When opposites compensate: proton flux crosstalk between envelope antiporters and ATP synthase
Ritu Singh

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
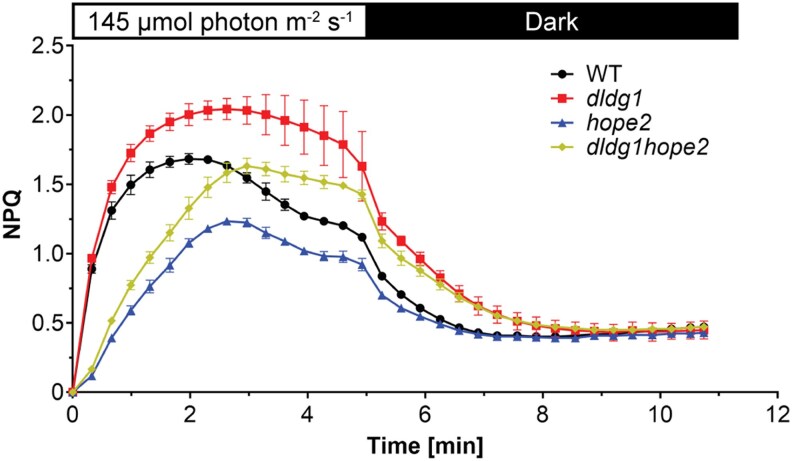 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsATP Synthase and ATPases Research · Fuel Cells and Related Materials · Advanced MRI Techniques and Applications
Photosynthesis underpins life on Earth by converting solar energy into the chemical energy that sustains nearly all food webs. In plants, photons are absorbed by chloroplasts and funneled to the photosystem reaction centers. Electrons excited in PSII travel through the electron transport chain to PSI, producing NADPH, while the cytochrome b6f complex pumps protons into the thylakoid lumen to establish a proton-motive force (pmf). ATP synthase harnesses this pmf to generate ATP, and together ATP and NADPH fuel the Calvin–Benson cycle (Blankenship 2002). However, fluctuating environments frequently challenge this delicate energy balance. When absorbed light energy exceeds the capacity for carbon fixation, the electron transport chain becomes over-reduced, leading to the production of reactive oxygen species. To prevent such damage, plants employ photoprotective strategies, most prominently nonphotochemical quenching (NPQ). The rapid, energy-dependent component of NPQ (qE) is activated by lumen acidification (ΔpH), which drives the xanthophyll cycle and protonates the PSII protein subunit S to safely dissipate excess energy as heat (Ruban 2016).
Maintaining chloroplast energy balance requires precise regulation of proton and ion gradients across both thylakoid and envelope membranes. While ATP synthase provides the main route for proton efflux from the lumen, chloroplast envelope proteins also influence lumen pH and photoprotection. One such envelope protein, DAY-LENGTH-DEPENDENT DELAYED-GREENING1 (DLDG1), is a putative K⁺(Ca²⁺)/H⁺ antiporter whose loss in Arabidopsis thaliana causes lumen over-acidification and elevated NPQ (Harada et al. 2019; Suzuki and Masuda 2023). In contrast, the hope2 mutant, harbouring a single amino acid change in the ATP synthase γ-subunit, shows defective regulation of proton conductivity (gH⁺) and reduced NPQ. These contrasting phenotypes raised the possibility that DLDG1 and ATP synthase might work together to fine-tune proton flux and optimize photoprotection (Takagi et al. 2017; Degen et al. 2023).
In a recent Plant Physiology study, Trinh and colleagues (Trinh et al. 2025) investigated the genetic interplay between the envelope transporter DLDG1 and the chloroplast ATP synthase by creating a dldg1hope2 double mutant. Plants were grown under short-day conditions, and NPQ kinetics were measured across light–dark transitions. A striking partial compensation emerged: the slow, diminished NPQ response of hope2 was substantially restored in the double mutant, particularly during the first minutes of illumination under low and moderate actinic light. NPQ levels in the double mutant approached those of the dldg1 single mutant, which is known to exhibit elevated photoprotective quenching.
Since the enhanced NPQ of dldg1 requires both ΔpH and PSII protein subunit S activity (Harada et al. 2019; Suzuki and Masuda 2023), the authors hypothesized that the rescue observed in the double mutant reflects enhanced ΔpH formation supplied by the dldg1 background. To probe this mechanism, the authors measured proton conductivity using the electrochromic shift (ECS), a technique that assesses the pmf. The result showed a balancing effect: while hope2 displayed elevated H+ conductivity (gH⁺) and dldg1 exhibited reduced gH⁺ and increased pmf, the double mutant had (gH⁺) values closer to wild type. Moreover, the characteristic pmf increase of dldg1 was absent in the double mutant, supporting the idea of mutual compensation in proton flux. Importantly, western-blot analyses detected no changes in the redox status of the ATP synthase γ-subunit, suggesting that this genetic interaction arises from metabolic adjustments rather than a redox switch.
