007, 2024; Vega et al. 2009; Medo and Cagáň 2011), although experimental validation in this area remains limited. Scattered studies have indicated that entomogenous fungi are more commonly found in areas with rich vegetation, relatively high humidity, suitable temperatures, and shaded environments. Chuang et al. (2024) found that in Taiwan, China, entomogenous fungi belonging to the Cordycipitaceae are more readily collected in regions with an average temperature of 19–20.5 °C, rainfall ranging from 2,300 to 3,150 mm, and relative humidity between 81% and 82.5%. Wang et al. (2025) collected and described two new genera and 13 new species within the Clavicipitaceae, associated with whiteflies and scale insects on various host plants in Yunnan and Hainan, China.
Sichuan is recognized as one of the key biodiversity research hotspots, owing to its diverse ecosystems, complex habitat conditions, and rich plant species. While the region supports a wide variety of insect fauna, entomogenous fungi have not been systematically studied, and their species diversity is likely significantly underestimated. In our ongoing search for entomogenous fungi associated with miniature insects, several new taxa of entomopathogenic fungi (Nectriaceae; Podonectriaceae) were isolated from scale insects collected in Sichuan Province, primarily from bamboo and walnut hosts (Xu et al. 2021a; Liu et al. 2023). To further explore the diversity of entomogenous fungi, we conducted field collections at multiple sites across Sichuan. Detailed morphological descriptions are provided along with relevant ecological data.
Comprehensive phylogenetic analyses were performed using a seven-locus dataset (ITS, LSU, SSU, tef1-α, rpb1, rpb2, and act) to ensure accurate taxonomic position. This study enriches the entomogenous fungi database in Sichuan and offers valuable strains for sustainable pest control in agriculture and forestry.
Materials and methods
Fungal collection and isolation
Specimens consisting of whole leaves or branches bearing fruiting bodies were collected from Chengdu, Dazhou, Guangan, Leshan, and Meishan in Sichuan Province, China, placed in sterilized plastic bags, and transported to the laboratory for analysis. The procedure for obtaining axenic cultures, as described by Chomnunti et al. (2014), involved isolating pure cultures, transferring them to PDA slants for 10 days of cultivation, and subsequently storing them at 4 °C. The specimens were deposited in the Herbarium of Sichuan Agricultural University (SICAU), Chengdu, China, while the strains were stored in the Culture Collection of Sichuan Agricultural University (SICAUCC), China.
Morphological observations
Fruiting bodies were observed and photographed using a dissecting microscope NVTGG (Shanghai Advanced Photoelectric Technology Co. Ltd., Shanghai, China) fitted with a VS-800C micro-digital camera (Shenzhen Weishen Times Technology Co. Ltd., Shenzhen, China). The dimensions of conidiomata, paraphyses, conidiophores, conidiogenous cells, and conidia were measured from field samples and photographed using an Olympus BX43 compound microscope equipped with an Olympus DP22 digital camera, in conjunction with ACDSee v3.1 software. Measurements were made using Tarosoft® Image Frame Work v.0.9.7 (Tarosoft (R), Nontha Buri, Thailand). Lactophenol cotton blue reagent was used to observe the conidiogenous structures and determine the number of septa. To observe and document the color and texture of the colonies, fresh plates were prepared from purified colonies and incubated at 25 °C for 1 to 3 weeks.
DNA extraction, amplification, and sequencing
Total genomic DNA was extracted from mycelia grown on PDA or single fruiting bodies using the Plant Genomic DNA extraction kit (Tiangen, China). Primer pairs ITS5 and ITS4 (White et al. 1990), NS1 and NS4 (White et al. 1990), LR0R and LR5 (Vilgalys and Hester 1990), EF1-983F and EF1-2218R (Rehner and Buckley 2005), RPB1-Ac and RPB1-Cr (Lombard et al. 2015), fRPB2-5F and fRPB2-7cR (Liu et al. 2000), and ACT512F/ACT-783R (Carbone and Kohn 1999) were used to amplify the internal transcribed spacer (ITS), the partial small subunit nuclear rDNA (SSU), the nuclear ribosomal large subunit (LSU), the translation elongation factor tef1-α, the largest subunit of RNA polymerase II (rpb1), the second largest subunit of RNA polymerase II (rpb2), and the actin gene (act), respectively. Additionally, for Cladosporium species, EF1728F and EF2 (Lombard et al. 2015) were used to amplify tef1-α.
Polymerase chain reaction (PCR) was performed in a 25 μL reaction mixture containing 22 μL Master Mix (Beijing TsingKe Biotech Co. Ltd., Beijing, China), 1 μL DNA template (10–30 ng/μL), and 1 μL of each primer (120–150 ng/μL). The amplification reactions were performed as described by Lombard et al. (2015), Dai et al. (2016), and Wang et al. (2022a). PCR products were sequenced at TsingKe Biological Technology Co. Ltd., Chengdu, China. The newly generated sequences were deposited in GenBank.
Phylogenetic analyses
Phylogenetic analyses were conducted using sequences from Cladosporium (ITS, act, and tef1-α), Moelleriella (LSU, rpb1, and tef1-α), and Podonectria (ITS, LSU, SSU, tef1-α, and rpb2). Multigene sequences from various species (see Tables 1–3) were obtained from GenBank, along with additional sequences generated in this study. DNA alignments were carried out using the MAFFT v.7.429 online service (Katoh et al. 2019), and ambiguous regions were excluded using BioEdit version 7.0.5.3 (Hall 1999). Multigene sequences were concatenated using Mesquite software (Maddison and Maddison 2019). Multigene phylogenetic analyses were conducted using maximum likelihood (ML) and Bayesian inference (BI) methods. The optimal nucleotide substitution model was identified with MrModeltest v.2.2 (Nylander 2004).
ML and BI analyses were performed using the CIPRES Science Gateway web server (Miller et al. 2010). For the ML analysis, RAxML-HPC2 on XSEDE (v.8.2.10) (Stamatakis 2014) was employed with the GTR+GAMMA substitution model and 1,000 bootstrap iterations. For BI analyses, the best-fit models were selected using MrModeltest v.2.2 for each dataset: Cladosporium (ITS: SYM+I+G, act: GTR+G, tef1-α: GTR+I+G), Moelleriella (LSU: GTR+I+G, rpb1: GTR+I+G, tef1-α: GTR+I+G), and Podonectriaceae (ITS: GTR+I+G, LSU: GTR+I+G, SSU: GTR+I+G, tef1-α: GTR+I+G, rpb2: GTR+I+G). The analyses were computed with six simultaneous Markov Chain Monte Carlo (MCMC) chains, run for 1,000,000 to 10,000,000 generations with a sampling frequency of every 100 generations. The burn-in fraction was set to 0.25, and the run was automatically terminated when the average standard deviation of split frequencies fell below 0.01.
Phylogenetic trees were visualized using FigTree v.1.4.3 (Rambaut and Drummond 2016) and further edited with Adobe Illustrator CS6 (Adobe Systems Inc., United States). Maximum likelihood bootstrap values (MLBS) of 60% or greater and Bayesian posterior probabilities (BIPP) of 0.95 or higher were indicated on the trees.
