The impact of the let-7 family on the pathophysiological mechanisms of traumatic brain injury: a systematic review
Natalia Radenza, Renata Mangione, Saviana Antonella Barbati, Francesco Bellia, Giuseppe Caruso, Antonio Belli, Giuseppe Lazzarino, Barbara Tavazzi, Giacomo Lazzarino, Angela Maria Amorini, Valentina Di Pietro

TL;DR
This paper reviews how the let-7 family of microRNAs behaves after traumatic brain injury and explores their potential as biomarkers and therapeutic targets.
Contribution
A systematic review of let-7 miRNA expression patterns and roles in traumatic brain injury pathophysiology.
Findings
Human studies show upregulation of let-7 family members in peripheral biofluids after TBI.
Animal studies suggest let-7 miRNAs modulate neuroinflammation, apoptosis, and metabolism post-TBI.
Let-7c and let-7i are linked to microglial activation and IL-6 regulation in TBI models.
Abstract
The lethal-7 (let-7) family of microRNAs (miRNAs) plays a crucial role in regulating key biological functions, including cell differentiation, inflammation, metabolism, and proliferation. Their dysregulation has been implicated in various diseases, including cancer and neurological disorders. Despite extensive knowledge of their roles in normal physiology and disease, the involvement of let-7 miRNAs in the pathophysiology of traumatic brain injury (TBI) remains incompletely understood. To systematically identify and analyze the differential expression of let-7 family members following TBI across human and animal models and to assess their potential role as diagnostic biomarkers and therapeutic targets. A systematic review was conducted in accordance with Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines. Literature was retrieved from PubMed,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
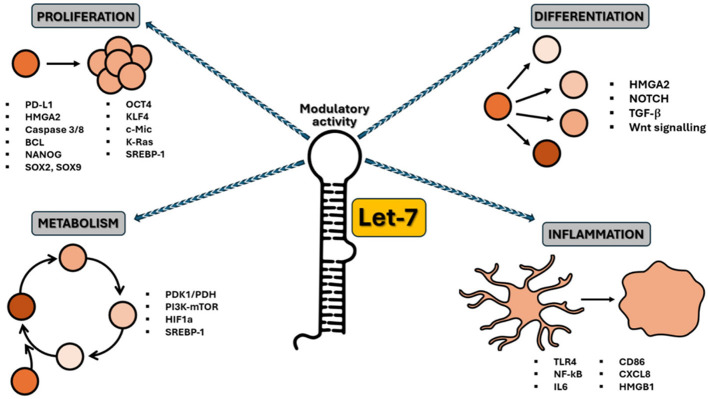 Figure 1
Figure 1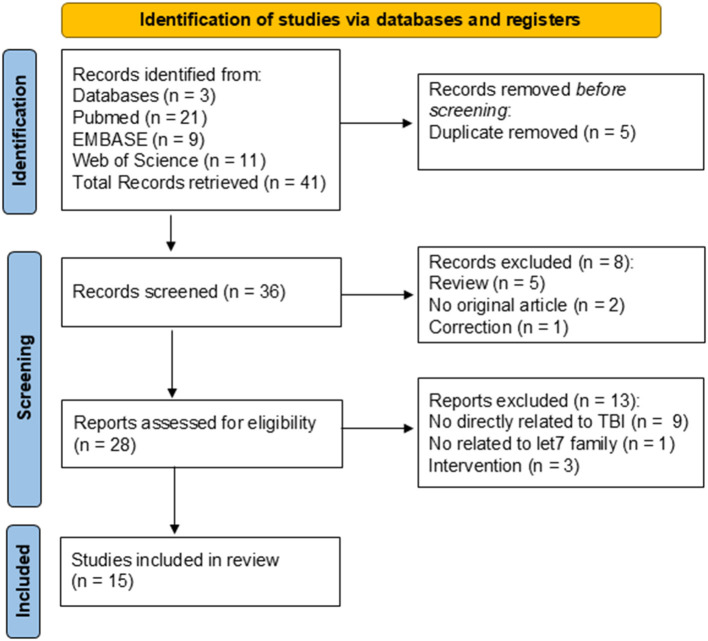 Figure 2
Figure 2|
|
|
|
|---|---|---|
| Development and cell differentiation | Let-7 family identified as a heterochronic gene regulating specific proteins to coordinate developmental timing in | ( |
| Developmental stage in | ( | |
| Let-7 performed an essential role in controlling cellular proliferation in the blastocyst and subsequent tissue development | ( | |
| Let-7 miRNAs play a functional role in the development of human neural progenitors by suppressing HMGA2 and NOTCH to regulate glycogenesis. Its normal expression is also considered critical for the maintenance of the blood–brain barrier | ( | |
| Let-7 expression throughout the development and maintenance of the cardiovascular system is highly significant in the protection and recovery of both endothelial cell and BBB integrity during normal conditions, as well as times of stress | ( | |
| Proliferation and tumor suppressor | Let-7 has been found to modulate signaling between fibroblast growth factors (FGF) and TGF-β in endothelial cells, limiting endothelial-to-mesenchymal transition and consequent proliferation | ( |
| Let-7 suppresses the translation of several oncogenes, including Myc, K-Ras, and HMGA2. Let-7 is deregulated in a wide variety of cancer types and influences CSC maintenance, metabolism, tumorigenesis, and metastasis. Let-7g was demonstrated to inhibit the expression of RAS in non-small cell lung cancer and consequently arrest tumorigenesis | ( | |
| Increased let-7a expression represses chemoresistance and tumorigenicity through the ablation of CSC-like properties. In gastric cancer, let-7b was shown to directly suppress c-myc expression to promote gastric stem cell differentiation and drug sensitivity | ( | |
| The biogenesis of let-7 is inhibited by LIN28A and LIN28B, two highly related RBPs and proto-oncogenes. Activation of either is responsible for the global post-transcriptional downregulation of let-7 miRNAs in many cancers, which is associated with advanced tumor stages | ( | |
| Wnt/β-catenin pathway represses let-7 expression post-transcriptionally by the transactivation of LIN28B to augment breast CSC phenotypes | ( | |
| LIN28A facilitates glioblastoma neurosphere formation through constitutively activating K-Ras and suppressing let-7b and let-7. Targeting LIN28 and/or restoring let-7 expression, by inhibiting tumor metabolic activities in CSCs, may provide potential therapeutic strategies for cancer treatment | ( | |
| Dysregulation of let-7 is thought to be influential in the development of vascular collaterals that support tumors | ( | |
| Elevated LIN28A/B expression boosts fatty acid synthesis, aiding cancer progression through SREBP1. LIN28A/B interact with SREBP-1 and SCAP mRNAs, increasing SREBP-1 translation and maturation, speeding up the conversion of fatty acids, and protecting cancer cells from lipotoxicity | ( | |
| Immune response | Let-7g can improve several endothelial functions, including a decrease in senescence, inflammation, monocyte adhesion, and an increase in angiogenesis | ( |
| Let-7c regulates dental inflammation by negatively affecting the DMP1-mediated NF-κB signaling pathway | ( | |
