Association of a Novel IgG3 Allele With Malaria in Children From the Sepik Region of Papua New Guinea
Maria Saeed, Elizabeth H Aitken, Myo T Naung, Caitlin Bourke, Kenneth W Wu, Rhea J Longley, Amy W Chung, Timon Damelang, Benson Kiniboro, Ivo Mueller, Stephen J Rogerson

TL;DR
A new IgG3 antibody variant is found to be common in children in Papua New Guinea and is linked to fewer malaria infections, especially from Plasmodium vivax.
Contribution
The discovery of a novel IgG3 allele (IGHG3*30) associated with reduced malaria infections in children from East Sepik, Papua New Guinea.
Findings
78% of children in the study had the IGHG3*30 allele, which is associated with fewer Plasmodium infections.
Children with IGHG3*30 had on average one fewer Plasmodium vivax infection over 18 months.
IGHG3*30 carriers had lower IgG levels to P. vivax vaccine candidate proteins.
Abstract
Susceptibility to malaria can be influenced by host genetic factors, including immune response genes. Antibodies against Plasmodium antigens are known to play an important role in protection from clinical disease. Polymorphisms in these antibodies may result in different functional properties that could provide protection from malaria. Immunoglobulin G1 (IgG1) and immunoglobulin G3 (IgG3) alleles and IgG3 hinge region were investigated by polymerase chain reaction and Sanger sequencing in a longitudinal cohort of children aged 1–3 years (N = 203) from the East Sepik region of Papua New Guinea. Linear regression was used to investigate associations between immunoglobulin alleles and Plasmodium infections. Seventy-eight percent of the children were either heterozygous (n = 82 [40%]) or homozygous (n = 77 [38%]) for IGHG3*30 (G3m29), a novel IgG3 allele. G3m29 has a long hinge region of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
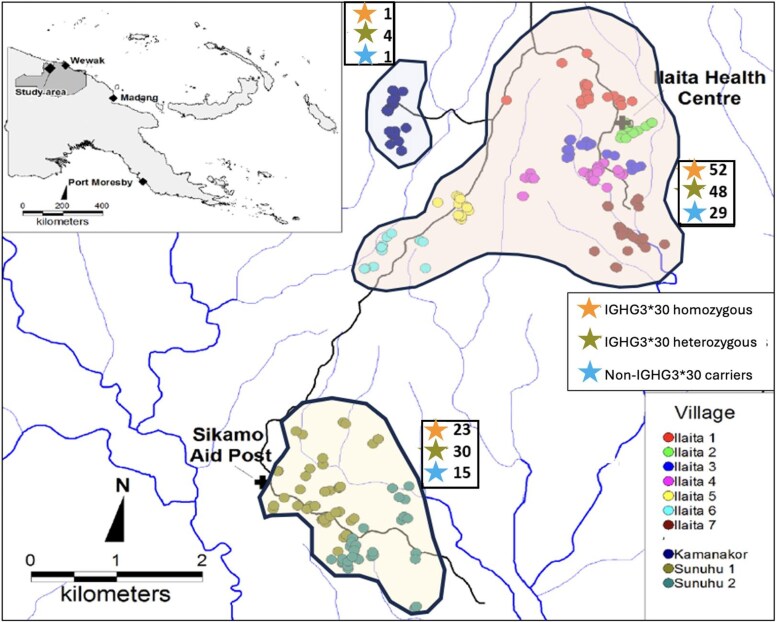 Figure 1
Figure 1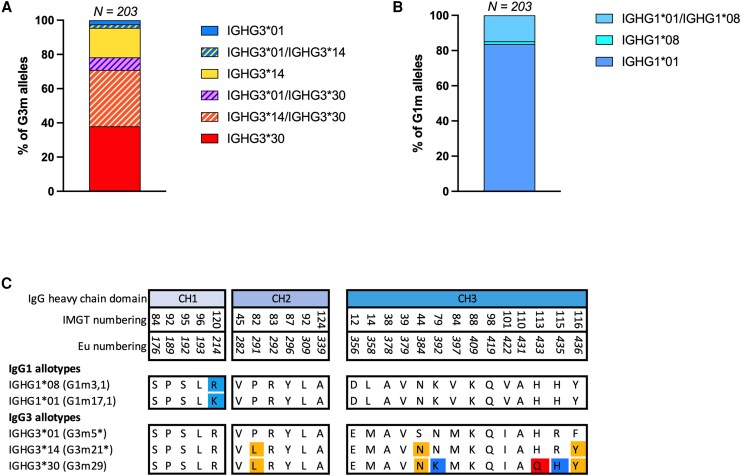 Figure 2
Figure 2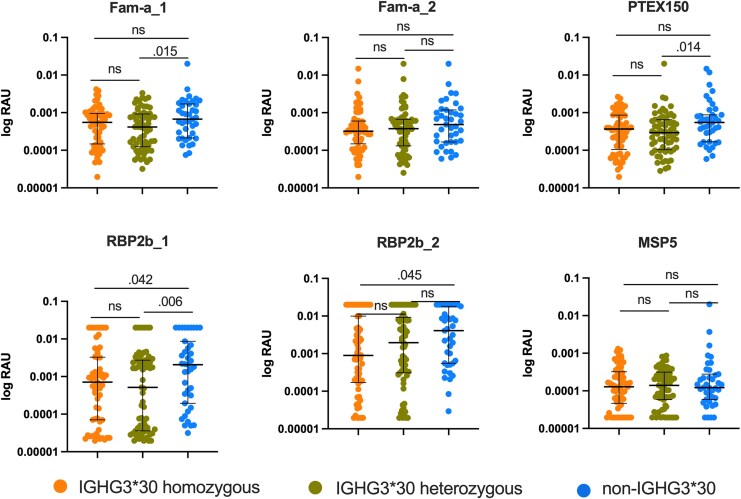 Figure 3