Abbreviations
act The actin gene
BI Bayesian inference
bp Base pair
BIPP Bayesian inference posterior probabilities
DNA Deoxyribonucleic acid
ITS internal transcribed spacer
LSU The nuclear ribosomal large subunit
MEA Malt extract agar medium
ML Maximum likelihood
MLBS Maximum likelihood bootstrap proportions
PCR Polymerase chain reaction
PDA Potato dextrose agar medium
rpb1 The largest subunits of RNA polymerase II
rpb2 The RNA polymerase II second largest subunit
SICAU The herbarium of Sichuan Agricultural University
SICAUCC The culture collection in Sichuan Agricultural University
SNA Synthetic low nutrient agar medium
SSU The partial small subunit nuclear rDNA
tef1-α The translation elongation factor 1α
Results
Sequencing and phylogenetic analyses
Nucleotide BLAST results from GenBank indicated that our isolates were related to Cladosporium, Moelleriella, and Podonectria. To determine the phylogenetic relationships of our isolates within these genera, three phylogenetic analyses were conducted in this study.
Phylogenetic analysis of the genus Cladosporium
An initial analysis of isolates from the three Cladosporium species complexes (C. cladosporioides, C. herbarum, and C. sphaerospermum) revealed that all Cladosporium isolates examined in this study belong to the C. cladosporioides species complex. The final alignment consisted of 122 taxa within the C. cladosporioides species complex, rooted with C. sphaerospermum (CBS 193.54) and C. longissimum (CBS 300.96) (C. sphaerospermum species complex) (Fig. 1). The alignment contained 2,008 characters (act = 1–273, ITS = 274–973, tef1-α = 974–2,008), including gaps. The best-scoring randomized accelerated maximum likelihood tree, with a final likelihood value of –20,491.871768, is presented. The matrix had 1,070 distinct alignment patterns, with 43.95% undetermined characters or gaps. The gamma distribution shape parameter α = 0.232201, and the tree length = 5.319454. The Bayesian analysis resulted in 200,002 trees after 10,000,000 generations, from which 150,002 were used for calculating posterior probabilities after the first 25% of trees, representing the burn-in phase, were discarded. Phylogenetic trees generated from ML and BI analyses were similar in overall topologies (Fig. 1). Within the C. cladosporioides species complex, our two isolates formed a clade with C. guizhouense, exhibiting strong bootstrap support values (92% MLBS/0.97 BIPP). Meanwhile, another two isolates clustered in a separate clade, distinct from known species within this complex, suggesting that they represent new species. Cladosporium kuwanaspidis sp. nov. (SICAUCC 25-0063, SICAUCC 25-0064) formed a monophyletic group closely related to C. perangustum.
Phylogenetic tree based on maximum likelihood analysis of the combined act, ITS, and tef1-α sequence alignment dataset of the Cladosporium cladosporioides complex. Numbers above the nodes represent MLBS values (≥ 60%, left) and BIPP values (≥ 0.95, right), with lower values denoted as “–”. The tree is rooted to C. sphaerospermum (CBS 193.54) and C. longissimum (CBS 300.96). Ex-type strains are highlighted in bold, and isolates from the present study are shown in red.
Phylogenetic analysis of the genus Moelleriella
A combined LSU–rpb2–tef1-α dataset, consisting of 62 taxa from the Clavicipitaceae and one outgroup taxon, Purpureocillium lilacinum (CBS 284.36, CBS 431.87) from the Ophiocordycipitaceae, was used for phylogenetic analyses (Fig. 2). The alignment contained 2,729 characters (LSU = 1–1,040, rpb1 = 1,041–1,788, tef1-α = 1,789–2,729), including gaps. The best-scoring tree with a final likelihood value of –27,989.959271 is presented. The matrix had 1,379 distinct alignment patterns, with 19.07% undetermined characters or gaps. The gamma distribution shape parameter α = 0.246493, and the tree length = 2.839149. The Bayesian analysis resulted in 31,182 trees after 50,000,000 generations, from which 23,388 were used for calculating posterior probabilities after the first 25% of trees, representing the burn-in phase, were discarded. Phylogenetic trees generated from ML and BI analyses were similar in overall topologies. Phylogeny from the combined sequence data analysis indicates that all the isolates belong to Moelleriella (Fig. 2). The isolates clustered in clades different from known species of Moelleriella, indicating that they are new species. Moelleriella eucalypti sp. nov. (SICAU 25-0072, SICAU 25-0073, SICAU 25-0074, SICAU 25-0075) formed a monophyletic group closely related to M. sinensis. Moelleriella boehmeriae sp. nov. (SICAUCC 25-0061 and SICAUCC 25-0062) formed a new clade closely related to M. jinuoana. Finally, M. cinnamomum sp. nov. (SICAUCC 25-0067, SICAUCC 25-0068) and M. citrus sp. nov. (SICAUCC 25-0059, SICAUCC 25-0060) formed a well-supported group containing M. raciborskii, M. simaoensis, M. hainanensis, M. puerensis, and M. pseudothanathonensis, but clustered in separate clades.
Phylogenetic tree based on maximum likelihood analysis of the combined LSU, rpb1, and tef1-α sequences within the Clavicipitaceae. Numbers above the nodes indicate MLBS values (≥ 60%, left) and BIPP values (≥ 0.95, right), with lower values denoted as “–”. The tree is rooted to Purpureocillium lilacinum (CBS 431.87, CBS 284.36). Ex-type strains are highlighted in bold, and isolates from the present study are shown in red.
Phylogenetic analysis of the genus Podonectria
To infer the relationships of the Podonectria-like taxa, a combined dataset of ITS, LSU, SSU, tef1-α, and rpb2 sequences was used to construct the phylogenetic tree. Phylogenetic analyses were conducted using a combined five-gene dataset comprising 52 taxa, with the tree rooted using Tubeufia javanica (MFLUCC 12-0545) and T. chiangmaiensis (MFLUCC 11-0514). The alignment contained 5,721 characters (ITS = 1–769, LSU = 770–1,744, rpb2 = 1,745–2,882, SSU = 2,883–4,301, tef1-α = 4,302–5,794), including gaps. The best-scoring tree with a final likelihood value of –41,343.562028 is presented. The matrix had 2,466 distinct alignment patterns, with 46.25% undetermined characters or gaps. The gamma distribution shape parameter α = 0.247235, and the tree length = 3.864868. The Bayesian analysis resulted in 4,602 trees after 1,000,000 generations, from which 3,452 were used for calculating posterior probabilities after the first 25% of trees, representing the burn-in phase, were discarded. Phylogenetic trees generated from ML and BI analyses were similar in overall topologies. The phylogeny derived from the combined sequence data analysis indicates that the novel strains form an independent lineage within the Podonectriaceae (Fig. 3). Based on the phylogenetic tree, our isolates clustered in a clade with species of Podonectria.
Phylogenetic tree based on maximum likelihood analysis of the combined ITS, LSU, SSU, tef1-α, and rpb2 sequences within the Pleosporales. Numbers above the nodes represent MLBS values (≥ 60%, left) and BIPP values (≥ 0.95, right), with lower values denoted as “–”. The tree is rooted to Tubeufia javanica (MFLUCC 12-0545) and T. chiangmaiensis (MFLUCC 11-0514). Ex-type strains are highlighted in bold, and isolates from the present study are shown in red.
