Effects of Tagetes erecta on the Pigmentation of Large Yellow Croaker(Larimichthys crocea)
Siyu Zhang, Tianwei Wang, Lin Chen, Chengwei Huang, Yali Wang, Rongrong Ma, Jianping Wang, Zehui Su, Subin Cui, Dongmin Zhu, Jiahui Zhu, Kun Hu

TL;DR
This study shows that adding Tagetes erecta to the diet of large yellow croaker improves their body color more effectively than other supplements, boosting their commercial value.
Contribution
The study demonstrates that Tagetes erecta extract enhances body color in L. crocea more effectively than astaxanthin or amaranth extract.
Findings
T. erecta increased the a∗ value (red–green) 1.25-fold compared to the control group.
The ventral b∗ value (yellow–blue) was significantly elevated (p = 0.028) in the T. erecta group.
T. erecta improves color by regulating carotenoid uptake and melanogenesis.
Abstract
Larimichthys crocea (L. crocea), an important species in mariculture, is highly valued for its nutritional benefits, delicate flavor, and distinctive golden body, making it a popular choice among consumers. However, suboptimal breeding conditions and high stocking densities, often lead to reduced flesh texture and a deterioration in body color. Given the significant role body color plays in determining the commercial value of this species, this study evaluated the impact of various dietary supplements on color enhancement. One basal diet and three separate experimental diets were formulated: one supplemented with 200 mg/kg Tagetes erecta extract (2% lutein), one with 200 mg/kg astaxanthin (2%), and one with 200 mg/kg amaranth (Amaranthus spp.) leaf extract (85%). The results indicated that T. erecta was more effective in enhancing the body color of L. crocea than the other two…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1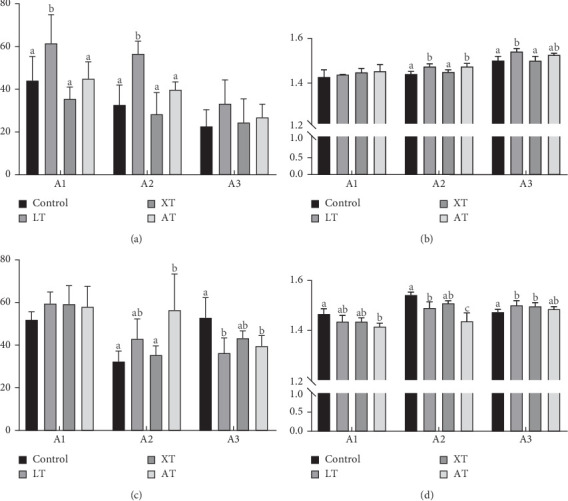 Figure 2
Figure 2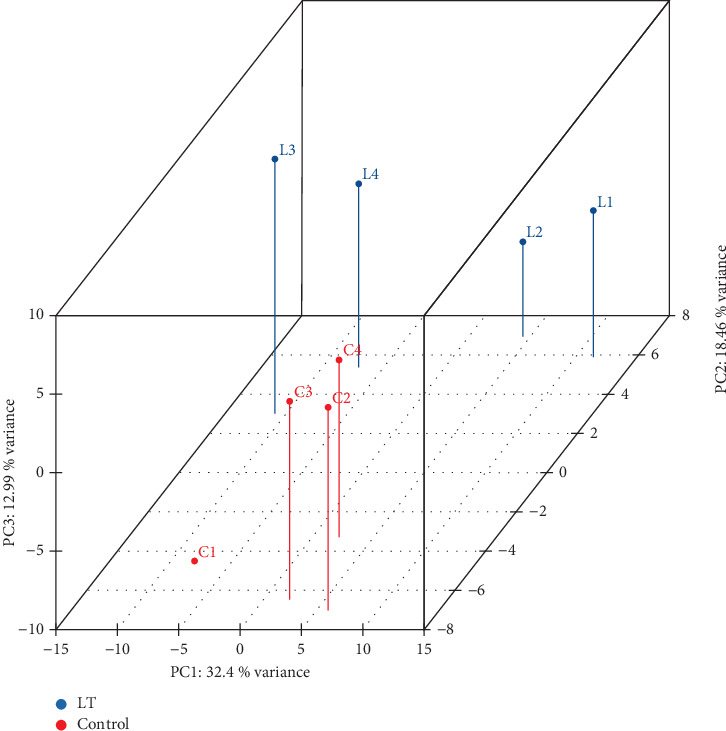 Figure 3
Figure 3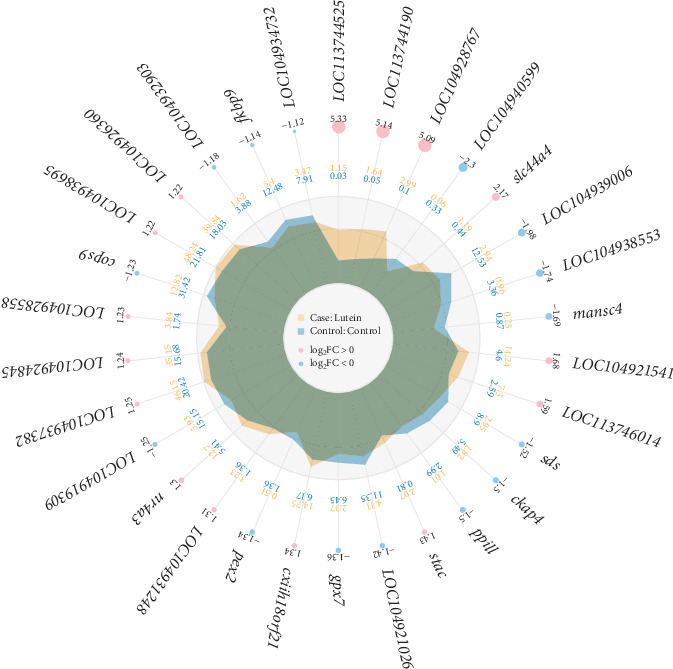 Figure 4
Figure 4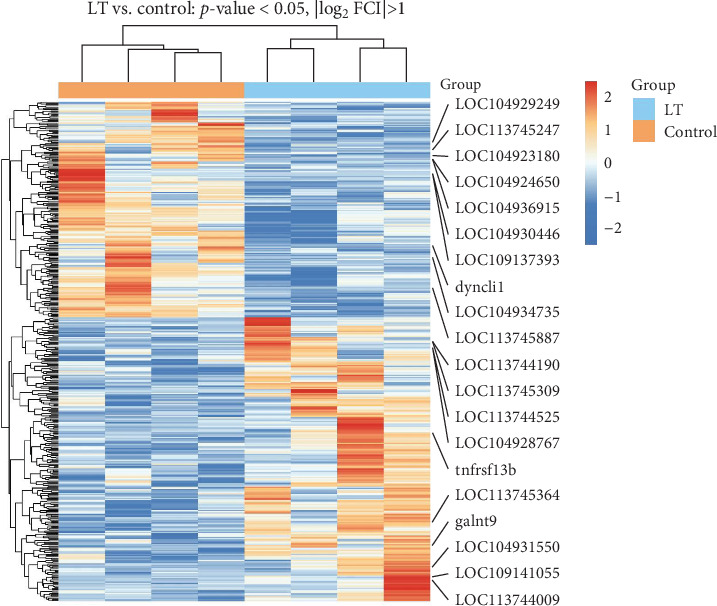 Figure 5
Figure 5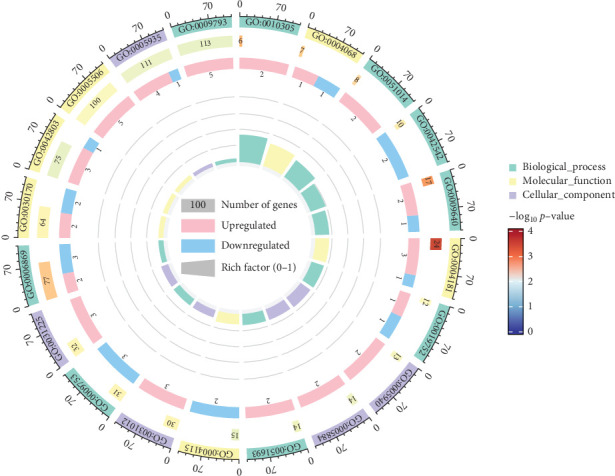 Figure 6
Figure 6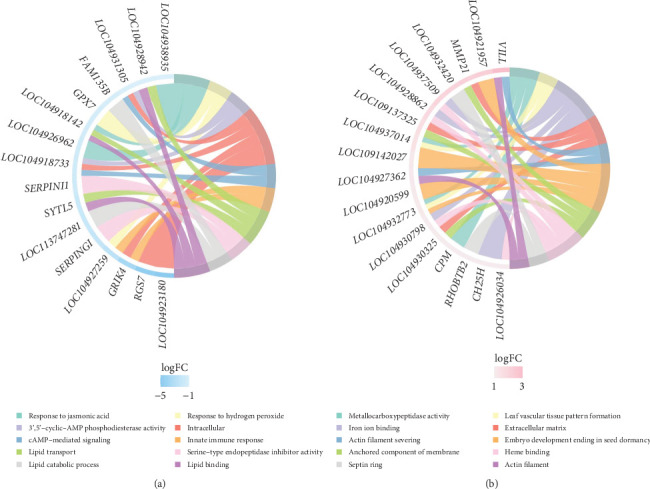 Figure 7
Figure 7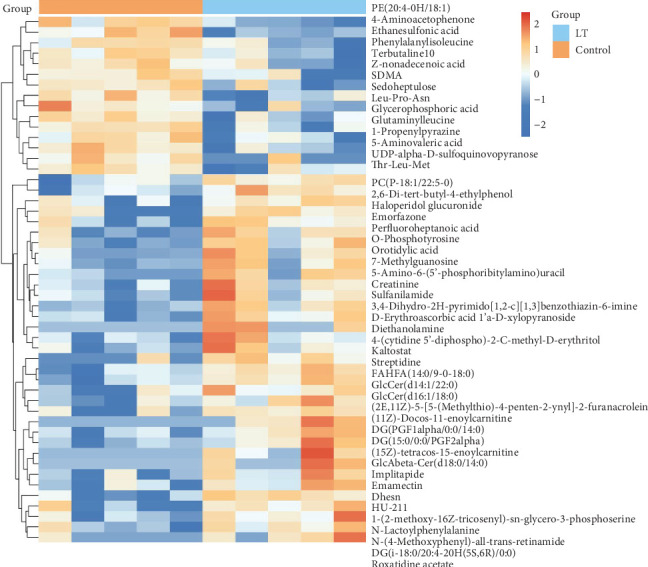 Figure 8
Figure 8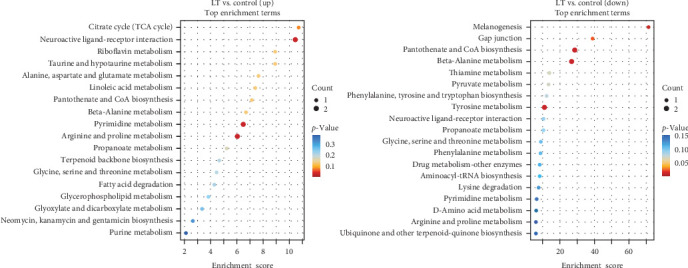 Figure 9
Figure 9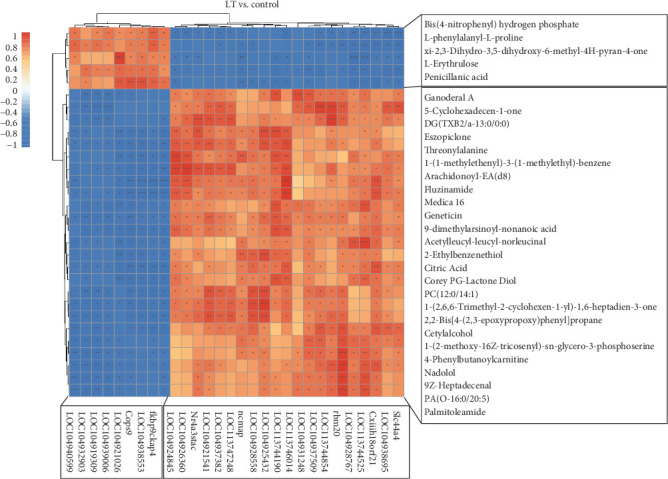 Figure 10