| Let-7i is involved in leukocyte modulation by inhibiting CD86, CXCL8, and HMGB1 expression | ( | |
| Let-7 family members differentially act to influence microglial functions | ( | |
| Let-7a targets HMGB1, which mediates microglial activation | ( | |
| Let-7 and miR-98 resulted in reduced leukocyte adhesion to and migration across endothelium, diminished expression of pro-inflammatory cytokines, and increased BBB tightness | ( | |
| Let-7i downregulates TLR4 and MMP9 in order to reduce pro-inflammatory signaling and protect BBB integrity | ( | |
| Macrophages exhibit high expression levels of miRNA; let-7c inhibits the polarization of macrophages toward the M1 phenotype while enhancing M2 polarization, modulating macrophage plasticity | ( | |
| Let-7 directly inhibits IL6 expression | ( | |
| Let-7 can also target receptor TLR4, affecting the inflammatory response | ( | |
| Let-7 is suppressed by LIN28 transcription induced by NFkB | ( | |
| Metabolism | PDK1 blocks glucose from entering mitochondrial oxidative phosphorylation (OXPHOS) and promotes aerobic glycolysis. In breast cancer stem cells (BCSCs), increased activity involves the long non-coding RNA H19 sequestering let-7 miRNA, which activates HIF1a and raises PDK1 expression | ( |
| Early studies with transgenic mice showed that the let-7/LIN28 axis regulates glucose metabolism. LIN28A/B represses let-7, which enhances glucose metabolism by activating the insulin-P13K-mTOR pathway and increasing insulin sensitivity | ( | |
| Neuroprotection and Neurodegeneration | Higher levels of let-7g* and miR-98 expression have been associated with decreased neurotoxicity following stroke and improved neurovascular perfusion | ( |
| Dying neurons release let-7, which acts as a damage-associated molecular pattern (DAMP) by binding to the TLR7 receptor. This interaction activates microglia and macrophages, leading to increased neurotoxicity. | ( | |
| In EAE, let-7 worsens disease by promoting harmful immune cell differentiation | ( | |
| In newts, reducing let-7 is essential for tail regeneration, as let-7 mimics hinder this process | ( | |
| Increased levels of let-7 have been suggested as potential biomarkers for multiple sclerosis, stroke, and Alzheimer's disease | ( | |
| miR-98 downregulates endothelial tight junction proteins, negatively affecting BBB integrity | ( | |
| miR-98 protects BBB integrity to positively influence ischemic stroke outcomes | ( | |
| miR-let-7a suppresses α-Synuclein-induced microglia inflammation through targeting STAT3 in Parkinson's disease | ( |
|
|
|
|---|---|
| Published in English | Studies published not in English |
| Primary research published in peer-reviewed journals | Non-primary data articles, Conference abstracts, commentaries, editorials, and letters |
| Traumatic Brain Injury, all severities and time-points | Other neuropathology |
| Human and animal studies | |
| Brain tissue and all biofluids | None |
| Qualitative and quantitative microRNA expression analysis, including at least one member of the let-7 family | Studies focusing on post-translational modifications, mutations/allelic variants, and intervention |
|
|
|
|
|
|
|
|---|---|---|---|---|---|
| Wyczechowska et al. ( | Human | Serum | Concussion | 2 h;18 h; end of season | let-7c (↑ 2 h+18 h vs. end of season); let-7c (↑ 2 h vs. end of season; ↑ 18 h vs. end of season); let-7b (↑ 18 h vs. 2 h and end of season) |
| Svingos et al. ( | Human | Serum | Concussion | Baseline; 4 h | let-7a (↑ 4 h) |
| Ma et al. ( | Human | Serum | Severe TBI | 2 h; 12 h; 24 h; 48 h; 72 h | let-7b and let-7f (↑ 12hvs2h); let-7a (↓12 h vs. 2 h); let-7f (↑ 24 h vs. 2 h); let-7a (↓24 h vs. 2 h); let-7b (↑ 48 h vs. 2 h); let-7f (↑ 72 h vs. 2 h) |
| Mitra et al. ( | Human | Plasma | Mild TBI | 28 days from injury | No changes in let-7f |
| Petrova et al. ( | Human | CSF, plasma | Severe TBI | from 2 M up to 2 Y with or without TBI | Let-7b (↓CSF); No changes in plasma |
| Seršić et al. ( | Human | CSF | Severe TBI | D 1-2, D 3-4, D 5-6, and D 7-12 CSF pools | let-7a, b, c, d, e, f, g, i (↓ over the time-points) |
| Di Pietro et al. ( | Human | Saliva | Concussion | 48 h/72 h | let-7i (↑ 36 h) |
| Johnson et al. ( | Human | Saliva | Concussion | 14 D; 4 W | let-7f (↑ PCS vs. ACS); let-7a associated with prolonged symptoms; let-7b associated with fatigue |
| Di Pietro et al. ( | Human | Saliva | Concussion | In-match; post-match; 36 h-48 h | let-7a, b, i, f (↑ in concussion vs. different controls) |
| Lv et al. ( | Mouse | Brain | CCI (impact velocity at 3.5 m/s, deformation depth at 2.0 mm, and duration for 150 ms) | from 6 h to 5 D | let-7c (↓ from 6 h to 5 days) |
| Miao et al. ( | Mouse | Brain | CCI (impact velocity 4.5 m/s, deformation depth at 1.5 mm) | 3 W | let-7c (↓) |
| He et al. ( | Mouse | Brain | HSI (2.5 mm posterior to bregma, 2.3 mm lateral to the sagittal suture, and 3 mm below the dura) | 3 D, 7 D, 14 D | let-7c (↓3D); let-7i (↓3D, 7D); |
| Johnson et al. ( | Rat | Brain, serum | Blast Injury (unilateral frontal 10% PBB) | 4 h; 7 D | let-7i (↑ 4 h, brain); let-7i (↓7 D, serum); |