Taxonomy
Cladosporium
kuwanaspidis
Taxon classificationAnimaliaCladosporialesCladosporiaceae
X.L. Xu & C.L. Yang sp. nov.
C57536B9-493B-5E7F-B279-114DAE7312BB
MB 858369
Etymology.
In reference to the generic name for the associated scale insect (Kuwanaspis howardi).
Diagnosis.
Similar to Cladosporium perangustum in having conidiophores of comparable size, but C. kuwanaspidis differs by its unbranched conidiophores and larger ramoconidia.
Type.
CHINA • Sichuan Province, Meishan City, Hongya County. Infected scale insects (Kuwanaspis howardi) were found on the culms of bamboo (Pleioblastus amarus (Keng) P. C. Keng), 29°41.88'N, 103°14.04'E, alt. 540 m, 13 Mar. 2021, C.L. Yang, YCL202103004 (SICAU 25-0082 – holotype preserved in the Herbarium of Sichuan Agricultural University; living culture SICAUCC 25-0063 – ex-holotype stored in the Culture Collection in Sichuan Agricultural University).
Description.
Parasitic on scale insect from Pleioblastus amarus (Poaceae). Sexual morph: Not observed. Asexual morph: Hyphomycetous. Mycelium superficial and immersed, with abundant sporulation on the surface of scale insect. Conidiophores erect, fasciculate, usually macronematous, cylindrical, subnodulose or nodulose, geniculate, septate, unbranched, pale brown to brown, slightly roughened to verruculose, thick-walled, and pronounced loci, 45–120 × 3.5–6 μm. Conidiogenous cells terminal or intercalary, cylindrical, sometimes sinuous, proliferation sympodial, 14–25(–80) × 2.5–6.5 μm, conidiogenous loci at the apex (2–5) or in lateral shoulders (0–2). Ramoconidia olive to brown, septate or aseptate, ellipsoidal to subcylindrical, smooth- and thick-walled, 6–13.5 × 3–5.5 μm. Secondary ramoconidia oblong, pale brown, 0–1 septate, 2–4 distal hila, 5.5–9 × 2.5–5 μm. Conidia numerous, catenate, forming short branched chains, aseptate, olive to brown, smooth- and thin- walled, ellipsoid-ovoid, obovoid, 2–7.5 × 2–4 μm. Intercalary conidia limoniform, oval to ellipsoid, with hila protuberant, 3.5–7.5 × 2.5–4 μm. Terminal conidia globose to ellipsoid, sometimes hila evident, 2–4 × 2–3.5 μm. Microcyclic conidiogenesis absent. In vitro on SNA: Mycelium abundant, submerged, overgrowing whole culture dishes, hyphae straight to slightly sinuous, septate and branched, olive to brown, and thick-walled, 2–3.5 µm wide. Conidiophores erect, occasionally branched, brown, thick-walled, 33–118 × 2.5–4 µm. Ramoconidia olive to brown, narrowly ellipsoid to cylindrical-oblong, subcylindrical, septate or aseptate, smooth- and thick-walled, 6–17.5 (–20) × 2–5 µm. Conidia in simple and branched acropetal chains, light olive, aseptate, smooth- and thin-walled, ellipsoidovoid, obovoid, fusiform, 3–6.5 × 2–4 µm. Intercalary conidia oval to ellipsoid, with hila protuberant, 3.5–6.5 × 2.5–4 μm. Terminal conidia globose to ellipsoid, 2.5–4 × 2–4 μm.
Cladosporium kuwanaspidis. A–C. Symptoms and appearance of colonies observed on Kuwanaspis howardi. D–G. Conidiophores. H. Ramoconidia (white arrows), secondary ramoconidia (red arrows), and terminal conidia (yellow arrows). I–K. Colonies on MEA, PDA, and SNA for 7 days. Scale bars: 200 µm (B, C); 10 µm (D–H).
Culture characteristics.
Conidia of all types can germinate within 12 h in sterile distilled water, with germination tubes forming from any part of the conidial body. Colonies on MEA attaining 40–50 mm diameter, after 1 week at 25 °C, greyish-green to greyish toward regular margins, velvety, sporulation profuse, radially furrowed, aerial mycelium abundant, reverse dark green. Colonies on PDA attaining 35–40 mm diameter, after 1 week at 25 °C, grey-olivaceous to iron grey, pale greenish grey toward regular margins, reverse dark green. Colonies on SNA attaining 20–30 mm diameter, after 1 week at 25 °C, smoke-grey to olivaceous-grey, aerial mycelium loose, reverse olivaceous to brown.
Host.
Kuwanaspis howardi (Diaspididae).
Habitat.
Subtropical evergreen broad-leaved forests, particularly bamboo forests, are common habitats. Kuwanaspis howardi often causes damage in the lower canopy of bamboo forests. Occasionally, Cladosporium kuwanaspidis can be observed on bamboo culms. Scale insects are more easily found in shaded and sheltered areas.
Distribution.
China, Sichuan Province, Meishan City.
Material examined.
CHINA • Sichuan Province, Meishan City, Hongya County. Infected scale insects (Kuwanaspis howardi) were found on the culms of bamboo (Pleioblastus amarus), 29°41.88'N, 103°14.04'E, alt. 540 m, 9 Mar. 2021, C.L. Yang, YCL202103004 (living culture SICAUCC 25-0063); • ibid. YCL202103004-1 (SICAU 25-0083, living culture SICAUCC 25-0064).
Notes.
The ITS base-pair comparison between Cladosporium kuwanaspidis (SICAUCC 25-0063) and the phylogenetically affiliated ex-type culture of C. perangustum (CBS 125996) shows no nucleotide differences. However, nucleotide differences in the act and tef1-α regions are 1.86% (4 bp, 0 gap) and 4.01% (15 bp, 2 gaps), respectively. Morphologically, C. perangustum has larger ramoconidia compared to those observed in C. kuwanaspidis on both the host and SNA medium (25–45 µm vs. 6–13.5 µm and 6–20 µm, respectively). In addition, compared to the length of intercalary conidia (4–19 µm) observed in C. perangustum (CBS 125996), our isolates exhibit shorter intercalary conidia on both the host and PDA medium (3.5–7.5 µm and 3.5–6.5 µm, respectively) (Bensch et al. 2010).
Cladosporium
guizhouense
Taxon classificationAnimaliaCladosporialesCladosporiaceae
S.Y. Wang, Yong Wang bis & Y. Li, MycoKeys 91: 151–168 (2022) sp. nov.
ED7038F0-BC36-58B0-BC5E-62217B81C6E3
MB 842407
Description.
Parasitic on aphids from Telosma cordata (Burm. f.) Merr. (Apocynaceae). Sexual morph: Not observed. Asexual morph: Hyphomycetous.