Figure 10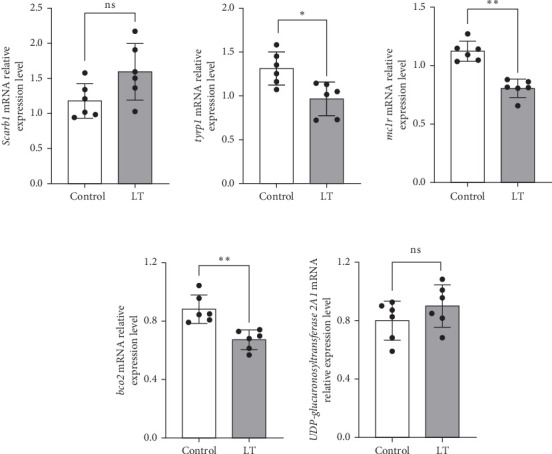 Figure 11
Figure 11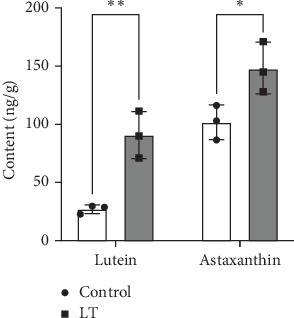 Figure 12
Figure 12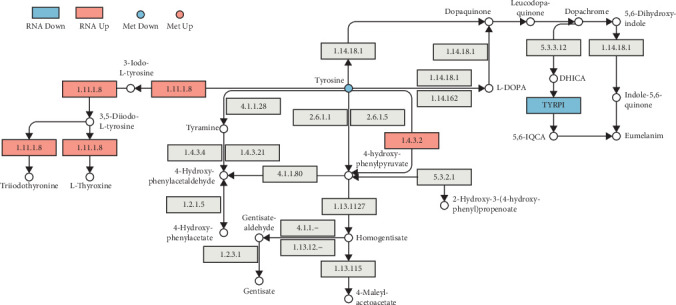 Figure 13
Figure 13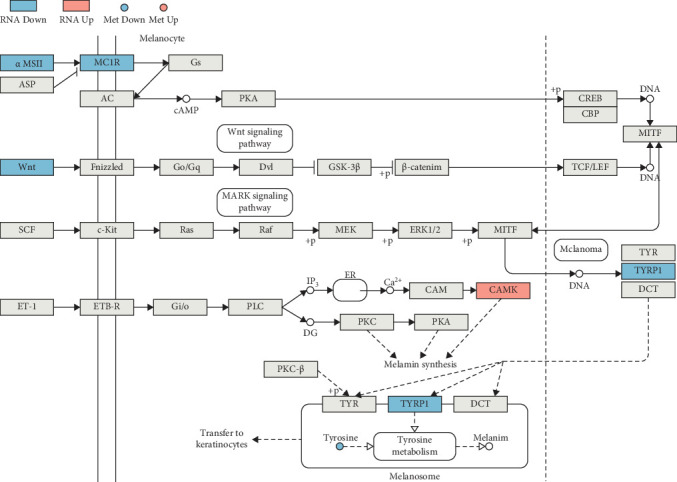 Figure 14
Figure 14- —Scientific and Technological Project of Ningbo
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquaculture Nutrition and Growth · Antioxidant Activity and Oxidative Stress · Animal Nutrition and Physiology
1. Introduction
Larimichthys crocea (L. crocea), as a key marine aquaculture organism in China, holds the top position in annual mariculture yield [1, 2]. It is highly prized for its rich nutritional content, including protein, vitamins, and trace elements, as well as its tender meat and pleasing taste [3]. Owing to the declining availability of wild population resources, artificial breeding has become increasingly prevalent in recent years. Despite this, cultured L. crocea faces challenges such as diminished skin pigmentation, degraded muscle flavor, and a looser meat texture compared to wild specimens [4–6]. Body color is an essential indicator of both the health and market value of fish and it plays a crucial role in classification within the industry. Thus, maintaining or enhancing the color characteristics of L. crocea during artificial breeding is vital to improving its economic value.
Fish body coloration is composed of chromatophores and carotenoids serve as the primary color-presenting substances in erythrophores and xanthophores. The formation of chromatophores relies on gene-governed neural crest-derived precursor cells [7]. Fish are unable to synthesize carotenoids de novo, so their pigmentation primarily depends on carotenoids in their diet [8]. Studies have shown that supplementing with carotenoids can enhance the body color of fish [9]. Common carotenoids found in fish include β-carotene, lutein, zeaxanthin, tunaxanthin, and astaxanthin. Regarding body color enhancement, Yi et al. [10] found that the body color of L. crocea is largely influenced by the deposition of yellow pigments. Supplementing the diet of juvenile L. crocea with astaxanthin and lutein for 9 weeks significantly increased the red and yellow values of their skin. Similarly, Yi et al. [11] demonstrated that incorporating shrimp shell powder into the feed improved epidermal pigmentation while elevating carotenoid compound concentrations in L. crocea.