Since photosystem performance is sensitive to environmental conditions, the authors also compared mutant responses under varying CO₂ conditions and ambient air. Under low CO_2_ conditions, no significant differences in PSII electron transport were observed between wild-type and mutant plants. However, at moderate (400 ppm) and high (1500 ppm) CO_2_, hope2 mutants showed significantly lower PSII electron transport than dldg1. This indicates that dldg1 and hope2 mutations exert opposing effects on photosynthetic electron transport under carbon-sufficient conditions. Notably, dldg1hope2 double mutant did not significantly differ from dldg1, indicating that the hope2-associated reduction in photosynthetic electron transport activity may be partially complemented by the dldg1 mutation. Similarly, under ambient air, compared with wild type, hope2 plants showed reduced NPQ induction and elevated electron pressure at PSII, while these defects were partially alleviated in the double mutant. Photosynthetic performance at both PSII and PSI therefore supports the conclusion that introducing the dldg1 null mutation into the hope2 background provides partial functional compensation, mitigating the impaired photoprotection and electron transport of hope2.
Would this biochemical compensation benefit plant growth? To address this, the authors compared biomass and photoinhibition under short-day, long-day, constant-light, and fluctuating-light conditions that revealed a more complex picture. Under short days, dldg1 plants resembled wild type, hope2 accumulated slightly less biomass in line with reduced CO₂ assimilation, and the double mutant showed only a modest improvement. In long days or constant light, both dldg1 and dldg1hope2 developed transient pale-green leaves with reduced maximum PSII efficiency (Fv/Fm) that recovered upon maturation. Most strikingly, under fluctuating light, hope2 plants were dwarfed and photoinhibited, and the double mutant performed even worse, with a pronounced drop in Fv/Fm. These findings underscore that compensation at the photosystem level does not guarantee enhanced growth under dynamic environments and may even compromise fitness. Interestingly, the authors found that the pale-green phenotype of dldg1 (and dldg1hope2) under constant light was partially alleviated by NaCl supplementation, paralleling the behavior of KEA1/2 antiporter mutants and hinting at shared roles in stromal ion homeostasis.
Collectively, these findings support a model in which DLDG1 influences stromal pH and ionic balance in ways that modulate ATP synthase activity and thereby affect lumen acidification and qE (Fig. 1). In hope2, defective metabolic regulation of ATP synthase leads to excessive proton efflux from the lumen; removing DLDG1 shifts stromal conditions to restrict this efflux, restoring ΔpH and enabling NPQ induction. Whether this interplay reflects a direct functional partnership or an indirect effect mediated by other transporters such as KEA1/2 or rapid-activated chloroplast ion channels remains an open and exciting question.
This study highlights the remarkable plasticity of chloroplast ion transport networks, where mutations in seemingly distinct components can compensate for one another at the biochemical level. Yet it also underscores the challenge of scaling from molecular adjustments to whole-plant fitness: what stabilizes photoprotection under controlled light may not guarantee improved performance in fluctuating natural environments.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Blankenship RE . Molecular mechanisms of photosynthesis. John Wiley & Sons; 2002. 10.1002/9780470758472 · doi ↗
- 2Degen GE, Jackson PJ, Proctor MS, Zoulias N, Casson SA, Johnson MP. High cyclic electron transfer via the PGR 5 pathway in the absence of photosynthetic control. Plant Physiol. 2023:192(1):370–386. 10.1093/plphys/kiad 08436774530 PMC 10152662 · doi ↗ · pubmed ↗
- 3Harada K, Arizono T, Sato R, Trinh MDL, Hashimoto A, Kono M, Tsujii M, Uozumi N, Takaichi S, Masuda S. DAY-LENGTH-DEPENDENT DELAYED-GREENING 1, the Arabidopsis homolog of the cyanobacterial H+-extrusion protein, is essential for chloroplast p H regulation and optimization of non-photochemical quenching. Plant Cell Physiol. 2019:60(12):2660–2671. 10.1093/pcp/pcz 20331665522 · doi ↗ · pubmed ↗
- 4Ruban AV . Nonphotochemical chlorophyll fluorescence quenching: mechanism and effectiveness in protecting plants from photodamage. Plant Physiol. 2016:170(4):1903–1916. 10.1104/pp.15.0193526864015 PMC 4825125 · doi ↗ · pubmed ↗
- 5Suzuki K, Masuda S. Arabidopsis mutants lacking DLDG 1 and non-photochemical quenching-related proteins reveal the regulatory role of DLDG 1 in chloroplast p H homeostasis. FEBS Lett. 2023:597(13):1761–1769. 10.1002/1873-3468.1468737339934 · doi ↗ · pubmed ↗
- 6Takagi D, Amako K, Hashiguchi M, Fukaki H, Ishizaki K, Goh T, Fukao Y, Sano R, Kurata T, Demura T, et al Chloroplastic ATP synthase builds up a proton motive force preventing production of reactive oxygen species in photosystem I. Plant J. 2017:91(2):306–324. 10.1111/tpj.1356628380278 · doi ↗ · pubmed ↗
- 7Trinh MDL, Esmailpourmoghadam E, Sato R, Miyake C, Palmgren M, Masuda S. Stromal p H regulation by the envelope-localized DLDG 1 modulates H+ translocation across thylakoid membranes via plastidial ATP synthase. Plant Physiol. 2025:199(1):kiaf 373. 10.1093/plphys/kiaf 37340857592 · doi ↗ · pubmed ↗