| Balakathiresan et al. ( | Rat | Serum, CSF | Blast Injury (2.5-ft compression chamber connected to a 15-ft expansion chamber). SSI: three serial BOP of 120 kPa at an interval of 2 h; LII: 3 BOP of 120 kPa at an interval of 24 h | 3 h and 24 h after the last BOP exposure | let-7i (↑ in Serum and CSF of SSI at both time-points) |
| Sajja et al. ( | Mouse | Plasma | Blast injury (mild to moderate; 17, 17 × 3, and 20 psi blast) | let-7 a, b, g (↓ 17 x 3 psi) |
|
|
|
|
|
|
|
|
|
|
|
|---|---|---|---|---|---|---|---|---|---|
| Cohort | No | Yes | Yes | Yes | No | Yes | No | No | Yes |
| Control or comparison group | Yes | Yes | Yes | Yes | Yes | No | Yes | Yes | Yes |
| Pre/Post intervention data | No | Yes | No | Yes | No | No | No | No | Yes |
| Random assignment of participants to the intervention | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| Random selection of participants for assessment | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Follow-up rate of 80% or more | No | Yes | Yes | Yes | Yes | No | NA | Yes | Yes |
| Comparison groups equivalent in sociodemographic | Yes | Yes | No | NR | NR | NA | No | Yes | Yes |
| Comparison groups equivalent at baseline on disclosure | No | Yes | No | Yes | No | No | Yes | No | Yes |
| Overall Risk of Bias | High | Low | Unclear | Low | High | High | High | Unclear | Low |
|
|
|
|
|
|
|
|
|---|---|---|---|---|---|---|
| Was the allocation sequence adequately generated and applied? | Yes | Unclear | Unclear | Unclear | Unclear | Unclear |
| Were the groups similar at baseline, or were they adjusted for confounders in the analysis? | Yes | Yes | Yes | Yes | Yes | Yes |
| Was the allocation to the different groups adequately concealed during? | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear |
| Were the animals randomly housed during the experiment? | Unclear | Unclear | Yes | Yes | Unclear | Unclear |
| Were the caregivers and/or investigators blinded from knowledge of which intervention each animal received during the experiment? | Yes | No | Yes | Unclear | Unclear | Unclear |
| Were animals selected at random for outcome assessment? | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear |
| Was the outcome assessor blinded? | Yes | Yes | Yes | Unclear | Unclear | Unclear |
| Were incomplete outcome data adequately addressed? | Yes | Yes | Yes | Yes | Yes | Yes |
| Are reports of the study free of selective outcome reporting? | Yes | Yes | Yes | Yes | Unclear | Yes |
| Was the study apparently free of other problems that could result in a high risk of bias? | Yes | Yes | Yes | Unclear | Unclear | Yes |
| Overall Risk of Bias | Low | Low | Low | Unclear | Unclear | Unclear |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTraumatic Brain Injury Research · Traumatic Brain Injury and Neurovascular Disturbances · Cardiac Arrest and Resuscitation
1 Introduction
The lethal-7 (let-7) microRNA (miRNA, miR), named because its knockout was lethal during development (1), was the first miRNA discovered in humans (2). It is well conserved across species (3), from Caenorhabditis elegans (C. elegans) to humans, even though higher animals tend to have increased numbers of let-7 isoforms. Both C. elegans and Drosophila melanogaster (D. melanogaster) possess a singular let-7 isoform (let-7a), while humans and mice both possess 10 isoforms (miR-98, miR-202, and let-7a, 7b, 7c, 7d, 7e, 7f, 7g, and 7i) across 12 and 3 genetic loci, respectively (4, 5). The let-7 isoforms (apart from miR-202) contain a conserved ‘seed sequence' with the nucleotides GAGGUAG in positions 2 to 8 of each miRNA molecule (5, 6). The preservation of this sequence between isoforms implicates similarly conserved biological targets and evolutionarily retained functions.
Expression of let-7 is ubiquitous in adult mammalian tissues and known to increase throughout development (7), but the specific expression levels of each let-7 isoform are context-dependent and tightly regulated (8). Due to its roles in developmental differentiation, proliferation, inflammation, and cellular metabolism, dysregulation of let-7 expression has been implicated in several pathologies, including cancers and cerebral and cardiovascular disorders (5).
In embryogenesis, let-7 plays a crucial role in controlling the development from the first trimester to birth (9–11). In the central nervous system (CNS), let-7 enhances glycogen biosynthesis by suppressing HMGA2 and NOTCH (12) and is implicated in maintaining endothelial cell integrity (3), thus providing an optimal functionality of the blood-brain barrier (13). Given its role as a regulator of angiogenesis, let-7 can also control cancer progression (14). Reduced let-7 expression is a common hallmark of most cancers (15), leading to the upregulation of genes critical for DNA replication, such as programmed cell death ligand 1 (PD-L1) (16) and high mobility group AT-hook 2 (HMGA2) (17). Additionally, apoptotic genes like caspase 3 (18), caspase 8, and B-cell CLL/lymphoma (BCL) (19), or the pluripotency transcription factors NANOG, SOX2, SOX9, OCT4, KLF4, and c-Myc are modulated by let-7 and help the survival of cancer stem cells (CSCs) (20, 21).