However, there remains a gap in understanding the effects of natural plant-extract pigment supplements on body color in L. crocea. This study selected three widely distributed, highly safe, and carotenoid-rich plant extracts, to explore the effects of these three kinds of pigment supplements on the body color formation of L. crocea.
Tagetes erecta L., a plant rich in natural lutein, has been widely used as a coloring agent in food products and as a nutritional component in diets for both humans and livestock. The petals of T. erecta are a major source of lutein for the food and pharmaceutical industries [12]. In zebrafish (Danio rerio), bioactive components derived from T. erecta interact with the melanocortin-1 receptor, potentially inhibiting melanin biosynthesis [13]. However, the effects of marigold (T. erecta) on skin coloration in L. crocea have not been systematically investigated. Another source of natural pigments, Amaranthus tricolor, is recognized for its high antioxidant content, pigments, minerals, and phytochemicals [14]. In the Chinese feed additive variety list (2013), amaranth is a colorant that can be used in pets and ornamental fish. Known for its nontoxicity, bright color, and stable nature, it is frequently used as a safe and nutritious natural edible pigment [15]. Astaxanthin, a well-known carotenoid, is commonly included in fish feed to enhance red pigmentation and improve the overall body color of fish.
Consequently, the present study explored the impact of three distinct experimental diets, respectively, on the apparent color change and underlying mechanisms in L. crocea, with the aim of providing a reference for improving both color and production in artificial breeding systems. This study conducted a comparative assessment of the differential impacts on pigmentation in L. crocea among three separate dietary supplements: T.erecta, amaranth (Amaranthus spp.) leaf extract, and astaxanthin—a pigment commonly used in aquaculture.
2. Materials and Methods
The experimental procedures involving animal subjects and the research methodology underwent rigorous evaluation and received formal approval by the Ethics Committee of Shanghai Ocean University (Approval Number: SHOU-DW-2023-17).