Figure 3| Characteristics | Total | IGHG3*30 | Non-IGHG3*30 |
|
|---|---|---|---|---|
| Age, mo, mean ± SD | 23.5 ± 8.2 | 22.9 ± 8.1 | 25.4 ± 8.3 | .074 |
| Hemoglobin, g/dL, mean ± SD | 9.0 ± 0.95 | 9.0 ± 0.98 | 9.0 ± 0.82 | .967 |
| Sex | ||||
| Male | 117 (57.63) | 94 (59.12) | 23 (52.27) | .416 |
| Female | 86 (42.36) | 65 (40.88) | 21 (47.72) | .416 |
| Village | ||||
| Ilaita 1 | 20 (9.85) | 15 (9.43) | 5 (11.36) | .703 |
| Ilaita 2 | 12 (5.91) | 9 (5.66) | 3 (6.82) | .725 |
| Ilaita 3 | 20 (9.85) | 15 (9.43) | 5 (11.36) | .703 |
| Ilaita 4 | 27 (13.30) | 23 (14.46) | 4 (9.09) | .456 |
| Ilaita 5 | 13 (6.40) | 9 (5.66) | 4 (9.09) | .484 |
| Ilaita 6 | 12 (5.91) | 11 (6.92) | 1 (2.27) | .468 |
| Ilaita 7 | 25 (12.31) | 18 (11.32) | 7 (15.91) | .412 |
| Kamanakor | 6 (2.95) | 5 (3.14) | 1 (2.27) | .999 |
| Sunuhu 1 | 43 (21.18) | 33 (20.75) | 10 (22.73) | .778 |
| Sunuhu 2 | 26 (12.81) | 21 (13.20) | 5 (11.36) | .746 |
| Total No. of | 12.5 ± 4.95 | 12.1 ± 5.1 | 13.9 ± 4.3 |
|
| Allele | No. of Children | LL (806 bp) | SS (430 bp) | LS (806/430 bp) |
|---|---|---|---|---|
| IGHG3 allele | ||||
| IGHG3*01 | 5 | 5 | … | … |
| IGHG3*14 | 35 | 32 | 3 | … |
| IGHG3*30 | 77 | 77 | … | … |
| IGHG3*01/IGHG3*14 | 4 | 4 | … | … |
| IGHG3*01/IGHG3*30 | 15 | 15 | … | … |
| IGHG3*14/IGHG3*30 | 67 | 64 | … | 3 |
| IGHG1 allele | … | … | … | |
| IGHG1*01 | 170 | … | … | … |
| IGHG1*01/IGHG1*08 | 30 | … | … | … |
| IGHG1*08 | 3 | … | … | … |
| Infection | Coefficient | (95% CI) |
|
|---|---|---|---|
|
| −1.73 | (−3.39, −.08) |
|
| Asymptomatic | −1.60 | (−2.89, −.46) |
|
|
| −1.06 | (−2.01, −.12) |
|
|
| −0.19 | (−1.04, .63) | .640 |
| Symptomatic | −0.49 | (−1.32, .34) | .246 |
|
| 0.03 | (−0.63, .70) | .915 |
|
| −0.50 | (−1.09, .08) | .094 |
|
| −0.09 | (−.20, .02) | .115 |
|
| −0.09 | (−.22, .03) | .125 |
| Infection | Coefficient | (95% CI) |
|
|---|---|---|---|
|
| |||
| Gm IGHG1*01, IGHG3*30 heterozygous | −2.95 | (−5.35, −.55) |
|
| Gm IGHG1*01, IGHG3*30 homozygous | −0.14 | (−1.61, 1.31) | .842 |
| Asymptomatic | |||