The let-7 family appears critical also for the progression of inflammation, with various members associated with early inflammatory pathways (5) through the well-established mechanism of action via the LIN28/let-7 axis. LIN28B has been identified as a molecular link between inflammation and cancer through a positive feedback loop involving NF-κB activation and the release of repression from the let-7-regulated IL-6 gene (22). More recent studies have associated the LIN28/let-7 axis with glucose metabolism because of its regulatory effects on PDK1, the function of which is to inhibit the activity of the pyruvate dehydrogenase (PDH) subunits of the pyruvate dehydrogenase complex (PDHC). Increased glucose metabolism is attributed to LIN28A/B-mediated repression of let-7 (23). In addition, in breast cancer, whilst LIN28A and LIN28B promote the oxidative metabolism of glucose, let-7, by acting on PDK1, promotes glycolytic consumption of glucose in a hypoxia-independent manner. This study suggests a direct regulation of the LIN28/let-7 axis in the glycolytic/oxidative modulation of glucose metabolism, even under normal oxygen levels (24). Inhibition of the enzymatic function of PDH shifts glucose-dependent energy production from mitochondrial oxidative phosphorylation (OXPHOS) to glycolysis, thus acting as a gatekeeper of glucose metabolism. Increased activity in breast cancer stem cells (BCSCs) is also associated with elevated expression of the long non-coding RNA H19, which sequesters let-7 miRNA, leading to activation of HIF1a and enhancing downstream target PDK1 expression (25).
Furthermore, elevated expression of LIN28A/B enhances de novo fatty acid synthesis, promoting cancer progression via SREBP1. Lin28A/B directly associates with the mRNAs of both SREBP-1 and SCAP, enhancing the translation and maturation of SREBP-1, thereby accelerating metabolic conversion of saturated and unsaturated fatty acids and protecting cancer cells from lipotoxicity (26). A summary of the aforementioned functions of the let-7 family is presented in Table 1 and illustrated in Figure 1.
Schematic representation of the let-7 family activity as a crucial regulator of various cellular processes. Let-7 miRNAs regulate the cell cycle, particularly by inhibiting genes that drive cell division, thereby preventing uncontrolled proliferation. Let-7 can also act as a tumor suppressor, inhibiting the expression of oncogenes such as RAS, MYC, and HMGA2 and helping to prevent cancer progression. During development, let-7 miRNAs control cell differentiation by promoting the transition from stem cells to differentiated cells. Let-7 is also involved in regulating metabolic pathways, influencing lipid and glucose metabolism, and maintaining cell energy homeostasis. In the immune system, let-7 miRNAs modulate inflammation and immune cell activation, contributing to immune response regulation.
Despite the numerous evidence of let-7 implication in regulating the biological and metabolic functioning of nervous cells, there are no direct studies in models of neurodegeneration that have allowed us to establish whether its roles are neuroprotective or neurodegenerative. Most existing data link altered let-7 expression to neurological disorders such as multiple sclerosis, stroke, and Alzheimer's disease (27), while its role in traumatic brain injury (TBI) remains understudied. TBI is a leading cause of disability worldwide and the most frequent cause of death in individuals under 35 in Western countries (28).
Its global incidence is estimated at over 900 cases per 100,000 people annually, corresponding to nearly 69 million new cases each year. By 2030, the burden of TBI-related disability is expected to surpass that of Alzheimer's disease and cerebrovascular disorders, creating significant personal and socioeconomic costs (29, 30).
TBI is defined as a non-congenital injury to the brain caused by the nervous cell absorption of part of the external physical forces acting at the time of injury (primary insult), which induces the sudden alteration of a significant number of biochemical, metabolic, and molecular processes (secondary insult), modifying brain cell homeostasis and provoking transient or long-lasting (up to permanent) impairment of consciousness, neurocognitive deficits, neuromotor disabilities, or psychological disturbances. Grossly, TBIs are classified using the Glasgow Coma Scale (GCS), which categorizes TBI patients into mild (GCS range 13–15), moderate (GCS range 9–12), and severe (GCS range 3–8), based on a score obtained after specific neurological assessments, such as eye opening, motor, and verbal responses.
Despite this alarming epidemiological evidence, effective treatments remain scarce, and researchers are just beginning to unravel the complex cellular, molecular, and biochemical changes that influence TBI outcomes (31–35). It is now well accepted that the concept of the neurometabolic cascade involves a change in ionic cell homeostasis, mitochondrial dysfunction with energy penalty, sustained oxidative/nitrosative stress, glutamate excitotoxicity, glucose dysmetabolism, profound changes in protein and gene expressions, dangerous neuroinflammation, damage to the BBB permeability, and induction of apoptosis with a consequent significant increase in the nervous cell mortality rate (36, 37). The duration and the severity of the aforementioned neurochemical alterations are strictly dependent on the amount of energy absorbed at the time of impact and, ultimately, on the biomechanics of TBI.
Patients with mild (mTBI) injuries (including concussions, a peculiar type of mTBI frequently encountered in sports) often go undiagnosed because they are frequently characterized by rapidly resolving clinical symptoms. Notwithstanding, experimental and clinical studies have shown that mTBI and concussions provoke biochemical/metabolic/molecular changes in nervous cells lasting much longer than symptom resolution, which may have severe pathological and neurological consequences, particularly in the case of repeated injuries (38).
Diagnostic approaches for TBI rely on imaging and biomarkers. Although computed tomography (CT) remains the preferred method for assessing focal injuries, it lacks the sensitivity to effectively detect diffuse damage (39). Magnetic resonance imaging (MRI) offers greater resolution but is costly and less accessible (40). Among biochemical markers, proteins such as S100b, GFAP, and ubiquitin C-terminal hydrolase-1 (UCH-L1) (41, 42) have been widely studied, and the combination of GFAP and UCH-L1 has recently been FDA-approved for clinical use in evaluating mTBI (43). However, challenges in clinical translation persist, highlighting the need for novel biomarkers. Epigenetic mechanisms, including small non-coding RNAs (sncRNAs), DNA methylation, and histone modifications, have emerged as critical regulators of the post-injury response. miRNAs and snoRNAs are increasingly recognized as modulators of neuroinflammation and neuroprotection, while reciprocal regulation between miRNAs and epigenetic modifiers (e.g., DNA methyltransferases, histone deacetylases) suggests a dynamic network shaping long-term injury outcomes (44).