2.1. Fish Management and Experimental Design
The breeding site was located at the cage base of Baishi Mountain, Xiangshan Port, Ningbo Institute of Ocean and Fisheries Research, Zhejiang Province. Healthy “Yongdai No.1” L. crocea fry were selected and placed in a seawater cage (3.0 m × 3.0 m × 4.0 m) for 2 weeks, during which they were fed with pellet feed provided by Zhejiang Qiangpu Biotechnology Co., Ltd. prior to the formal experiment, the commercial feed contained 11% moisture, 46% crude protein, and 2% crude fat. After the 2-week temporary feeding period, the experimental fish were starved for 24 h. About 30–40 g L. crocea juveniles with strong physiques and consistent specifications were randomly allocated into experimental groups, with each enclosure containing a cohort of 150 individuals. To explore the effect of feeding natural pigments on the body color of L. crocea during the breeding process, three experimental groups were setup: T. erecta extract addition group (LT), astaxanthin addition group (XT), and amaranth addition group (AT). Each treatment was replicated three times. Tagetes erecta extract standardized to contain 2% (w/w) lutein as constituent and astaxanthin (1%, w/w) were purchased from Shanxi Huike Plant Development Co., Ltd., and 85% (w/w) amaranth leaf extract was purchased from Rouya (Shanghai) Food Additives Trade Co., Ltd. Based on preliminary experimental results from our laboratory and previous studies by Swian et al. [16], Biswas et al. [17], and Pailan et al. [18], dietary supplementation with 80–400 mg/kg lutein or astaxanthin extract for 4 weeks significantly improved fish body coloration. Three experimental groups were added pigment supplements: T. erecta extract 200 mg/kg, astaxanthin 200 mg/kg, and amaranth leaf extract 200 mg/kg. The control group was fed a basal diet without pigment supplement. During the breeding process, the pigment supplements were weighed daily and mixed with water and adhesive (purchased from Jiangsu Fishery Doctor Aquaculture Technology Co., Ltd.). The adhesive exhibits chemical inertness toward pigment additives and feed components. All experimental groups were fed the same basal diet. The growth experiment employed satiation feeding, with the first feeding at 5:00 AM and the second feeding at 4:30 PM. The formal experimental breeding cycle lasted 4 weeks. During the experiment, the water temperature ranged from 21.0 to 24.5°C, pH levels were maintained between 8.0 and 8.2, dissolved oxygen concentrations varied from 8.2 to 8.9 mg/L, and the salinity was approximately 27.9‰.
2.2. Sampling
After 28 days of culture, the test fish were subjected to a 24-h starvation period, followed by sample collection. The ventral skin of L. crocea exhibits a silvery-white coloration under light conditions, but turns golden-yellow when maintained in darkness [19]. Preliminary tests confirmed that the ventral skin coloration of L. crocea stabilizes after 40 min of dark exposure. Consequently, the collection procedure was divided into light and dark treatment groups. Seven individuals from each experimental enclosure were randomly chosen and anesthetized using a eugenol solution at a dilution ratio of 1:10,000. Data on weight, body length, and body height were recorded. Growth parameters were measured as follows:
Wt and W0 denote final body weight (FBW) and initial body weight (g), respectively, and t represents the culture period (days).
The light treatment group was sampled under natural light, while the dark treatment group was first acclimated in a dark room for 40 min before sampling. Continuous oxygen delivery in the dark treatment group ensured the activity of the fish. L^^∗^^, a^∗^, and b^∗^ values for ventral regions were measured. Additionally, 1 cm × 2 cm skin samples were taken from the control, AT, and LT groups under dark treatment conditions. These samples were placed in liquid nitrogen and preserved in ultralow temperature storage units maintained at −80°C. The skin pigment content in both the control and LT groups was measured and expression changes of differentially expressed genes (DEGs) and DEMs between the control and LT groups were assessed.
2.3. Body Color Determination
A portable spectrophotometer (CS-410, Chromatograph Technology, Zhejiang Co., Ltd.) was used to measure skin brightness (L^^∗^^), red–green (a^∗^), and yellow–blue (b^∗^) at ventral skin under both light (L) and dark (D) conditions. The average values for each measurement were calculated. These values were then used to compute five color coefficients: a^∗^/b^∗^, (a^∗^/b^∗^)2, chroma value (CV), and hue value (HV), according to the following calculation formulas:
2.4. Transcriptome Analysis
2.4.1. RNA Extraction
Total RNA from control group and LT group were performed utilizing TRlzol Reagent (Invitrogen, CA, USA). The purity and concentration of the extracted RNA were quantified with the NanoDrop 2000 spectrophotometer (Thermo Scientific, USA), and the integrity of the RNA was verified using the Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA)
2.4.2. Preparation of Sequencing Libraries and Assembling Transcriptomes
For the construction of RNA sequencing (RNA-seq) libraries, high-quality RNA samples (3 μg) were processed for RNA-seq library preparation. mRNA enrichment was achieved by capturing polyadenylated transcripts using magnetic beads conjugated with poly-T oligonucleotides. Construction of the RNA-seq libraries was carried out with the VAHTS Universal V6 RNA-seq kit, adhering to the standard operating procedures provided by the manufacturer. cDNA fragments within the size range of 400–500 bp were purified using the AMPure XP system (Beverly, California, USA). The Illumina PCR Primer Cocktail was utilized to amplify adapter-ligated DNA fragments through 15 rounds of PCR. Postpurification with the AMPure XP system, the library concentration was measured using the Agilent 2100 Bioanalyzer with a high-sensitivity DNA chip. The final libraries were sequenced using the Illumina NovaSeq 6000 system [20].
2.4.3. Transcriptome Sequencing
High-throughput sequencing was conducted on the NovaSeq 6000 platform (Illumina), generating bidirectional sequence fragments of 150 bp read length. Using the fastp software tool to subject quality control (QC) and preprocessing [21]. After filtering out substandard reads, the resulting high-quality data were aligned against the reference genome using the HISAT2 software for sequence mapping [22]. The level of gene expression was determined through the FPKM method [23] and read counts were generated through the application of the HTSeq-count software tool [24]. Q value < 0.05 and foldchange > 2 or foldchange < 0.5 identified using DESeq2 were considered as DEGs. Hierarchical clustering analysis of DEGs was implemented in R (version 3.2.0) to examine expression trends across distinct groups and samples. Furthermore, a radar plot depicting the top 30 DEGs was constructed using the ggradar, highlighting both upregulated and downregulated gene expression patterns.
2.4.4. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analyses
GO [25] and KEGG [26] pathway enrichment analysis of DEGs was carried out using the Reactome and WikiPathways resources, with the hypergeometric test applied for statistical evaluation in R (version 3.2.0). Significant terms were identified for each analysis. KEGG pathway analysis was conducted using Cluster Profiler (v 3.4.4) software to examine the functional pathways associated with the DEGs.