Mycelium superficial and immersed, with abundant sporulation on the surface of aphids. Conidiophores erect, fasciculate, macronematous, cylindrical, subnodulose or nodulose, geniculate, septate, branched, pale brown, slightly roughened to verruculose, thick-walled, and pronounced loci, 111–367 × 3.5–5 μm. Conidiogenous cells integrated, terminal or intercalary, cylindrical, cylindrical-oblong, sometimes geniculate, 20–65 × 3–6 μm, conidiogenous loci at the apex (2–5) or in lateral shoulders (0–4). Ramoconidia olive-green, 0–4-septate, ellipsoidal to subcylindrical, smooth- and thick-walled, 10–26 × 3.5–7.5 μm. Secondary ramoconidia pale brown, oblong, oblong-ellipsoid, 0–1septate, 1–4 distal hila, 8.5–13.5 × 3.5–7.5 μm. Conidia numerous, catenate, forming short branched chains, aseptate, olive to light olive, smooth- and thin-walled, variable in size and shape, ellipsoid-ovoid, obovoid, and fusiform, 4–10 × 3–6.5 μm. Intercalary conidia aseptate, olive to light olive, ellipsoid-ovoid, fusiform, 6–10 × 4–6.5 μm. Terminal conidia aseptate, olive to light olive, obovoid, 4–6.5 × 3–5.5 μm. Microcyclic conidiogenesis absent. In vitro on SNA: Mycelium abundant, submerged, overgrowing whole culture dishes, hyphae straight to slightly sinuous, septate and branched, light olive-green to olive-green, and thick-walled, 1.5–4.5 µm wide. Conidiophores erect, branched, light olive-green, thick-walled, 43–163 × 2.5–4 µm. Ramoconidia pale olivegreen, narrowly ellipsoid to cylindrical-oblong, subcylindrical, septate or aseptate, smooth- and thick-walled, 6–27 × 3–5 µm. Conidia in simple and branched acropetal chains, light olive, aseptate, smooth- and thin-walled, variable in size and shape, ellipsoid-ovoid, obovoid, fusiform, 3–8 × 2.5–4 µm. Intercalary conidia light olive, aseptate, smooth- and thin-walled, mostly ellipsoid, 3.5–8 × 2.5–4 µm. Terminal conidia light olive, aseptate, smooth- and thin-walled, mostly ovoid, 3–4.5 × 2.5–3.5 µm.
Cladosporium guizhouense. A. Symptoms observed on host. B, C. Appearance of colonies on aphids. D–H. Conidiophores. I. Ramoconidia (white arrows), secondary ramoconidia (red arrows), and terminal conidia (yellow arrows). J–M. Colonies on MEA, PDA, OA, and SNA for 7 days. Scale bars: 1000 µm (B); 200 µm (C); 20 µm (D–I).
Culture characteristics.
All conidia can germinate within 12 h in sterile distilled water, with tubes forming from any part. Colonies on MEA attaining 35–45 mm diameter, after 1 week at 25 °C, dark green, white to greyish toward regular margins, velvety, sporulation profuse, radially furrowed, aerial mycelium abundant, reverse dark green. Colonies on PDA attaining 30–40 mm diameter, after 1 week at 25 °C, greyishgreen, greyish toward irregular margins, velvety, sporulation profuse, radially furrowed, aerial mycelium abundant, reverse dark green. Colonies on OA attaining 30–40 mm diameter, after 1 week at 25 °C, greyish olivaceous, white toward irregular margins, sporulation profuse, reverse olivaceous. Colonies on SNA attaining 25–30 mm diameter, after 1 week at 25 °C, olivaceous, flat, white toward regular and wide margins, aerial mycelium loose, reverse olivaceous.
Host.
Aphids (Aphididae), leaves of plants, and uredinia of Hemileia vastatrix (Pucciniaceae).
Habitat.
In tropical and subtropical regions, it can infect insects, act saprophytically on plant tissues, and function as a fungicolous fungus on the uredinia of rust. During the middle and late stages of aphid damage, numerous aphids die on the underside of plant leaves, becoming enveloped in mycelial tissue.
Distribution.
China, Sichuan Province, Guangan City. China, Guizhou Province, Guiyang City. Ethiopia, Oromia Region, Illubabor Gore. Brazil, Minas Gerais, Viçosa.
Material examined.
CHINA • Sichuan Province, Guangan City, Yuechi County. Infected aphids were found on the underside of the leaves of Telosma cordata, 30°44.18"N, 106°30.80"E, alt. 480 m, 31 Jan. 2020, X.L. Xu, XXL202001003 (SICAU 25-0076, living culture SICAUCC 25-0057); • ibid. XXL202001003-1 (SICAU 25-0077, living culture SICAUCC 25-0058).
Notes.
Phylogenetically, our collections grouped with isolates of Cladosporium guizhouense within the C. cladosporioides complex, showing strong statistical support (92% MLBS, 0.97 BIPP) (Fig. 1). A total of 6 bp nucleotide differences were observed between our isolate SICAUCC 25-0057 and the ex-type culture of C. guizhouense (GUCC 401.7), with 0 bp in ITS, 2 bp in act, and 4 bp in tef1-α (Wang et al. 2022b). Morphologically, our isolate is fully consistent with C. guizhouense, showing no noticeable differences (Wang et al. 2022b; Pereira et al. 2024).
Moelleriella
eucalypti
Taxon classificationAnimaliaHypocrealesClavicipitaceae
X.L. Xu, Feng Liu & C.L. Yang sp. nov.
D19B1D61-701B-506C-BBEF-3202B64D2F12
MB 858370
Etymology.
In reference to the generic name of host plants.
Diagnosis.
Similar to Moelleriella sinensis in having somewhat similar stromata, M. eucalypti differs by having longer and wider paraphyses, shorter and wider conidiogenous cells, and wider conidia.
Type.
CHINA • Sichuan Province, Dazhou City, Heibaoshan National Forest Park. Infected scale insects were found on the underside of leaves of Eucalyptus sp. (Myrtaceae), 31°55.30'N, 107°47.75'E, alt. 690 m, 21 Jul. 2022, Feng Liu, LF202207001A (SICAU 25-0072 – holotype preserved in the Herbarium of Sichuan Agricultural University).
Description.
Parasitic on scale insects found on eucalyptus leaves. Sexual morph: Not observed. Asexual morph: Coelomycetous. Stromata yellow to dark orange in fresh specimens, pale yellow to white in old, entirely covering the insect hosts, thickened pulvinate, umbonate to hemispheric, cottony, sessile, globose to subglobose, tubercules on the surface, 1.5–2.5 mm in diameter, 1–2 mm in high. Conidiomata orifice scattered or circularly arranged, oval or elongate flask shaped, narrow orifices, 120–460 × 80–155 μm. Phialides hyaline, stick-shaped to cylindrical, 10–18 × 1–2.5 μm. Conidia hyaline, fusiform, yellow conidial masses, 8–12 × 1.5–3 μm. Paraphyses present, hyaline, filiform, 90–180 × 1.0–1.6 μm.
Moelleriella eucalypti. A. Fungus-infected scale insect on the underside of leaves. B–D. Stromata containing conidiomata with conidial masses. E–I. Section of stroma showing conidioma. J–M. Phialides and conidia at the tips with paraphyses. N. Conidia. Scale bars: 500 mm (B–D); 200 μm (E, F); 50 μm (H, I); 10 μm (J, K).
Culture characteristics.
No germination was observed due to the specimens being dried and the culture not being obtained.
Host.
Scale insects (Coccidae).
Habitat.
Subtropical monsoon evergreen broad-leaf forest. Scale insects were infected on the underside of eucalyptus leaves.
Distribution.
China, Sichuan Province, Dazhou City.
Material examined.