Within this context, several miRNAs, such as miR-21, miR-92a, miR-155, and members of the let-7 family, have shown altered expression following TBI and have been linked to processes including inflammation, metabolism, and neuronal survival (45). Importantly, miRNAs are highly stable in biological fluids and resistant to enzymatic degradation and freeze–thaw cycles, making them attractive candidates for non-invasive diagnostics (45). While emerging evidence supports their role as biomarkers, the contribution of let-7 specifically to TBI pathophysiology remains poorly defined. To clarify this relationship, the present review systematically evaluates the evidence for the differential expression of let-7 family members in TBI. It explores their potential role as both biomarkers and mechanistic mediators of injury outcomes.
To clarify current understanding, this systematic review examined the literature to answer the question: Which let-7 family members are differentially expressed in TBI?
2 Materials and methods
This systematic review was registered with PROSPERO in 2025, CRD420251129282. Available from https://www.crd.york.ac.uk/PROSPERO/view/CRD420251129282.
2.1 Systematic review
A systematic review was undertaken using the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines and the PRISMA checklist (46, 47). The research question, “To identify the differentially expressed let-7 family members in TBI,” enabled the generation of focused keywords that were used to search and retrieve relevant records from three databases. Explicit inclusion and exclusion criteria were developed and used to complete an abstract. The full-text screening of the records was conducted to determine those eligible for inclusion. This process was done by hand by a primary reviewer, who included articles that were subsequently checked by a second reviewer (NR, VDP).
2.2 Search strategy
The Population, Intervention, Comparison(s), and Outcome(s) (PICOs) framework for systematic reviews was utilized to define and focus the research question. This Cochrane Collaboration-recommended system ensures that the full scope of evidence is considered within defined parameters, and a quantitative investigation of the results can be undertaken.
- Population: All animal species.2) Intervention: TBI. Human participants will be assessed by a clinician, and any validated TBI animal model will be included.3) Comparisons: Healthy age-matched controls with no TBI or sham animals.4) Outcomes: The fold change in miRNA expression profiles between TBI and controls.
2.3 Search terms and databases
The following keywords were selected to represent the PICO criteria and combined with Boolean operators to generate the string search: (Traumatic Brain Injury OR TBI OR concussion) AND (let-7a OR let-7b OR let-7c OR let-7d OR let-7e OR let-7f OR let-7g OR let-7i OR mir-98 OR miR-202). No filters were applied. The search was carried out in three databases: PubMed, EMBASE through Ovid, and Web of Science. The records retrieved were collated in EndNote 20 (Clarivate, Philadelphia, PA, USA), where they were screened for duplicates, and any identified were removed. The remaining abstracts were then manually assessed for eligibility by the two independent reviewers using our predefined inclusion and exclusion criteria outlined in Table 2.
2.4 Data extraction
Data were extracted from the final included studies and imported into Microsoft Excel (Microsoft Corporation, Redmond, WA, USA) (Table 1). The title of the article, authors, and abstract summaries were saved to identify the studies. The species, tissue, TBI severity, TBI model, time points, and results, including let-7 up- or down-regulation compared to control groups, were recorded.
2.5 Risk of bias
Risk of bias was assessed using two different tools, the Evidence Project risk of bias tool for human studies (48) and the SYRCLE risk of bias (RoB) tool for animal studies (49). Risk for each study was assessed across the different domains by two independent reviewers (VDP and GL), with any disagreements being settled through discussion. The Evidence Project risk of bias tool is a tool for assessing the risk of bias across both randomized and non-randomized study designs. The tool consists of eight items, each assessed as either present (yes) or absent (no), some of which are also marked as not applicable or not reported. The items are as follows: (1) cohort, (2) control or comparison group, (3) pre-post intervention data, (4) random assignment of participants to the intervention, (5) random selection of participants for assessment, (6) follow-up rate of 80% or higher, (7) equivalence of comparison groups in socio-demographics, and (8) equivalence of comparison groups at baseline regarding outcome measures. Items (1)–(3) provide an overview of the study design, while the other items address additional aspects of study rigor. The SYRCLE risk of bias (RoB) tool for animal studies instead used the following domains to assess the potential risk of bias across the studies: random sequence generation, baseline characteristics described, correct timing of randomization, allocation concealment, random housing, blinding, random outcome assessment, incomplete data, sample size calculation, and primary outcome specified.
3 Results
3.1 Systematic review
Electronic searches of PubMed, EMBASE, and Web of Science generated 21, 9, and 11 records, respectively, making a total of 41 records. After duplicate removal, 36 abstracts were screened, and n = 8 were excluded because of the review or the abstract. Twenty-eight full-text records were assessed for eligibility, and 15 papers met all the criteria and were included in this review. Causes for exclusion were: no direct relationship to TBI (n = 9), no relationship to the let-7 family (n = 1), and intervention (n = 3). The PRISMA diagram is presented in Figure 2.
PRISMA diagram of systematic review search.
3.2 Data extraction
The primary data were extracted from each study and listed in Table 3, showing references, species, tissues, TBI severities or models, time points for assessment, and expression of let-7 family members.
3.3 Risk of bias
The Evidence Project risk of bias tool indicated that five are cohort studies, and most of the studies have a control or comparison group. Only three studies collected data pre- and post-intervention. Random assignment of participants to the intervention was not applicable for all studies; while datasets were nearly complete, the equivalent comparison groups on socio-demographics or baseline disclosure varied across studies. This variation resulted in a high overall risk of bias for four studies, an unclear risk for two, and a low risk for the remaining three (Table 4). RoB analysis found that all included articles, referring to data obtained in experimental TBI in laboratory animals, specified the primary outcomes and described baseline characteristics. All studies had full data availability with no missing data. However, for many of them, it is unclear whether the randomly housed animals or concealed treatment allocations. Incidents of random sequence generation, blinding, and random outcome assessment were not always specified. Thus, the overall risk of bias was judged to be unclear for three studies, potentially invalidating certain findings of the included papers, and low for the remaining three studies. RoB analysis results are depicted in the table below (Table 5).