2.5. Quantitative Reverse Transcription PCR (qRT-PCR) Verification
To validate the RNA-Seq results, five genes were selected for a qRT-PCR analysis using a 2 × SYBR GREEN Master Mix. Primers were developed using sequence data obtained from the NCBI database and β-actin served as the internal control gene for normalization purposes. The thermal cycling conditions were as follows: 95°C for 10 min and 40 cycles at 95°C for 10 s and 60°C for 30 s. A melting curve was generated at the end of the amplification process. Primers for RT-PCR are shown in Table 1.
2.6. Metabolome Analysis
2.6.1. Sample Extraction
Reagents were precooled at −20 °C prior to the experiment. Samples from the control group and the LT group (30 mg) were weighed into EP tube and add two small steel balls. Then, 400 μL of a 4:1 (v/v) methanol–water mixture, which included a mixed internal standard at 4 μg/mL, was introduced. Precooling the mixture in a −40°C refrigerator for 2 min, the samples were ground using an automatic rapid grinding instrument (Wonbio-E, Shanghai Wanbai Biotechnology Co., Ltd.) at 60 Hz. The extraction process was carried out ultrasonically in an ice-water bath, followed by incubation at −40 °C for 2 h. The samples were then subjected to centrifugation at 13,000 rpm for 20 min at 4°C. A 150 μL aliquot was withdrawn using a syringe, passed through a 0.22 μm organic membrane filter, introduced into an LC via. Processed samples were preserved at −80 °C pending liquid chromatography/mass spectrometry (LC-MS) analysis. For quality assurance, a pooled QC sample was generated by combining equal volumes of extracts from each individual sample.
2.6.2. LS-MS Analysis
The samples were analyzed on an integrated LC-MS platform comprising a Waters ACQUITY UPLC I-Class Plus and a Thermo QE Plus high-resolution tandem mass spectrometer.
2.6.2.1. LC Conditions
The chromatographic column used was the ACQUITY UPLC HSS T3 (100 mm × 2.1 mm, 1.8 μm); column temperature: 45°C; injection volume: 3 μL; flow rate: 0.35 mL/min. The mobile phases consisted of A (water containing 0.1% formic acid) and B (acetonitrile). The gradient elution procedure is detailed in Table 2.
2.6.2.2. Mass Spectrometer Conditions
After T3 chromatography separation, primary and secondary spectra were acquired using the mass spectrometer.
2.7. Targeted Quantitative Detection of Lutein and Astaxanthin
2.7.1. Sample Pretreatment
Biological samples stored at −80 °C were ground to a fine powder using a low-temperature grinder (50 Hz, 1 min) in a liquid nitrogen environment. Approximately 100 mg of the ground sample (±5 mg) was weighed, mixed with 1.5 mL of acetone/n-hexane/ethanol (1:1:2, V/V/V) extraction solvent, and vortexed thoroughly. The extract was sonicated for 10 min using an ultrasonic device (KQ5200E, Kunshan Shumei Ultrasonic Instrument Co., Ltd.), then, incubated at 4°C for 16 h. The shaken sample solution was centrifuged at 12,000 rpm for 5 min. The supernatant was transferred to a new centrifuge tube and concentrated in a low-temperature cold trap (Auto R1-Plus, Beijing Jiam Instrument Co., Ltd.). After concentration, the residue was reconstituted in 100 μL of methanol/tert-butyl methyl ether (3:1, V/V), subjected to filtration using a 0.22 μm membrane, following which the filtrate was transferred into an injection vial.
2.7.2. LC-MS Conditions
2.7.2.1. LC Conditions
Chromatographic conditions: YMC Carotenoid C30 column (100 ×, 4.6 mm, 5 μm); temperature: 35 °C, volume of injection: 1 μL; flow rate: 0.3 mL/min; Phase A: 0.1% formic acid, Phase B: methanol; elution gradient: 0–2 min, A phase = 90%; 2–6 min, A phase = 90%–10%; 6–8 min, A phase = 10%; 8–8.1 min, A phase = 10%–90%; 8.1–10 min, A phase = 90%.
2.7.2.2. Mass Spectrometer Conditions
Ion source: ESI (Turbo Spray); polarity: positive; spray voltage: 5500 V; sheath gas: 30 psi; collision gas pressure: 9 psi; atomization temperature: 550°C; scanning mode: MRM. Targeted detection of compound ion pairs and collision voltage were shown in Table 3.
2.7.2.3. QC
In order to minimize the impact of instrument stability on test results, a quality assurance protocol was implemented. A QC sample was introduced following the analysis of 10 experimental samples. The total ion chromatogram (TIC) for the MS analysis of the control sample was overlaid to ensure data repeatability and reliability.
2.7.2.4. Standard Curve and Content Calculation
Standard solutions of varying concentrations were systematically prepared for method validation, and the ion abundance values corresponding to the target quantification ions of each concentration were measured and documented. Standard curves for different substances were generated by plotting the standard peak area against the standard concentration. The integral peak area ratios from all analyzed specimens were processed using the linear regression equation of the standard curve to determine the corresponding analyte concentrations. Further substitution into the formula yielded the substance content data for the actual samples. The content calculation formula is as follows:
c: The concentration value (ng/μL) applying the measured peak area ratio of the sample to the standard curve.
V: Volume (μL) of the sample reconstitution.
m: Weighed sample mass (g).
2.8. Statistical Analysis
All data were compiled and processed statistically using Microsoft Excel 2016. Prior to statistical analyses, normality was assessed using the Shapiro–Wilk test and homogeneity of variance evaluated via Levene's test. Growth performance, skin coloration, gene expression quantification, and targeted metabolomics data were analyzed by one-way ANOVA, with statistical significance set at p < 0.05. Results are presented as mean ± standard error of the mean (SEM).
3. Results
3.1. Changes in Body Color of Experimental Fish
Table 4 presents the growth performance of L. crocea fed diets supplemented with pigment additives for 28 days. Fish receiving pigment-enriched feeds showed no significant changes in growth performance compared to the control group. FBW, percent weight gain (PWG), and specific growth rate (SGR) remained statistically comparable (p > 0.05) between pigment-supplemented and basal diet-fed cohorts.
After the experiment, the ventral skin of the control group and three experimental groups were measured using a spectrophotometer under both light and dark conditions. The measurement positions are depicted in Figure 1. The CV and HV were subsequently calculated. The results are presented in Table 5 and Figure 2. Under dark conditions, the chroma of LT and AT increased at Locations A2 and A3. All three experimental groups exhibited HV to 0° (indicating redder color) at Locations A1 and A2 under dark conditions, with LT and AT showing similar trends in CV and HV changes. The XT group displayed minimal changes. Under light conditions, only LT group showed significantly higher chroma than the control group at Locations A1 and A2.