| Gm IGHG1*01, IGHG3*30 heterozygous | −2.387 | (−4.15, −.061) |
|
| Gm IGHG1*01, IGHG3*30 homozygous | −0.37 | (−1.45, .71) | .496 |
|
| |||
| Gm IGHG1*01, IGHG3*30 heterozygous | −1.43 | (2.81, −.05) |
|
| Gm IGHG1*01, IGHG3*30 homozygous | −0.05 | (−.89, .79) | .901 |
|
| |||
| Gm IGHG1*01, IGHG3*30 heterozygous | −0.65 | (−1.87, .56) | .291 |
| Gm IGHG1*01, IGHG3*30 homozygous | 0.08 | (−.65, .83) | .815 |
| Symptomatic | |||
| Gm IGHG1*01, IGHG3*30 heterozygous | −0.62 | (−1.84, .58) | .308 |
| Gm IGHG1*01, IGHG3*30 homozygous | −0.17 | (−.91, .57) | .650 |
|
| |||
| Gm IGHG1*01, IGHG3*30 heterozygous | 0.20 | (−.76, 1.17) | .677 |
| Gm IGHG1*01, IGHG3*30 homozygous | 0.34 | (−.24, .93) | .246 |
|
| |||
| Gm IGHG1*01, IGHG3*30 heterozygous | −0.79 | (−1.64, .05) | .065 |
| Gm IGHG1*01, IGHG3*30 homozygous | −0.52 | (−1.04, −.00) |
|
|
| |||
| Gm IGHG1*01, IGHG3*30 heterozygous | −0.14 | (−.31, .02) | .094 |
| Gm IGHG1*01, IGHG3*30 homozygous | 0.01 | (−.08, .11) | .796 |
|
| |||
| Gm IGHG1*01, IGHG3*30 heterozygous | −0.124 | (−.31, .060) | .187 |
| Gm IGHG1*01, IGHG3*30 homozygous | 0.020 | (−.092, .13) | .725 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGlycosylation and Glycoproteins Research · HIV Research and Treatment · Complement system in diseases
Malaria is a life-threatening disease affecting millions of people worldwide. In 2023, there were 597 000 malaria-related deaths worldwide, mostly in children <5 years of age [1]. Malaria has unarguably altered the human genome and has been a driving force behind evolutionary selection [2]. Red blood cell polymorphisms such as sickle cell anemia, thalassemia, glucose-6-phosphate dehydrogenase deficiency, and ovalocytosis are reported to be protective against severe malaria and are widely distributed in malaria-endemic regions [3]. Acquisition of malaria-specific antibodies is associated with protection from clinical malaria [4, 5], and antibody features like Fc-mediated effector functions have the potential to control malaria [6, 7].