CHINA • Sichuan Province, Dazhou City, Heibaoshan National Forest Park. Infected scale insects were found on the underside of leaves of Eucalyptus sp., 31°55.30'N, 107°47.75'E, alt. 690 m, 21 Jul. 2022, Feng Liu, LF202207001A (SICAU 25-0072); • ibid. LF202207001B (SICAU 25-0073); • ibid. LF202207001C (SICAU 25-0074); • ibid. LF202207001D (SICAU 25-0075).
Notes.
Phylogenetically, our collections are closely related to Moelleriella sinensis, showing few sequence differences with the type strain CGMCC3.18911, specifically 1 bp in LSU (0.12%, 0 gap) and 8 bp in rpb1 (1.10%, 0 gap), while the tef1-α sequence is invalid. Moelleriella sinensis was first described on whitefly nymphs (Hemiptera) and later found on scale insects (Coccidae) in Thailand (Chen et al. 2020; Khonsanit et al. 2021). The described specimens of M. sinensis are primarily characterized by flat to umbonate, pale yellow pulvinate stromata and fusiform conidia. However, our collections differ from M. sinensis in having pulvinate stromata that are yellow to dark orange when fresh, pale yellow to white when old, and thicker in appearance. Additionally, they have longer and wider paraphyses (90–180 × 1.0–1.6 μm vs. 43.2–68.9 × 0.6–0.8 μm), shorter and wider conidiogenous cells (10–18 × 1–2.5 μm vs. up to 30 μm × 0.8–1.3 μm), and wider conidia (1.5–3 μm vs. 1.3–1.8 μm). Hence, we introduce M. eucalypti as a new species, based on the distinct morphological differences observed.
Moelleriella
boehmeriae
Taxon classificationAnimaliaHypocrealesClavicipitaceae
X.L. Xu & C.L. Yang sp. nov.
D057A820-33B7-5F5C-A339-0D4170E17819
MB 858372
Etymology.
In reference to the generic name of host plants.
Diagnosis.
Similar to Moelleriella jinuoana in having yellow to orange, globose stromata with a narrow hypothallus, M. boehmeriae differs by its slower growth on PDA, unique colony characteristics, and wider conidia.
Type.
CHINA • Sichuan Province, Leshan City, Muchuan County. Infected scale insects were found on the stems of Boehmeria spicata (Thunb.) Thunb., 28°47.91'N, 103°55.63'E, alt. 900 m, 12 Mar. 2021, C.L. Yang, YCL202103003 (SICAU 25-0080 – holotype preserved in the Herbarium of Sichuan Agricultural University; living culture SICAUCC 25-0061 – ex-holotype stored in the Culture Collection in Sichuan Agricultural University).
Description.
Parasitic on scale insect from Boehmeria spicata (Urticaceae). Sexual morph: Not observed. Asexual morph: Coelomycetous. Stromata yellow to orange in fresh specimens, entirely covering the insect hosts, thickened pulvinate, globose, tubercules on the surface, closely aggregated, some with narrow hypothallus, 0.8–2.5 mm diameter. Hyphae of stromata form compact textura epidermoidea. Conidiomata simple depressions of surface, round or irregular shape, no mature conidiomata observed.
Moelleriella boehmeriae. A, B. Immature stromata over scale insects. C. Conidioma formed in PDA. D. Phialides with conidia at the tips. E–G. Phialides with developing conidia. H. Conidia. I. Germinating conidia. J, K. Colonies obverse and reverse on PDA at 25 °C after 3 weeks. L. Conidial mass on PDA. Scale bars: 1000 µm (B); 50 µm (C); 20 µm (D); 10 µm (E–I).
Culture characteristics.
The conidia germinate within 12 h in sterile distilled water, with germ tubes extending from both ends of the conidia. Colonies on PDA slowgrowing, attaining a diameter of 5–7 mm in 28 days at 25 °C. Colonies compact pulvinate, surface velutinous, pale yellow to yellow. Conidial masses usually abundant, yellow. Reverse of colony pale orange. Conidiomata U-shaped, subcircular to circular. Conidia unicellular, hyaline, smooth, fusoid with rounded ends, 9.5–12.5 × 3–4 µm. No paraphyses were observed.
Host.
Scale insects (Coccidae).
Habitat.
Subtropical monsoon evergreen broad-leaf forests serve as the habitat for Moelleriella boehmeriae, which can infect scale insects found on the stems of Boehmeria spicata. Infections are typically observed at lower heights, close to the ground.
Distribution.
China, Sichuan Province, Leshan City.
Material examined.
CHINA • Sichuan Province, Leshan City, Muchuan County. Infected scale insects were found on the stems of Boehmeria spicata, 28°47.91'N, 103°55.63'E, alt. 900 m, 12 Mar. 2021, C.L. Yang, YCL202103003 (SICAU 25-0080, living culture SICAUCC 25-0061); • ibid. YCL202103003-1 (SICAU 25-0081, living culture SICAUCC 25-0062).
Notes.
Phylogenetic analysis revealed that Moelleriella boehmeriae clusters within the Globose clade and is closely related to M. jinuoana (Fig. 2), sharing characteristics such as yellow to orange, globose stromata with a narrow hypothallus (Wang et al. 2024). Although detailed morphological comparisons were limited due to the absence of mature conidiomata in our specimen, M. boehmeriae differs by its slower growth on PDA and distinct colony characteristics. The colonies of M. boehmeriae are yellow on the front and light orange on the reverse, whereas those of M. jinuoana are pale orange to orange on the front and brownish on the reverse. Additionally, M. boehmeriae has wider conidia (3–4 µm) observed in culture compared to M. jinuoana, which has conidia measuring 2.1–2.9 µm on the substrate. In addition, M. boehmeriae differs from the type strain M. jinuoana (YHH MJBP2309031) by 16 bp (1.76%, 0 gap) in the tef1α region, 2 bp (0.28%, 0 gap) in the rpb1 region, and 4 bp (0.47%, 2 gaps) in the LSU region, respectively.
Moelleriella
cinnamomum
Taxon classificationAnimaliaHypocrealesClavicipitaceae
X.L. Xu & C.L. Yang sp. nov.
EA4791CE-91B4-574B-98EC-EE4590D1454F
MB 858373
Etymology.
In reference to the generic name of host plants.
Diagnosis.
Similar to Moelleriella simaoensis, M. puerensis, M. raciborskii, and M. citrus, M. cinnamomum differs by having red-orange, thickened pulvinate stromata, longer conidia and paraphyses, and the absence of a hypothallus surrounding the stroma.
Type.
CHINA • Sichuan Province, Chengdu City, Chongzhou County. Infected scale insects were found on the underside of leaves of Cinnamomum cassia (L.) D. Don., 30°48.82'N, 103°31.52'E, alt. 851 m, 1 May 2021, C.L. Yang, YCL202105001 (SICAU 25-0086 – holotype preserved in the Herbarium of Sichuan Agricultural University; living culture SICAUCC 25-0067 – ex-holotype stored in the Culture Collection in Sichuan Agricultural University).
Description.
Parasitic on scale insects found on the underside of leaves of Cinnamomum cassia (Lauraceae). Sexual morph: Not observed. Asexual morph: Coelomycetous. Stromata red-orange in fresh specimens, mostly solitary, sometimes gregarious, globose to subglobose, thickened pulvinate up to 1 mm, 0.9–2.8 mm in diameter. Conidiomata aggregated in the center of the stroma and widely open, simple depressions of the surface without distinct rims. In section, conidioma flask-shaped or irregular, shallow. Conidial masses orange. Conidia hyaline, smooth, one-celled, fusoid, with acute ends, 13–16.5 × 2–3 µm. Paraphyses present, linear, filiform, up to 102 µm long.