3.2. Illumina Sequencing and Quality Assessment
After count data were obtained, protein-coding genes were filtered to exclude those with zero counts across all samples. A total of 21,355 genes were detected, including 19,210, 19,358, 19,593, and 19,102 genes in control group. The LT group samples exhibited 19,744, 19,503, 19,056, and 18,853 detected genes, respectively. The number of genes detected in the two groups converged.
Principal component analysis (PCA) was conducted to assess the spatial arrangement and clustering characteristics of samples across different groups. The results are shown in Figure 3.
3.3. Analysis of DEGs
A total of 410 DEGs were identified between the control group and the LT group. Compared to the control group, 233 genes were upregulated and 177 genes were downregulated in the LT group. The radar plot of differential expression is shown in Figure 4, and the clustering heat map is presented in Figure 5. Notably, genes such as LOC104937478 (L-amino-acid oxidase, enables L-amino-acid oxidase activity), LOC104937491 (L-amino-acid oxidase-like, enables L-amino-acid oxidase activity), LOC104931550 (amine sulfotransferase, enables sulfotransferase activity), LOC113744190 (rab-3A-interacting protein-like, enables guanyl-nucleotide exchange factor activity), LOC104928767 (adhesion G protein-coupled receptor G2a, enables guanyl-nucleotide exchange factor activity), tnfrsf13b, and galnt9, among others, were significantly upregulated. In contrast, genes such as tyrp1b, ptprnb, dync1i1, LOC109137393 (Group 3 secretory phospholipase A2, enables phospholipase A2 activity), LOC104924650 (CD109 antigen, enables endopeptidase inhibitor activity), LOC113745247 (chromatin complexes subunit BAP18-like, promotes triple-negative breast cancer progression), and other were significantly downregulated.
3.3.1. GO Annotation of DEGs
Following differential gene analysis, DEGs were annotated using the GO database. DEGs between groups were performed according to genomic annotation information and comprehensive functional characterization. In the LT group, compared to the untreated controls, the upregulated GO terms in biological processes included leaf vascular tissue pattern formation and actin filament severing, while downregulated terms included response to jasmonic acid and response to hydrogen peroxide. In molecular function, upregulated GO terms included metallocarboxypeptidase activity and iron ion binding, while downregulated terms included inhibitor activity, 3′, 5′-cyclic-AMP phosphodiesterase (PDE) activity, and serine-type endopeptidase activity. In cellular components, upregulated GO terms included extracellular matrix and anchored component of the membrane, while downregulated terms included intracellular and extracellular regions. Figure 6 presents the GO enrichment analysis circle diagram, highlighting the top 20 classifications with the smallest p-values.
3.3.2. KEGG Pathway Analysis of Different Gene Expressions
A total of 122 KEGG pathways were significantly overrepresented in the DEGs between the LT and control groups. Of these, 66 pathways were upregulated, with eight pathways significantly upregulated, including phenylalanine, tyrosine, and tryptophan metabolic production, tyrosine metabolic pathway, and phenylalanine metabolic pathway. In contrast, 66 pathways were downregulated, with nine significantly downregulated, including melanogenesis and valine, leucine, and isoleucine biosynthesis. Additionally, 10 pathways exhibited partial upregulation and partial downregulation. The enrichment analysis of related differential expression pathways and key DEGs is presented in Figure 7.
3.4. Analysis of Differential Expressed Metabolites
In this study, a total of 3596 biochemical compounds were identified in the comparative analysis between experimental conditions and untreated controls, including 323 first-order metabolites, 1134 second-order metabolites, 165 third-order metabolites, and 1974 fourth-order metabolites. Compared to the control group, 99 metabolites, such as threonylalanine, citric acid, cetyl alcohol, and hypoxanthine, were significantly upregulated in the LT group, while 55 metabolites, including L-tyrosine, serine-glyoxylate, 4-aminoacetophenone, and isonicotinic acid, were significantly downregulated. The cluster heat map of the top 50 significant DEMs with the smallest p-values is shown in Figure 8.
3.4.1. KEGG Pathway Analysis of Differentially Expressed Metabolites
KEGG pathway enrichment analysis of the significant DEMs identified 18 upregulated pathways and 19 downregulated pathways. Upregulated pathways included signaling molecules and interaction, nucleotide metabolic pathway, amino acid metabolic pathway, and metabolic pathway of other amino acids. Downregulated pathways included pantothenate and CoA biosynthesis, beta-alanine metabolic pathway, tyrosine metabolic pathway, melanogenesis, gap junction, and thiamin metabolism, among others. The 20 pathways with the minimal p-values are listed in Figure 9.
3.5. Combined Transcriptomics/Metabolomics Analysis
Correlation analysis between DEGs and DEMs from transcriptomics and metabolomics identified the top 30 most significantly altered items, with their correlation coefficients calculated and visualized in a heatmap (Figure 10). DEGs and DEMs were mapped to the KEGG pathways, revealing their collective involvement in 14 pathways, including linoleic acid metabolism, alanine, tyrosine, tryptophan biosynthesis, tyrosine metabolism, melanogenesis, and phenylalanine metabolism, as listed in Table 6.
3.6. qRT-PCR Analysis
Genes associated with skin pigment synthesis, including scarb1, tyrp1b, mc1r, bco2, and UDP-glucuronosyltransferase 2A1, were selected for qRT-PCR validation based on high-throughput sequencing results. The qRT-PCR data experimentally validated the accuracy of the sequencing results. Gene expression levels are presented in Figure 11.
3.7. Analysis of Specific Content of Lutein and Astaxanthin in Larimichthys crocea Skin
The contents of lutein and astaxanthin in the ventral skin of L. crocea from both the control and experimental groups were measured using LC-MS. As shown in Table 7 and Figure 12, the LT group exhibited significantly higher levels of lutein and astaxanthin relative to the control group.
4. Discussion
Larimichthys crocea is highly prized by consumers for its tender flesh and rich nutritional value, making it a popular species in recent cage aquaculture practices. The color of this fish is a crucial determinant of its economic value [27]. However, the variation in breeding environments often leads to a decline in body color in farmed L. crocea, significantly reducing its market value. Studies have demonstrated that supplementing feeds with appropriate additives can enhance the body color of farmed animals [28–31]. Marigold, a traditional Chinese herbal medicine, is rich in lutein, which has been shown to not only improve body color but also enhance the immune function of farmed animals [32–35].