Antibody allotypes on the heavy chain of immunoglobulin G (IgG) 1, IgG2, IgG3, and IgG4 constant region are written as G1m, G2m, G3m, and G4m, respectively. IgG3 is the most polymorphic subclass with 15 reported allotypes, mostly occurring in the C_H_2 and C_H_3 domains [8, 9]. These polymorphisms may have structural and/or functional consequences, including changes in length of the hinge region, antibody half-life, and Fc-mediated effector functions [9, 10]. IgG3 allotypes G3m15* and G3m16* encoded by IGHG317, IGHG318, IGHG319, IGHG322, and IGHG323 have arginine to histidine substitutions at the 435 position (p. Arg435His) in the C_H_3 domain, which are associated with increased transplacental IgG3 transfer [10–12], and infants of G3m15 and/or G3m16* allele-carrying mothers have less clinical malaria [12]. Some IgG3 allotypes alter binding affinities for effector molecules, such as Fcγ receptors (FcγRs) or C1q, which can influence effector functions such as antibody-dependent cellular phagocytosis (ADCP), cellular cytotoxicity, and complement activation [13–17]. Therefore, it is imperative to investigate the significance of G3m allotypes in malaria.
Recently a novel IgG3 allele, hereafter referred to as IGHG330 (G3m29), was reported in a cohort of pregnant women from the Sepik region in Papua New Guinea (PNG) [18]. IGHG330 is characterized by a novel substitution of histidine to glutamine at the 433 position (p. His433Glu) in the C_H_3 domain [18]. This position has not been described as a polymorphic position before. Functional amino acid substitutions include Pro291Leu, Ser384Asp, Arg435His, and Phe436Tyr [18]. IGHG330 also has an Arg392Lys substitution whose functional significance is unclear [19]. IGHG330 has altered functions and interactions with FcγRs. Plasma from women with the IGHG330 allele was able to induce significantly higher levels of ADCP but not significantly different antibody-dependent neutrophil phagocytosis (ADNP) of Plasmodium falciparum–infected erythrocytes compared to plasma from non-IGHG330 allele–carrying women. Moreover, monoclonal antibodies made with IGHG3*30 heavy chains showed increased affinity to FcγRIIa and significantly higher ADCP of P falciparum VAR2CSA DBL3-coated beads and 2-fold higher ADNP of P falciparum–infected erythrocytes as compared to other Gm allotypes [18].
Here we examine the prevalence and potential protective role of IGHG330 allele, in a cohort of 203 young PNG children followed longitudinally for Plasmodium spp infection and febrile illness over a period of 18 months [20]. The G1m and G3m allotypes of the children were determined and associations with number of malaria infections were examined. We also measured the length of the hinge region and compared levels of IgG against a panel of Plasmodium vivax antigens in children with and without the IGHG330 allele.
MATERIALS AND METHODS
Study Design and Sample Collection
The cohort details and sample collection protocols have been described previously [20]. In brief, in a longitudinal study, 264 PNG children aged 1–3 years from 11 villages in the Illaita area of Maprik District, East Sepik Province, PNG (Figure 1), were recruited in March 2006 and actively followed up for 18 months for Plasmodium spp infection and febrile illness [20]. Plasmodium spp infection was confirmed by light microscopy and polymerase chain reaction (PCR) [21]. DNA extracted from blood samples (N = 203) was available for this study to investigate IgG1 and IgG3 alleles and their association with Plasmodium spp infections. Baseline characteristics of the study participants are shown in Table 1.
Map of study area. Adapted from Lin et al, 2010 [20]. Dots represent the participant's houses, crosses represent health centers, black lines represent roads, and blue lines represent rivers and streams. Stars represent the number of homozygous IGHG330 allele carriers (top), heterozygous IGHG330 allele carriers (middle), and non-IGHG330 allele carriers (bottom).*
Definition of Categories
Plasmodium spp infections were identified and categorized (Supplementary Table 1) [20–22]. The summary of Plasmodium spp infections in children is shown in Supplementary Table 2.
Ethics Approval
Ethical approval was obtained from institutional review boards of the PNG Medical Research Advisory Committee (approval 05.19), University Hospitals Case Medical Center (Cleveland, Ohio, USA), and the Swiss Tropical Institute. Written informed consent was obtained from parent(s) or guardian(s) before recruitment. The Walter and Eliza Hall Institute of Medical Research human research ethics committee (07/07) provided the ethical approval for samples to be used for antibody investigations.