Moelleriella cinnamomum. A, B. Stromata over scale insects on the underside of leaves. C. Section of stroma showing conidioma. D. Phialides and conidia at the tips with paraphyses. E. Paraphyses with phialides bearing developing conidia. F. Conidia. G. Germinating conidia. H, I. Colonies obverse and reverse on PDA at 25 °C after 3 weeks. J. Conidial mass on PDA. Scale bars: 1000 µm (B); 500 µm (C); 20 µm (D); 10 µm (E–G).
Culture characteristics.
Conidia germinate in sterile water within 12 h, with germination tubes typically growing laterally from both ends of the spore and occasionally from the middle. Colonies on PDA at 25 °C attaining 15 mm diameter in 20 days. Colonies pale yellow and formed compact pulvinate with abundant slimy masses. Conidial masses light yellow. Colonies reverse dark yellow with pale yellow margins. Conidia 9–15 × 1.8–2.8 µm. Paraphyses rare.
Host.
Scale insects (Coccidae).
Habitat.
Subtropical monsoon evergreen broad-leaf forests serve as the habitat for Moelleriella cinnamomum, which can infect scale insects found on the underside of Cinnamomum cassia leaves. Infections are typically observed throughout the trees, particularly in the lower canopy.
Distribution.
China, Sichuan Province, Chengdu City.
Material examined.
CHINA • Sichuan Province, Chengdu City, Chongzhou County. Infected scale insects were found on the underside of leaves of Cinnamomum cassia, 30°48'49.57"N, 103°31'31.19"E, alt. 851 m, 1 May 2021, C.L. Yang, YCL202105001 (SICAU 25-0086, living culture SICAUCC 25-0067); • ibid. YCL202105002 (SICAU 25-0087, living culture SICAUCC 25-0068).
Notes.
Our collections were nested in the Moelleriella clade related to M. citrus, M. raciborskii, M. simaoensis, M. hainanensis, M. puerensis, and M. pseudothanathonensis (Fig. 2). Morphologically, M. cinnamomum is distinct from the phylogenetically related six species, with red-orange and thickened pulvinate, while the other species are flattened and more pale in color (Liu et al. 2006; Chaverri et al. 2008; Wang et al. 2022c, 2025; Yang et al. 2023a). Moelleriella raciborskii, M. simaoensis, and M. puerensis can co-occur in the same stromata with both sexual and asexual morphs, whereas only the asexual morph of M. cinnamomum has been observed in stromatal tissue. Moelleriella hainanensis differs from M. cinnamomum by having larger stromata (3–4 mm vs. 0.9–2.8 mm), and along with M. raciborskii and M. citrus, it also differs by exhibiting a light-colored hypothallus surrounding the stroma. Conidia in M. cinnamomum (13–16.5 µm) are longer than those of M. simaoensis (8.8–14 µm), M. puerensis (9.7–13.4 µm), M. pseudothanathonensis (10–12.5 µm), and M. raciborskii (11–14 µm). Moreover, M. cinnamomum has longer paraphyses (up to 102 µm) than M. citrus (85–100 µm), M. simaoensis (up to 95 µm) and M. raciborskii (40–70 µm).
Moelleriella
citrus
Taxon classificationAnimaliaHypocrealesClavicipitaceae
X.L. Xu & C.L. Yang sp. nov.
8D509E83-9215-5AB0-B171-097096DB3BC4
MB 858374
Etymology.
In reference to the generic name of host plants.
Diagnosis.
Similar to Moelleriella raciborskii in having pale yellow, thin, pulvinate stromata, M. citrus differs by its longer paraphyses and wider conidia.
Type.
CHINA • Sichuan Province, Chengdu City, Wenjiang District. Infected scale insects were found on the underside of leaves of Citrus sp., 30°42.40'N, 103°51.05'E, alt. 590 m, 10 Oct. 2020, X.L. Xu, XXL202010001 (SICAU 25-0078 – holotype preserved in the Herbarium of Sichuan Agricultural University; living culture SICAUCC 25-0059 – ex-holotype stored in the Culture Collection in Sichuan Agricultural University).
Description.
Parasitic on scale insect from citrus leaves (Rutaceae). Sexual morph: Not observed. Asexual morph: Coelomycetous. Stromata orange in fresh specimens, globose to subglobose, flattened pulvinate, 1.8–3.0 mm in diameter, surrounded by a hyaline hypothallus up to 1.6 mm wide and covered with confluent conidial masses of orange-yellow. Conidiomata simple and aggregated in the center of the stroma, and widely opened. Conidial masses orange. Conidia hyaline, smooth, one-celled, fusoid, with acute ends, produced in copious slime, 12–18.5 × 2.3–3.2 µm. Paraphyses present, arising from the hymenium of the conidioma, filiform, tapering at the apices, up to 85–100 µm long.
Moelleriella citrus. A, B. Fungus-infected scale insect on the underside of leaves. C. Section of stroma showing conidioma. D. Phialides and conidia at the tips with paraphyses. E. Conidia. F. Germinating conidia. G, H. Colonies obverse and reverse on PDA at 25 °C after 3 weeks. Scale bars: 1000 µm (A); 500 µm (B); 50 µm (C); 10 µm (D–F).
Culture characteristics.
Conidia germinate in sterile water within 12 h, with germination tubes typically growing laterally from both ends of the spore. Colonies on PDA at 25 °C attaining 25 mm diameter in 20 days. Colonies pale yellow and formed compact pulvinate with abundant slimy masses. Conidial masses light yellow, the colony reverse dark yellow, and the margins pale yellow. Conidia hyaline, smooth, onecelled, fusoid, 12–15.5 × 2.0–2.8 µm. Paraphyses rare.
Host.
Scale insects (Coccidae).
Habitat.
Subtropical monsoon evergreen broad-leaf forests serve as the habitat for Moelleriella citrus, which infects scale insects found on the underside of Citrus sp. leaves. Infections are typically observed throughout the trees.
Distribution.
China, Sichuan Province, Chengdu City.
Material examined.
CHINA • Sichuan Province, Chengdu City, Wenjiang District. Infected scale insects were found on the underside of leaves of Citrus sp., 30°42.40'N, 103°51.05'E, alt. 590 m, 10 Oct. 2020, X.L. Xu, XXL202010001 (SICAU 25-0078, living culture SICAUCC 25-0059); • ibid. XXL202010001-1 (SICAU 25-0079, living culture SICAUCC 25-0060).
Notes.
Moelleriella raciborskii is accepted as the teleomorph of Aschersonia placenta, which was previously linked to Hypocrella raciborskii (Chaverri et al. 2008). Morphologically, M. citrus is closely related to M. raciborskii, sharing characteristics such as a pale yellow, thin, pulvinate stroma, ovoid or subglobose tubercle processes developing on the periphery of the stroma, and orange ostioles. However, the paraphyses show differences in length, with our collection measuring 85–100 µm compared to 40–70 µm in the previous species, and our collection has larger conidia (12–18.5 × 2.3–3.2 µm vs. 11–14 × 1.5–2 µm) (Liu et al. 2006).