Dietary supplementation with three pigment additives can not significantly alter growth parameters in L. crocea over the 28-day trial. FBW, PWG, and SGR showed no statistically significant differences (p > 0.05). Short-term administration of pigment additives exerted no adverse effects on feeding behavior in L. crocea.
4.1. Effect of Tagetes erecta on Color Metrics in Larimichthys crocea
The L^^∗^^, a^∗^, and b^∗^ values are commonly employed to quantify fish coloration [36–38]. After 28 days of feeding L. crocea with three kinds of pigment supplements, ventral skin L^^∗^^, a^∗^, and b^∗^ values were measured using a spectrophotometer, and CV and HV were subsequently calculated. In comparison to the control and other treatment groups, the LT group exhibited significantly higher chroma and a more yellowish hue under various conditions. Based on these findings, the LT group was selected for further molecular-level investigations.
4.2. Tagetes erecta Regulates the Carotenoid Uptake and Metabolism Genes
Carotenoids, which share spectral similarities with pteridines, are responsible for producing red to yellow pigmentation in animals. These compounds are abundantly found in the red or yellow tissues of fish [39, 40]. In L. crocea, the yellow coloration of the body surface largely depends on carotenoid deposition.
The transport of carotenoids in the bloodstream is primarily facilitated by lipoproteins [41]. Among the key genes involved in carotenoid absorption and transport, ABCA1 plays a significant role, while SCARB1 is directly correlated to carotenoid pigmentation in vertebrate species, mediating their absorption and transport [42, 43]. Existing studies have indicated that both ABCA1 and SCARB1 are highly expressed in all-red Oujiang carp compared to their all-white counterparts [44]. The ABCA12 gene plays a crucial role in the translocation of lipid molecules, including ceramide, and plays a role in forming an extracellular lipid layer in the stratum corneum, contributing to skin barrier function [45]. In the present study, high-throughput sequencing revealed a significant upregulation of ABCA12 in the LT group (fold change = 2.7772, p = 0.0425) compared to the control group, with no significant difference observed in SCARB1 expression (fold change = 1.2501). However, qRT-PCR validation confirmed a slight upregulation of SCARB1 in the LT group (p = 0.0569), though not statistically significant. These results suggest that the more pronounced yellow coloration observed in the LT group may result from the upregulation of ABCA12 and SCARB1, which could enhance the transport, uptake, and deposition of carotenoids in the fish's skin.
Carotenoid oxygenases are crucial in carotenoid degradation, with three primary types: carotenoid isomerooxygenase (NinaB), β,β-carotene 15,15'-monooxygenase (BCMO), and β,β-carotene 9',10'-oxygenase (BCO2) [46]. BCO2 catalyzes the asymmetric cleavage of the 9'-10' double bond in the polyene backbone of carotenoids. It encodes a carotenoid lyase that mediates the cleavage of dietary carotenoids, preventing excessive carotenoid accumulation in the body, metabolizing lutein, and serving as an alternative pathway for retinoic acid synthesis [47–49]. In this study, qRT-PCR exhibited statistically significant decreases in BCO2 expression in the ventral skin of experimental fish in the LT group (fold change = 0.4798, p = 0.0015), suggesting a blockade in carotene degradation and subsequent deposition in the ventral skin.
Cytochrome P450 (CYP450) enzymes are extensively expressed across species and play key roles in pigment biosynthesis. For instance, cytochrome P450 Cit CYP97B regulates carotenoid diversity in citrus by hydroxylating β-cryptoxanthin [50]. Mundy et al. [51] demonstrated that the CYP 2J2-like gene cluster is essential for the coloration of red ketone carotenoids, while Xu et al. [52] proposed that CYP 2J2 promotes carotenoid deposition, particularly yellow pigments, via the arachidonic acid metabolic105and lipid pathways. CYP 26A1 is a retinoic acid-degrading enzyme [53–56], enhances melanin production [57]. In this study, high-throughput sequencing revealed significant upregulation of CYP 2J2 (fold change = 2.0239, p = 0.0384) and CYP 26A1 (fold change = 3.9715, p = 0.0119) in the LT group. These findings suggest that the enhanced yellow pigmentation in the LT group may result from increased carotenoid deposition due to upregulation of CYP 2J2, while the concurrent elevation of CYP 26A1 likely accelerates retinoic acid degradation, reducing melanin deposition.
4.3. Tagetes erecta Regulates the Melanogenesis and Metabolism Related Genes
Carotene coloration is influenced by melanocyte distribution [58]. Tyrosinase, a rate-limiting enzyme in controlling melanogenic, serves as a critical modulator of fish skin color. Previous studies have demonstrated a correlation between the activity of tyrosinase and the content of melanin in fish, with factors such as genetics, nutrition, differences in the environment, and developmental stages impacting this regulation [59–61]. Melanin production can also be inhibited by certain compounds that target the RAS/RAF/MAPK cascade or the cAMP–PKA–cAMP response element binding protein (CREB) signaling pathway [62–64]. cAMP modulates the activity of tyrosinase, the eumelanin-specific enzyme tyrosinase-related protein 1 (Tyrp1), and dopachrome tautomerase (Dct). Under normal conditions, these enzymes facilitate the deposition of eumelanin (brown to black pigment) in melanosomes. A decrease in intracellular cAMP levels shifts pigment production toward pheomelanin (yellow to red pigment) [65]. MC1R, expressed in melanocytes, activates intracellular cAMP levels, which in turn stimulates melanogenesis via α-MSH and the transcription of tyr, leading to increased melanin synthesis [66, 67]. PDE play a key role in regulating cAMP levels [68]. The level of intracellular cAMP is regulated by the antagonism of PDE, which can reduce cAMP into 5'-AMP [69, 70]. Transcription factors such as CREB and cAMP response element modulator (CREM) modulate gene transcription in response to increased cAMP levels. In this study, high-throughput sequencing revealed a significant reduction in MC1R expression (fold change = 0.4113, p = 0.0273), downregulation of pro-opiomelanocortin, which directs a-MSH synthesis (fold change = 0.4623, p = 0.0009), and a decrease in cAMP-specific 3',5'-cyclic PDE 4A (PDE4A) expression (fold change = 0.3300, p = 0.0060). The expression of high-affinity cGMP-specific PDE9A was significantly reduced (fold change = 0.4625, p = 0.0187). This reduction may be attributed to decreased MC1R expression, which impairs the normal activation of the cAMP signaling pathway, leading to reduced α-MSH synthesis. Consequently, the diminished demand for cAMP degradation results in downregulation of both PDE4A and PDE9A. Transcriptomic and qRT-PCR analyses also revealed a significant downregulation of tyrp1b (fold change = 0.3453, p = 0.0128). This suggests that the cAMP pathway's downregulation fails to activate the key melanin synthesis gene, tyrp1b, leading to reduced eumelanin expression on the body surface, increased pheomelanin production, and a visible alteration in the body color of L. crocea. Additionally, the significant upregulation of cAMP response element modulator a (crema; fold change = 2.3704. *p<*0.0010) may help compensate for the reduced cAMP levels, thus, balancing transcriptional responses.