3.4 Human studies
Wyczechowska and Svingos have investigated the expression of specific microRNAs in the blood of small groups of collegiate football players. Wyczechowska et al. (50) examined 21 athletes (18–23 years of age) over an entire practice and game season. Of them, 10 experienced a concussion, and the remaining 11 non-concussed athletes were used as the matched control group. Authors found that serum let-7c-5p increased in the concussed group during the acute phase post-injury (at 2 and 18 h) and demonstrated that circulating let-7c-5p levels had high specificity and sensitivity for distinguishing concussed from non-concussed players. Additionally, it was found that serum let-7b-3p was also elevated at 18 h post-injury. In the second study, Svingos et al. (51) studied 27 collegiate athletes, with a mean age of 18.8 ± 0.8 years, who were analyzed for let-7 levels in serum from peripheral blood before the beginning of the season (baseline) and within 4 h post-concussion. The levels of let-7a-5p showed a significant increase when comparing post-concussion measurements to the baseline values. In another study, Ma et al. (52) used serum as the sample of choice for let-7 determination. They selected circulating microRNA expression, which was evaluated in 20 patients (aged 32–56 years) at different times (2, 12, 24, 48, and 72 h) following severe TBI. The authors found that whilst let-7a was downregulated, let-7b and let-7f were upregulated. Finally, Mitra et al. found no changes in let-7f between individuals with mTBI who report post-concussive symptoms at the 28-day mark and those who do not (53).
Petrova and Seršić focused on cerebrospinal fluid (CSF) for the evaluation of let-7 levels following head injury. Petrova et al. (54) measured hsa-let-7b-5p in both CSF and blood of patients affected by prolonged disorders of consciousness (lasting from 2 months to 2 years) caused by prior severe brain damage. In 22 of them, the previous brain injury was caused by a severe TBI. Compared to the control group (10 patients undergoing surgical operations due to extracerebral volumetric brain neoplasms, meningiomas, and basilar artery aneurysms), it was found that the subgroup of severely injured TBI patients had an upregulation of let-7b in CSF but not in blood. Seršić et al. (55) analyzed 35 CSF samples from five patients with severe TBI over 12 days post-injury, pooling samples into intervals (days 1–2, 3–4, 5–6, and 7–12). They targeted 87 miRNAs and found high levels for many of them, including let-7a, b, c, d, e, f, g, and i, on the first day of post-injury, with a progressive decline for longer times after severe TBI.
Finally, three studies on concussed patients used saliva as the body fluid in which to perform the quantification of let-7. Di Pietro et al. conducted two of these studies. In 2018 (56), they collected saliva samples from 22 concussed athletes and 10 matched controls 48–72 h post-concussion, finding that let-7i-5p was significantly upregulated in the concussed group. A larger follow-up study in 2021 (57) involved male professional players in England's top rugby union tiers across two seasons (2017–2019). Samples were taken pre-season from 1,028 players and during standardized head injury assessments (HIAs) from 156 players at three time points (in-game, post-game, and 36–48 h post-game), along with samples from 102 uninjured players and 66 with musculoskeletal injuries. This study revealed increases in let-7a, b, i, and f in the saliva of concussed players compared to the different control groups, with let-7f-5p showing the highest area under the curve (AUC) at 36–48 h.
In the third study, Johnson et al. (58) evaluated the efficacy of measuring salivary microRNAs in identifying children at risk for prolonged concussion symptoms. In this cohort study of 52 patients aged 7–21 years, microRNAs were measured within 14 days after head injury and at 4 and 8 weeks following concussion. It was found that there was upregulation of salivary let-7f in patients with post-concussion syndrome (PCS) compared to those with acute concussion. Additionally, let-7a-3p accurately identified patients with prolonged symptoms, while high salivary levels of let-7b were associated with fatigue.
3.5 Animal studies
Three studies have indicated a decrease in let-7-5p levels in TBI-injured mouse brains. Lv et al. (59) demonstrated that let-7c-5p levels dropped rapidly after controlled cortical impact (CCI) TBI and gradually returned to baseline by 14 days post-injury. Miao et al. observed a downregulation of let-7c following TBI induced by a pneumatic device, with variations noted in mice that engaged in voluntary exercise prior to the injury (60). Additionally, He et al. (61) reported a reduction in let-7c, 3 days after a hippocampal stab injury (HSI), also noting downregulation of let-7i at 2- and 7-days post-injury. Johnson et al. (62) examined let-7i in a rat model of penetrating ballistic-like brain injury (PBBI), collecting ipsilateral brain tissue and serum from 4 h to 7 days post-injury. They found that brain tissue levels of let-7i increased at 4 h, while serum let-7i decreased in serum at 7 days after PBBI. Balakathiresan et al. (63) investigated serum and CSF levels of let-7i following post-blast wave exposure in two injury groups: a short interval injury group (SII), which was exposed to three serial blasts at 120 kPa every 2 h, and a long interval injury group (LII), where blasts were administered every 24 hours. Samples from both groups were collected at 3 and 24 h after the last blast, revealing the presence of let-7i in both serum and CSF immediately after injury. Sajja et al. (64) utilized a murine model of mild-to-moderate blast-induced TBI, exposing animals to blast intensities of 17, 17 × 3, and 20 psi using a calibrated blast simulator. The study found that plasma levels of brain-enriched miRNAs let-7a, b, and g were reduced at 17 × 3 and 20 psi, and let-7c was downregulated at 17 × 3 only, while let-7d levels increased in the 17 psi group.
4 Discussion
This systematic review revealed the involvement of the let-7 family in the pathobiological mechanisms of TBI across varying severities and time points. Data from experimental TBI in laboratory animals and humans have notably highlighted that the different expression patterns depend on whether let-7 levels are measured in brain tissue (animal studies) or biofluids (animal and clinical studies).
Human studies showed an overall upregulation of let-7 family members across different biofluids, such as plasma, serum, and saliva, regardless of trauma severity (ranging from concussions to severe TBIs), population demographics (athletes vs. the general population), age, gender, or time since injury (from one day post-injury onward). The only exception was in CSF, where downregulation was predominant. In contrast, animal studies primarily demonstrated a downregulation of let-7 expression in brain tissue, with only a few exceptions. This pattern is not unique to TBI and aligns with findings in other diseases, such as cancer, where tissue levels of let-7 are generally low, but levels in the bloodstream are elevated, suggesting its potential as a biomarker (65–67). The inverse trend between tissue and biofluid levels may indicate a common regulatory mechanism for microRNAs, independent of the pathology considered, where cells release microRNAs to decrease their intracellular concentration, resulting in elevated levels in biofluids.