Polymerase Chain Reaction
Genomic DNA was enriched using Illustra GenomiPhi V2 DNA amplification kit, Cytiva (GE Healthcare Life Sciences, Australia) as per the manufacturer's guidelines. PCR was optimized to amplify the genes encoding C_H_2 and C_H_3 domains of IgG3 [23], IgG1-C_H_1 [24], and IgG1-C_H_3 domains [25] and genes encoding IgG3 hinge [26]. Primer sequences are provided in Supplementary Table 3.
For the C_H_2 and C_H_3 domains of IgG3, 30- μL PCR reactions were prepared containing 2–5 ng of enriched DNA, 1X AccuPrime PCR buffer II, 0.6 μL of 100% dimethyl sulfoxide, 1.2 μL of each primer (0.5 μM), and 0.15 μL of AccuPrime Taq polymerase (Thermo Fisher). For the C_H_1 and C_H_3 domains of IgG1, 30-μL PCR reactions were prepared containing 2–5 ng of enriched DNA, 1X AccuPrime PCR buffer II, 0.6 μL of each primer (0.5 μM), and 0.15 μL of AccuPrime Taq polymerase. PCR conditions for the 2 reactions included 94°C for 30 seconds (initial denaturation), 94°C for 30 seconds (denaturation), 60°C for 30 seconds (annealing), 68°C for 1 minute (extension), and 35 cycles of amplification followed by 72°C for 7 minutes (final extension) on a BioRad T100 Thermal Cycler [18]. For the IgG3 hinge region, 25 μL PCR reaction was prepared containing 2–5 ng of enriched DNA sample, 1X HotStar buffer, 1 μL of each primer (1 μM), and 1.25 U of HotStarTaq polymerase (QIAGEN). HotStarTaq DNA polymerase was activated at 95°C for 15 minutes, then 38 cycles of 95°C for 30 seconds, 61°C for 30 seconds, and 72°C for 30 seconds, and final extension at 72°C for 7 minutes were performed [26].
PCR products were visualized on a 2% agarose gel in 1X TAE buffer at 110 V for 1 hour (Supplementary Figure 1). For identification of single-nucleotide polymorphisms, PCR products were submitted to the Australian Genome Research Facility (Victorian Comprehensive Cancer Centre, Parkville) for Sanger sequencing. Sequences were analyzed in Geneious Prime software (version 2024.0.5). For hinge length estimation, band sizes of 806, 618, and 430 bp corresponding to long (L), medium (M), and small (S) hinge phenotypes were identified.
Total IgG Levels to P vivax Proteins
Six P vivax antigens (fam-a_1, fam-a_2, RBP2b_1 [reticulocyte binding protein 2b], RBP2b_2, PTEX150 [Plasmodium translocon of exported proteins], and MSP5 [merozoite surface protein 5]) were selected because they were previously described as top serological markers of exposure [27]. There is high level of genetic diversity in Plasmodium antigens, so 2 commonly found haplotypes of P vivax fam-a and RBP2b were included. Proteins were coupled to BioRad COOH magnetic beads (BioRad, California, USA) according to the manufacturer's guidelines. Plasma samples collected at enrollment from study participants were diluted 1:100 and added to coupled beads to measure total IgG using multiplex Luminex system as previously described [27]. Raw mean fluorescence intensities were converted to relative antibody units using a 5-parameter logistic regression standard curve generated from the hyperimmune positive control plasma pool from PNG using RshinyAPP (https://shaziaruybal.shinyapps.io/COVIDClassifyR/).
Data Analysis
Data were analyzed using Stata17 and Prism version 10 (GraphPad Software). Associations between IGHG330 and Plasmodium spp infections were measured via linear regression. Associations between G1m and G3m allotypes were investigated using χ^2^ test. Differences in the antibody levels between individuals with and without IGHG330 were assessed by the nonparametric Mann-Whitney U test and Kruskal-Wallis test. Multiple linear regression was performed to assess the associations between IGHG3*30 allele carrier status and Plasmodium spp infections after adjusting for IgG levels to 6 P vivax proteins.