Podonectria
multiarmata
Taxon classificationAnimaliaPleosporalesPodonectriaceae
X.L. Xu & C.L. Yang sp. nov.
5B0521AE-FEA9-599F-B7AC-09D233734A3A
MB 85837
Etymology.
Refers to the species having conidia bearing multiple arm-like extensions.
Diagnosis.
Similar to species of Podonectria, P. multiarmata differs notably in sporulation and conidial morphology, particularly in the extension patterns, size, and septation of the conidial arms.
Type.
CHINA • Sichuan Province, Meishan City, Hongya County. Infected scale insects (Kuwanaspis howardi) were found on the culms of bamboo (Pleioblastus amarus), 29°41.88'N, 103°14.04'E, alt. 540 m, 13 Mar. 2021, C.L. Yang, YCL202103005 (SICAU 25-0084 – holotype preserved in the Herbarium of Sichuan Agricultural University; living culture SICAUCC 25-0065 – ex-holotype stored in the Culture Collection in Sichuan Agricultural University).
Description.
Habitat associated with scale insects Kuwanaspis howardi on Pleioblastus amarus (Poaceae). Sexual morph: Not observed. Asexual morph: Hyphomycetes. Colonies surround the scale insects, effuse, white, and diffuse outward to produce thin layers of hyphae. Mycelia branched, septate, 1–3 µm wide. Conidiophores inconspicuous, mononematous, short, straight or slightly curved, mostly reduced to conidiogenous cells. Conidiogenous cells subglobose to globose, acrogenous, determinate hyaline, with obviously conidiogenous loci, 4–6 × 4–7 µm. Conidia usually with two and three arms, V-shaped, Y-shaped or T-shaped, occasionally four arms, smooth, hyaline, each arm varies in length and sharply divergent, slightly constricted at the septa, straight or curved, and tapering toward the apical cell, 1–4septate for each arm, 20–40 (–50) µm long, 3.5–6 wide at the base, 1.5–3 wide near the apex. All arms arise from a bigger basal cell, measuring 6–8 µm wide.
Podonectria multiarmata. A–D. Views of colonies of scale insects. E, F. Conidiogenous cells with loci at the apex (yellow arrows). G. Conidiogenous cell with developing conidium. H–M. Conidia. N. Germinating conidium. O, P. Upper and reverse views of cultures on PDA after 3 weeks. Scale bars: 10 μm (E–M); 20 μm (N).
Culture characteristics.
Conidia germinate on PDA within 12 h, with the germination tube usually growing from the tip of the arm. Colonies grow slowly on PDA, reaching 2.5 cm in diameter after 20 days at 25 °C, flat, circular, white, gradually becoming light fleshy pink, and the back of colonies yellow.
Host.
Kuwanaspis howardi (Coccidae).
Habitat.
Subtropical monsoon evergreen broad-leaf forests serve as the habitat for Podonectria multiarmata, which infects scale insects found on the culms of bamboo. Infections are typically observed in moist, shaded environments.
Distribution.
China, Sichuan Province, Meishan City.
Material examined.
CHINA • Sichuan Province, Meishan City, Hongya County. Infected scale insects (Kuwanaspis howardi) were found on the culms of Pleioblastus amarus, 29°41.88'N, 103°14.04'E, alt. 540 m, 15 Mar. 2021, C.L. Yang, YCL202103005 (SICAU 25-0084, living culture SICAUCC 25-0065); • ibid. YCL202103005-1 (SICAU 25-0085, living culture SICAUCC 25-0066).
Notes.
Our collections clustered phylogenetically with species of Podonectria based on combined ITS, LSU, SSU, rpb2, and tef1-α sequence data (Fig. 3) (100% MLBS, 1.00 BIPP). Nucleotide BLAST sequence showed distinguishing identities with those related species P. sichuanensis (SICAU 16-0003), P. coccicola (pAak), P. novae-zelandiae (SICAUCC 21-0004), and P. kuwanaspidis (SICAUCC 21-0002) in ITS (90.38%, 88.45%, 89.89%, 88.26%), LSU (96.77%, 98.26%, 97.92%, 98.62%), SSU (99.62%, -, 99.81%, 100%), tef1-a (95.34%, -, 94.35%, 95.34%), and rpb2 (93.20%, -, 90.74%, 90.39%), respectively. Podonectria multiarmata resembles other Podonectria species in its asexual state, producing conidia with arms that are joined at a basal cell (Xu et al. 2021a). However, it differs in the length and divergence angle of each arm, and it also produces T-type and Y-type conidia with radiating divergent arms.
Discussion
Hypocreales (Ascomycota) and Entomophthoromycotina (Zoopagomycota) harbor the most common insect-pathogenic fungi (Hong et al. 2024), while insects in the Coccidae and Aleyrodidae exhibit the greatest documented diversity of fungal pathogens (Humber 2008). In this study, we identified seven species of entomopathogenic fungi associated with scale insects and aphids, belonging to Cladosporium, Moelleriella, and Podonectria, based on morphology and phylogeny. These findings provide valuable insights to support the further development and application of effective biological control strategies against pests.
Cladosporium is a genus in the family Cladosporiaceae of Dothideomycetes (Abdollahzadeh et al. 2020), isolated from a wide range of substrates and known as saprobes, endophytes, plant pathogens, and hyperparasites of other fungi and etiologic agents in vertebrate hosts, including humans (El-Morsy 2000; Abdel-Baky and Abdel-Salam 2003; Heuchert et al. 2005; Levetin and Dorsey 2006; Bensch et al. 2012; Sandoval-Denis et al. 2016; Prasannath et al. 2021a, 2021b; Xu et al. 2022; Pereira et al. 2024). The classification of Cladosporium is problematic due to the infrequency of the teleomorphic stage and the similarity in conidial structures. Mycologists have resolved the complexes of C. cladosporioides, C. herbarum, and C. sphaerospermum with the help of biomolecular tools. The occurrence of Cladosporium species within these three complexes, in association with insects, was summarized from the available literature by Nicoletti et al. (2024). In that list, about 46% of the entries were not identified due to the challenging classification of Cladosporium, and the insect-associated species were predominantly linked to Hemiptera (28.2%) and Coleoptera (27.6%). Cladosporium guizhouense seems to have a geographical distribution with a broad host range, based on records from leaves of Eucommia ulmoides (Eucommiaceae) (Wang et al. 2022b) and Citrus reticulata (Rutaceae) (Yang et al. 2023b). In addition, three isolates of C. guizhouense were obtained from uredinia of Austropuccinia psidii on leaves of Psidium guajava and Syzygium jambos in Brazil (Silva et al. 2023) and uredinia of Hemileia vastatrix on leaves of Coffea arabica from Africa (Pereira et al. 2024). This is the first report of C. guizhouense from aphids on Telosma cordata (Apocynaceae). The scale insect Kuwanaspis howardi is commonly distributed and harmful to bamboo. In our previous study, two Podonectria species and a Microcera species associated with native bamboo plants were discovered (Xu et al. 2021a). Cladosporium kuwanaspidis is the second entomopathogenic species found on bamboo (Pleioblastus amarus) in our investigations. As entomopathogenic C. cladosporioides is a potential candidate for biocontrol against whiteflies and aphids (Islam et al. 2019; Nicoletti et al. 2024), and as endophytic Cladosporium sp. has demonstrated insecticidal activity of α-glucosidase inhibitors against Spodoptera litura in vitro (Singh et al. 2015), more Cladosporium species need to be investigated as potential agents of biocontrol.