Based on the analysis of available literature, it can be stated that human studies have primarily focused on the use of let-7 as a biomarker for diagnostic and prognostic purposes. However, neither goal has been achieved to date, since (i) no correlation between let-7 levels in biofluids and TBI severity has been found, and (ii) no clear data have yet been produced correlating let-7 levels in biofluids with TBI patients' outcome.
Results of animal studies have explored more mechanistic aspects of the potential roles of let-7 variations in TBI. In the intricate landscape of one of the most complex and challenging pathologies affecting the brain, the let-7 family emerges as a crucial player in orchestrating the protective responses of nervous cells following TBI. Let-7 appears to play a crucial role in modulating the inflammatory cascade that typically follows TBI, which can both harm and help CNS recovery. Neuroinflammation is mainly driven by microglia and infiltrating macrophages, which amplify inflammation by releasing soluble cellular components in the injured brain (68, 69). Microglia, acting as the first responders to CNS injuries, can adopt either a “classically activated” M1 phenotype or an “alternatively activated” M2 phenotype (70, 71). While M1 polarization is associated with neuronal dysfunction and cell death, M2 polarization promotes anti-inflammatory responses, aids in debris clearance, and supports CNS repair. Modifying the M2/M1 ratio has shown promising therapeutic effects in TBI (72). MicroRNAs have been implicated in this modulation, with let-7c being one of the most abundant and conserved miRNAs linked to macrophage polarization (73, 74). Lv et al. (59) showed that let-7c was rapidly reduced in traumatically injured foci in the brains of adult C57BL/6J mice. The intracerebroventricular injection (ICV) of let-7c improved the outcomes of mice subjected to CCI by inhibiting neuroinflammation. In particular, the attenuation of microglia/macrophage activation was obtained through the inhibition of M1 polarization and enhancement of M2 polarization. In addition, authors showed that expressions of the let-7c putative target gene caspase-3 and of PKC-d (a modulator of caspase-3 effects) were inhibited. Miao et al. (60) also found that let-7c could be involved in the prevention and protection afforded by voluntary exercise in a mouse model of TBI induced by a pneumatic impact device. They showed that voluntary running wheel (RW) exercise prior to TBI significantly reduced the mortality rate and righting time in injured mice. They also showed that, following TBI, brain levels of let-7c were significantly different in the group of exercised mice compared to those of the untrained. Therefore, the authors proposed that an epigenetic mechanism, capable of inducing amelioration of functional deficits, might be involved in the beneficial effects of voluntary RW exercise following TBI.
It appears that let-7 can also help in minimizing TBI-associated secondary injury, particularly the neuroinflammatory processes caused by the release of pro-inflammatory cytokines (like TNF-α and IL-6), which can inadvertently cause further tissue damage. In this light, Johnson et al. used the rats' penetrating ballistic-like brain injury (PBBI) model to determine if changes in microRNA levels, including let-7i, were connected to those of neuroinflammatory mediators. Results showed an inverse correlation between let-7i and IL6, an interaction that was also previously described (75), suggesting let-7i as a plausible therapeutic target of TBI-induced inflammation. Hence, let-7 serves to mitigate the TBI-exacerbated neuroinflammatory response and directs immune cells toward protective functions. In this context, He et al. not only showed let-7i downregulation in a model of HSI in mice but also found that non-invasive intranasal delivery of let-7i significantly improves cognitive function and suppresses neuroinflammation, glial scar formation, and neuronal apoptosis via inhibition of the stimulator of interferon genes (STING). STING is a key regulator of inflammation, and its expression was found to be elevated in postmortem human TBI brains (76, 77). Balakathiresan et al. (63) and Sajja et al. (64) instead limited their studies to investigating microRNAs as potential biomarkers of TBI. They both found several members of the let-7 family differentially expressed in the CSF and serum of rodents undergoing blast injury.
However, since let-7′s roles are not limited to modulating mechanisms of inflammation, it is evident that the information obtained so far, in both experimental and clinical studies in TBI, is certainly not sufficient. For instance, several indications have been produced showing that the activity of the PDH complex, the key multi-enzymatic system ensuring the oxidation of pyruvate to form acetyl-CoA and thus fueling the Krebs cycle, is deeply modulated by let-7 through its action on the gene expressing the main PDH complex inhibitor (PDK, pyruvate dehydrogenase kinase). Using Marmarou's model of closed-head weight drop TBI in the rat, it was shown that profound glucose dysmetabolism (32) is associated with the PDH complex inhibition because of overexpression of PDK and concomitant downregulation of PDP (pyruvate dehydrogenase phosphorylase, the main activator of PDH) (78).
Additionally, although obtained by bioinformatic data only, it has also been indicated that let-7 may influence the homeostasis of N-acetylaspartate (NAA), the main neuron-specific N-acetylated amino acid deeply altered by TBI (79). Since the metabolism of NAA is strictly dependent on acetyl-CoA bioavailability (this compound is the acetyl group donor in the NAA synthesis catalyzed by NAT8L), it is easy to hypothesize that the let-7 modulation of both ASPA and PDK may have profound consequences on brain metabolism following TBI.
It should not, however, be disregarded that let-7 family members can influence neurogenesis. Particularly, the differentiation of neural stem cells into neurons in the TBI-injured brain may be highly relevant in aiding the replacement of lost or damaged cells after TBI. It should even be worth investigating whether they play a role in enhancing synaptic plasticity, which is crucial for functional recovery in cognitive tasks of the post-injured brain. Among the other potential roles of let-7 in the post-injury brain, the mitigation of oxidative/nitrosative stress, one of the mechanisms of nervous cell damage potently activated by TBI, might certainly be better characterized. Indications obtained under different pathological conditions, but TBI, suggest that let-7 is mutually regulated by FOXO3 and PRDX1 expression levels, both involved in the cellular capacity to maintain a correct oxido-reductive state of -SH groups of cysteine, deeply altered under conditions of oxidative/nitrosative stress (80). Also in this case, it has been shown that TBI deeply alters the cerebral concentrations of GSH, the main cellular antioxidant specific for keeping free -SH groups in the correct oxido-reductive state (32, 37), thereby rendering plausible the involvement of let-7 in the overall damaging process mediated by excess production of ROS and RNS occurring following TBI. Modulation of PI3K/Akt and MAPK pathways, thus promoting cell survival and growth, reinforces the brain's resilience in the wake of injury.