RESULTS
Novel IGHG3*30 Allele Is Highly Prevalent in Papua New Guinean Children From Sepik
IgG3 polymorphisms of 203 PNG children from the Sepik region were determined (Table 2). Two previously described G3m allotypes were identified; IGHG301 (G3m5) and IGHG314 (G3m21) (Figure 2) [9]. The IGHG330 allele was highly prevalent, with 78% of the children being either homozygous or heterozygous for IGHG330 (Figure 2A).
IgG1 and IgG3 polymorphisms identified in children. Percentage of G3m alleles (A) and G1m alleles (B) in children. C, Highlighted amino acids correspond to the polymorphic positions in the heavy chain constant region of IgG1 and IgG3 allotypes. Abbreviations: Eu, European Union; IMGT, international ImMunoGeneTics information system®.
A PCR-based method was used to identify IgG3 hinge length polymorphisms. Based on amplicon size on agarose gel electrophoresis (Supplementary Figure 1D), children's genotypes were long (LL) (n = 197/203), short (SS) (n = 3/203), and LS heterozygous (n = 3/203); medium hinge length (MM) was not present. Majority of the children carrying an IGHG330 allele (n = 156/159) were homozygous for long hinge. Most of the IGHG314 children had long hinge (n = 100/106) but 3 IGHG314 children were homozygous for small hinge (n = 3/106), and 3 children (IGHG314*/IGHG3*30) were heterozygous “LS” (Table 2).
Two previously described IGHG1 alleles were also identified in the cohort: IGHG101 (G1m17,1) and IGHG108 (G1m3,1) (Figure 2B) [9]. Most children were homozygous for IGHG101 (n = 170), 30 were heterozygous for IGHG101/IGHG108, and only 3 were IGHG108 homozygous (Table 2).
Children Heterozygous for IGHG3*30 Have Fewer P vivax Asymptomatic Infections
To investigate whether IGHG330 is protective, the association between the presence of IGHG330 allele and Plasmodium spp infections was evaluated. IGHG330 allele carriers had on average 1.7 fewer Plasmodium spp infections in the follow-up period compared to non-IGHG330 allele carriers (β = −1.74 [95% confidence interval {CI}, −3.39, −.08]; P = .038). Notably, IGHG330 allele carriers had 1 fewer P vivax asymptomatic infection compared to non-IGHG330 carriers (β = −1.06 [95% CI, −2.01, −.12]; P = .028). The general trend was for less Plasmodium spp infection and morbidity in IGHG330 carriers compared to non-IGHG330 (Table 3).
To investigate the gene dosage effect of IGHG330, children were grouped as IGHG330-homozygous, IGHG330-heterozygous, or non-IGHG330 carriers. Children heterozygous for IGHG330 allele had on average 2.3 fewer Plasmodium spp infections in the follow-up period compared to non-IGHG330 allele carriers (β = −2.34 [95% CI, −4.13, −.54]; P = .011). Most notably, children heterozygous for IGHG330 had 1.4 fewer P vivax asymptomatic infections in the follow-up period compared to non-IGHG330 carriers (β = −1.47 [95% CI, −2.49, −.44]; P = .005). No similar association was seen for children homozygous for the IGHG3*30 allele (Supplementary Table 4).
To investigate the association of IGHG101 with malaria, children were grouped as IGHG101-homozygous and IGHG101-heterozygous. Children heterozygous for IGHG101 had on average 2 fewer Plasmodium spp infections in the follow-up period compared to children homozygous for IGHG1*01 (β = −1.95 [95% CI, −3.88, −.02]; P = .047); however, no significant associations were observed for specific Plasmodium infections (Supplementary Table 5).
Children Heterozygous for Gm Haplotype IGHG101, IGHG330 Have Fewer Plasmodium spp Infections
There was a statistically significant linkage between IGHG330 and IGHG101 (χ^2^ = 8.807, P = .012; Supplementary Table 6). Since IgG alleles are often inherited in combinations as haplotypes, IGHG330 and IGHG101 were grouped together as a haplotype to further investigate any association between Gm haplotypes and Plasmodium spp infections. Children with the Gm IGHG101, IGHG330 heterozygous haplotype had on average almost 3 fewer Plasmodium spp infections (β = −2.95 [95% CI, −5.35, −.55]; P = .016), compared to non-IGHG101, IGHG330 children (grouped together as “other haplotypes”). This effect was most significant for P vivax asymptomatic infections; children with the Gm IGHG101, IGHG330 heterozygous haplotype had on average 1.4 fewer infections (β = −1.4 [95% CI, −2.81, −.05]; P = .042) (Table 4).