The genus Moelleriella (Ascomycota, Hypocreales, Clavicipitaceae) was established to accommodate M. sulphurea (Bresadola 1896) and is characterized by globose to pyriform perithecia immersed in a brightly colored stroma with openings protruding from the stroma on scale insects (Coccidae, Homoptera) or whiteflies (Aleyrodidae, Homoptera), as well as multi-septate ascospores that disarticulate at the septa inside the ascus. In contrast, species of Hypocrella and the subsequently proposed genus Samuelsia, having Aschersonia-like anamorphs, have ascospores that remain whole (Petch 1921; Mains 1959; Kirk et al. 2001; Chaverri et al. 2008). The genus currently has about 52 records (https://www.speciesfungorum.org/Names/Names.asp, accessed on 4 November 2024), and species have been reported in Belize, Bolivia, Costa Rica, Côte d’Ivoire, China, Ecuador, Ghana, Guatemala, Guiana, Honduras, Jamaica, Mexico, Panama, the Philippines, Thailand, Trinidad, Venezuela, and Vietnam, frequently from Thailand, indicating it is more common in tropical regions (Liu et al. 2006; Chaverri et al. 2008; Qiu et al. 2009; Mongkolsamrit et al. 2011b, 2015; Li et al. 2016; Tibpromma et al. 2017; Chen et al. 2020; Yuan et al. 2020; Khonsanit et al. 2021). Moelleriella libera has been widely used due to its parasitism of large populations of whiteflies and scale insects in the wild (Zhang et al. 2018; Ingle et al. 2019; Qasim et al. 2020). In China, M. ochracea has previously been recorded from Fujian Province on homopteran cadavers (Qiu et al. 2009). In recent years, several species have been reported from the subtropical regions of Yunnan and Fujian provinces, viz. M. gracilispora on whitefly nymphs (Hemiptera) (Yuan et al. 2020), M. sinensis infecting whitefly nymphs (Hemiptera) (Chen et al. 2020), M. puerensis on whiteflies (Wang et al. 2022c), M. simaoensis on whiteflies (Yang et al. 2023a), and M. jinuoana and M. longzhuensis on scale insects and whiteflies (Wang et al. 2024). Based on our observation, four Moelleriella species were newly recorded on scale insects inhabiting Boehmeria spicata, Citrus sp., and Eucalyptus sp. in Sichuan Province. Moelleriella sinensis, M. cinnamomum, and M. citrus were accommodated in the Effuse clade (Chaverri et al. 2008; Wang et al. 2024), which is characterized by effuse to thin, pulvinate stromata with loose hyphal tissue, mostly having hypothalli. The new species M. boehmeriae was nested in the Globose clade (Chaverri et al. 2008), characterized by globose, darker stromata with compact tissue.
Podonectriaceae was proposed as a family in the Pleosporales to accommodate the genus Podonectria, which was confirmed by phylogenetic analyses (Dao et al. 2016; Yang et al. 2019b). Podonectria was traditionally recorded with a Tetracrium-like conidial stage (Petch 1921; Rossman 1978). Previous studies confirmed the link between the sexual morphs in Podonectria and the asexual morphs in Tetracrium with identical molecular sequences (Dao et al. 2015, 2016; Xu et al. 2021a). In addition, an asexual genus, Tetranacrium, typified by T. gramineum (Hudson and Sutton 1964), was recorded as an associated anamorph of Podonectria gahnia on scale insects. However, although the shape and development of conidia in the associated Tetranacrium are correlated with Tetracrium, the conidiomata in Tetranacrium are pycnidia, whereas in Tetracrium they are sporodochia. The relationship between Tetranacrium and Podonectria requires further phylogenetic and taxonomic studies with more samples. In this study, our collections had hyphomycetous anamorphs, which were consistent with Podonectria, identified based on the phylogenetic results of combined ITS, LSU, SSU, tef1-α, and rpb2 data. Most species of Podonectria are associated with armored scale insects on Citrus spp. Podonectria multiarmata is the third entomopathogenic species associated with Kuwanaspis howardi on bamboo (Pleioblastus amarus) in China.
Conclusion
Seven entomopathogenic fungi from Sichuan Province, China, are described, including six new species and one newly recorded species. These species are morphologically simple hyphomycetes or coelomycetes, with detailed identification based on morphological characteristics and phylogenetic analyses. These fungi warrant attention, particularly those associated with major agricultural and forestry pests. We predict that southwestern China harbors a rich diversity of entomopathogenic fungi, with many species yet to be discovered and evaluated. These fungi represent a crucial resource for future drug development and hold significant potential for pest management in agriculture and forestry.
Supplementary Material
XML Treatment for Cladosporium kuwanaspidis
XML Treatment for Cladosporium guizhouense
XML Treatment for Moelleriella eucalypti
XML Treatment for Moelleriella boehmeriae
XML Treatment for Moelleriella cinnamomum
XML Treatment for Moelleriella citrus
XML Treatment for Podonectria multiarmata
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abdel-Baky NF Abdel-Salam AH (2003) Natural incidence of Cladosporium spp. as a bio-control agent against whiteflies and aphids in Egypt.Journal of Applied Entomology 127: 228–235. 10.1046/j.1439-0418.2003.00662.x · doi ↗
- 2Abdollahzadeh J Groenewald JZ (2020) Evolution of lifestyles in Capnodiales.Studies in Mycology 95: 381–414. 10.1016/j.simyco.2020.02.00432855743 PMC 7426231 · doi ↗ · pubmed ↗
- 3Akutse KS Maniania NK (2013) Endophytic colonization of Vicia faba and Phaseolus vulgaris (Fabaceae) by fungal pathogens and their effects on the life-history parameters of Liriomyza huidobrensis (Diptera: Agromyzidae).Fungal Ecology 6: 293–301. 10.1016/j.funeco.2013.01.003 · doi ↗
- 4Ariyawansa HA Phukhamsakda C (2015) Revision and phylogeny of Leptosphaeriaceae.Fungal Diversity 74: 19–51. 10.1007/s 13225-015-0349-2 · doi ↗
- 5Bensch K Braun U (2012) The genus Cladosporium.Studies in Mycology 72: 1–401. 10.3114/sim 000322815589 PMC 3390897 · doi ↗ · pubmed ↗
- 6Bensch K Groenewald JZ (2015) Common but different: the expanding realm of Cladosporium.Studies in Mycology 82: 23–74. 10.1016/j.simyco.2015.10.00126955200 PMC 4774271 · doi ↗ · pubmed ↗
- 7Bensch K Groenewald JZ (2010) Species and ecological diversity within the Cladosporium cladosporioides complex (Davidiellaceae, Capnodiales).Studies in Mycology 67: 1–94. 10.3114/sim.2010.67.0120877444 PMC 2945380 · doi ↗ · pubmed ↗
- 8Bensch K Groenewald JZ (2018) Cladosporium species in indoor environments. Studies in Mycology 89: 177–301. 10.1016/j.simyco.2018.03.002PMC 590908129681671 · doi ↗ · pubmed ↗