4.4. Effect of Tagetes erecta in Larimichthys crocea: Dual Regulation of Citrate and Tyrosine Production
Citric acid, an intermediate in the tricarboxylic acid (TCA) cycle, plays a pivotal role in energy metabolism, macromolecular synthesis, and maintaining cellular redox balance [71]. Tributyl citrate has demonstrated antimelanin activity [72], and citric acid has been identified as a depigmenting agent in topical treatments for hyperpigmentation [73]. To validate the melanogenesis-inhibitory properties of succinic and citric acids, multiple studies have used B16-F10 cells to evaluate cell viability and melanin content. These studies demonstrated that treatment with succinic and citric acids resulted in a reduction of melanogenic activity [74].
In the nontargeted metabolomic analysis of this study, citric acid was significantly upregulated as a DEM (fold change = 1.7589), which may be attributed to the addition of lutein in the feed. This increase in citric acid synthesis in L. crocea could inhibit melanin production. L-tyrosine, a precursor of catecholamines and melanin, is hydroxylated by tyrosinase to L-dihydroxyphenylalanine (L-DOPA), which is further oxidized to dopaquinone and then to melanin [75]. Tyrosine levels may influence the transition of melanocytes from a proliferative to a melanin-synthesizing and terminally differentiated state [76]. In this study, L-tyrosine was significantly downregulated (fold change = 0.6793), which likely blocked melanin synthesis in the skin of L. crocea, enhancing the prominence of yellow pigments on the skin.
4.5. Transcriptome-Metabolome Analysis: Tagetes erecta Improve Skin Pigmentation in Larimichthys crocea Through Tyrosine and Melanin Production Pathways
Mapping DEGs identified by high-throughput sequencing and metabolites detected by nontargeted metabolomics to KEGG pathways revealed that lutein upregulated key enzymes involved in tyrosine-related metabolism (Figure 13), such as L-amino acid oxidase [EC: 1.4.3.2] (genes LOC1049374780 and LOC104937491). This upregulation accelerates the conversion of tyrosine away from eumelanin synthesis. Simultaneously, downregulation of TYRP1 expression inhibited eumelanin production, thus, reinforcing the yellow pigmentation characteristic on the surface of L. crocea.
In the melanin production pathway (Figure 14), significant downregulation of MC1R, pro-opiomelanocortin (LOC104921984), which guides the synthesis ofα-MSH, as well as Wnt10a, Tyrp1, and tyrosine-key regulators of melanin synthesis-resulted in a reduction of the entire melanin biosynthesis pathway (p = 0.0231). This led to decreased melanin synthesis and deposition, thereby accentuating the yellow pigment on the surface of L. crocea.
5. Conclusions
This study concludes that T. erecta supplementation significantly enhances the body color of L. crocea, compared with the astaxanthin feeding group and the amaranth feeding group, the T. erecta feeding group had the best effect on enhancing the yellow skin of large yellow croaker. The potential mechanism may involve the upregulation of carotenoid deposition and the downregulation of the expression of tyrosine, an important precursor in the melanin formation pathway. Therefore, the contribution of carotenoids to coloring became more prominent, thereby improving the body color of L. crocea. Critically, our findings demonstrate that dietary pigment additives, particularly lutein from T. erecta, enhance golden skin coloration in L. crocea, directly increasing market value.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Suo N. Wu Y. Zhou Z. Genome-Wide Association and Expression Analysis Revealed the Candidate Variants and Molecular Underpinnings of Cold-Stress Response in Large Yellow Croaker Marine Biotechnology 202224592794110.1007/s 10126-022-10155-735971020 · doi ↗ · pubmed ↗
- 2Mao J. Fu J. Qi X. Chen Y. Zhang B. Effect of Theaflavins on the Quality of Large Yellow Croaker, Larimichthys Crocea, During Refrigerated Storage Journal of the Science of Food and Agriculture 2023103126033604310.1002/jsfa.1267137119403 · doi ↗ · pubmed ↗
- 3Sun P. Bao P. Tang B. Transcriptome Analysis and Discovery of Genes Involved in Immune Pathways in Large Yellow Croaker (Larimichthys Crocea) Under High Stocking Density Stress Fish and Shellfish Immunology 20176833234010.1016/j.fsi.2017.07.0132-s 2.0-8502581767328698122 · doi ↗ · pubmed ↗
- 4Ma R. Meng Y. Zhang W. Mai K. Comparative Study on the Organoleptic Quality of Wild and Farmed Large Yellow Croaker Larimichthys Crocea Journal of Oceanology and Limnology 202038126027410.1007/s 00343-019-8353-02-s 2.0-85071872953 · doi ↗
- 5Yi X. Shen H. Li J. Effects of Dietary Vitamin E and Astaxanthin on Growth, Skin Colour and Antioxidative Capacity of Large Yellow Croaker Larimichthys Crocea Aquaculture Nutrition 201824147248010.1111/anu.125802-s 2.0-85021206600 · doi ↗
- 6Zheng J. L. Wan F. G. Chen Y. L. Comparative Study on the Morphological Characteristics, Nutritional Quality, and Tastes of Large Yellow Croaker From Five Cage Culture Areas: Relay Farming Improved Fish Quality Aquaculture 202459010.1016/j.aquaculture.2024.741030741030 · doi ↗
- 7Nord H. Dennhag N. Muck J. Hofsten J. V. Pax 7 Is Required for Establishment of the Xanthophore Lineage in Zebrafish Embryos Molecular Biology of the Cell 20162711170718742705365810.1091/mbc.E 15-12-0821 PMC 4884075 · doi ↗ · pubmed ↗
- 8Torrissen O. Hardy R. Shearer K. Scott T. Stone F. E. Effect of Dietary Canthaxanthin Level and Lipid Level on Apparent Digestibility Coefficients for Canthaxanthin in Rainbow Trout (Oncorhynchus Mykiss) Aquaculture 1990883-435136210.1016/0044-8486(90)90160-O 2-s 2.0-38249016407 · doi ↗