In the aftermath of TBI, the let-7 family serves as a multifaceted protector, navigating the complex interplay of inflammation, mitochondrial-related energy production, oxidative/nitrosative stress, and regeneration; each of them, if positively affected by let-7 modulation, may greatly improve nervous cell survival and, potentially, the outcome of TBI patients. Their contributions are vital for healing, illuminating pathways for future therapies that could help countless individuals reclaim their lives after the devastation of brain injury.
5 Limitations
The present research highlighted the promise of let-7 as both a biomarker and a therapeutic target. Changes in its expression can indicate the severity of TBI, offering insights into patient prognosis. Moreover, the potential to manipulate let-7 levels, through mimics or inhibitors, opens new avenues for treatment, aiming to harness its protective capabilities and enhance recovery. However, the use of miRNA signatures as a novel diagnostic and prognostic tool remains largely in the descriptive phase yet. While a wealth of data has been gathered across various disease states, translating this information into clinical applications requires larger studies and universally accepted guidelines. One major issue is the need for standardized protocols for miRNA extraction, quantification, and analysis. Variability in sample collection, processing methods, and analytical techniques can lead to inconsistent results, hindering the establishment of universally accepted miRNA biomarkers. A shared information infrastructure for data exchange, analysis, and protocol standardization would greatly enhance research in miRNA biomarker discovery. Furthermore, the biological complexity of miRNA regulation necessitates a deeper understanding of their roles in specific contexts. For instance, miRNA expression can be influenced by factors such as age, gender, lifestyle, and environmental conditions. Therefore, identifying well-characterized endogenous miRNAs that are consistent across different populations and conditions is crucial for their clinical applicability.
Regarding animal models of TBI, these are essential for understanding the underlying mechanisms and potential treatments, yet the variety of methodologies and tissues analyzed can lead to discrepancies in results. Methodologically, injuries can be induced through various techniques, including controlled cortical impact, fluid percussion, weight drop, or blast overpressure, leading to differences in injury severity and patterns. Additionally, the specific tissues analyzed, ranging from the brain cortex to subcortical structures and the cerebrospinal fluid, can yield varying insights into the pathophysiological changes following TBI. For example, while some studies focus on acute inflammatory responses in the cortex, others might assess long-term neurodegenerative changes in deeper brain regions. This variability can complicate comparisons across studies and hinder the establishment of a consensus on the biological mechanisms involved in TBI, highlighting the need for standardized protocols and comprehensive analyses.
Importantly, our review is limited by the quality of available evidence. Of the 15 studies included, 9 showed an unclear or high risk of bias. This limitation significantly affects the strength of our conclusions and underscores the need for further high-quality, standardized, and rigorous research in this field. Establishing universally accepted guidelines and infrastructure for data sharing and analysis will be crucial to advancing miRNA-based diagnostics and therapeutics in TBI.
6 Conclusions
We found that the let-7 family of microRNAs plays a multifaceted role in the response to TBI. By regulating inflammation, promoting neuronal survival, enhancing neurogenesis, and modulating key signaling pathways, let-7 microRNAs play a crucial role in mitigating damage and facilitating recovery. Their potential as biomarkers and therapeutic targets highlights their importance in advancing TBI treatment strategies. miRNAs may soon become invaluable tools in clinical practice, revolutionizing the way we approach disease management and patient care. Further research into their specific mechanisms and effects may lead to innovative approaches for improving outcomes in TBI patients; however, such research is mandatory.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Reinhart BJ Slack FJ Basson M Pasquinelli AE Bettinger JC Rougvie AE. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature. (2000) 403:901–6. 10.1038/3500260710706289 · doi ↗ · pubmed ↗
- 2Pasquinelli AE Reinhart BJ Slack F Martindale MQ Kuroda MI Maller B. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature. (2000) 408:86–9. 10.1038/3504055611081512 · doi ↗ · pubmed ↗
- 3Ali A Bouma GJ Anthony RV Winger QA. The Role of LIN 28. Int J Mol Sci. (2020) 21:3637. 10.3390/ijms 2110363732455665 PMC 7279312 · doi ↗ · pubmed ↗
- 4Hertel J Bartschat S Wintsche A Otto C Stadler PF. Lab Sot BC. Evolution of the let-7 micro RNA family. RNA Biol. (2012) 9:231–41. 10.4161/rna.1897422617875 PMC 3384580 · doi ↗ · pubmed ↗
- 5Bernstein DL Jiang X Rom S. micro RN As: their role in cerebral and cardiovascular diseases, inflammation, cancer, and their regulation. Biomedicines. (2021) 9:606. 10.3390/biomedicines 906060634073513 PMC 8227213 · doi ↗ · pubmed ↗
- 6Ruby JG Jan C Player C Axtell MJ Lee W Nusbaum C. Large-scale sequencing reveals 21U-RN As and additional micro RN As and endogenous si RN As in C. elegans Cell. (2006) 127:1193–207. 10.1016/j.cell.2006.10.04017174894 · doi ↗ · pubmed ↗
- 7Büssing I Slack FJ Grosshans H. let-7 micro RN As in development, stem cells and cancer. Trends Mol Med. (2008) 14:400–9. 10.1016/j.molmed.2008.07.00118674967 · doi ↗ · pubmed ↗
- 8Lee H Han S Kwon CS Lee D. Biogenesis and regulation of the let-7 mi RN As and their functional implications. Protein Cell. (2016) 7:100–13. 10.1007/s 13238-015-0212-y 26399619 PMC 4742387 · doi ↗ · pubmed ↗