By contrast, the number of Plasmodium spp infections in children with the Gm IGHG101, IGHG330 homozygous haplotype was similar to those with the other haplotypes (β = −.15 [95% CI, −1.61, 1.32]; P = .842). The general trend was for less disease in children heterozygous for Gm IGHG101, IGHG330 compared to other haplotypes, and no difference between children homozygous for Gm IGHG101, IGHG330 compared to other haplotypes. The exception to this was that children who were Gm IGHG101, IGHG330 heterozygous (β = −.79 [95% CI, −1.65, .05]; P = .065) or homozygous (β = −.52 [95% CI, −1.04, −.004]; P = .048) had trends toward fewer P falciparum symptomatic infections compared to other haplotypes (Table 4). We assessed the associations between the novel allele and P falciparum force of infection in IGHG3*30 carriers and noncarriers and found no significant association (P = .43).
Children With IGHG3*30 Have Lower Levels of Total IgG to P vivax Proteins
Previous studies investigating IgG allotypes have shown conflicting data in terms of allotype impact on IgG levels and subclass distribution [18, 28]. To understand at a high level any potential differences in IgG acquisition and how this may impact the association with reduced infections in IGHG330 allele carriers, we measured IgG to 6 P vivax recombinant proteins in children with available plasma samples at enrollment (n = 167). Overall, IGHG330 allele carriers had significantly less IgG to 3 of 6 P vivax antigens (Supplementary Figure 2). Notably, children heterozygous for IGHG330 had significantly less IgG to the same 3 proteins, whereas children homozygous for IGHG330 had significantly less IgG only to RBP2b_1 and RBP2b_2 (Figure 3). We grouped children into tertiles (low, medium, high) based on combined antibody levels to the 6 proteins and adjusted the regression model accordingly. We found that IGHG3*30 carriers had significantly fewer P vivax asymptomatic infections after adjusting for IgG levels (β = −.29 [95% CI, −1.20, .62]; P = .035) (Supplementary Table 7).
Total IgG levels to Plasmodium vivax proteins in IGHG330 homozygous, IGHG330 heterozygous, and non-IGHG330 carriers. Data are presented as median and interquartile range. The y-axis shows log-transformed relative antibody unit (RAU) values. P values were determined by Mann-Whitney U test; Abbreviation: ns; nonsignificant.*
DISCUSSION
Here, we confirmed the presence and high prevalence of the novel IgG3 allele, IGHG330, in a cohort of children from East Sepik region of PNG. The high prevalence of IGHG330 allele suggests a protective effect due to possible positive selection in an area of PNG with high malaria transmission. IgG3 variants with H435 (also included in IGHG330) are common in malaria-endemic African countries. Histidine in this position promotes IgG3 binding to FcRn, resulting in a longer half-life and greater transplacental transfer, which has been associated with protection from malaria [12]. However, the effects of IGHG330 on protective effector functions are less clear [18]. Additional studies on IGHG330 allele in other malaria-endemic regions are needed to clarify the potential selective pressures driving IGHG330 to a high prevalence and its role in malaria outcomes.
Similar proportions of children were heterozygous and homozygous for the IGHG330 allele, which is not following Hardy-Weinberg equilibrium (HWE). This could be due to the evolutionary forces like natural selection modulating the allelic distribution in this population [29]. The deviation from HWE in our study population is consistent with our earlier observations in pregnant women [18]. In both cases, we observed an unexpectedly high frequency of IGHG330 homozygotes. This pattern may indicate the action of nonneutral evolutionary forces. In our earlier study, we described functional features of IGHG330, such as stronger binding to FcγRIIa H131 and enhanced ADCP of P falciparum–infected erythrocytes, that could confer a selective advantage [18]. These immune benefits could align with the allele's association with reduced risk of asymptomatic P vivax infections we observed. In addition to positive selection, demographic factors could contribute. IGHG330 is most common in East Sepik, a geographically isolated region with a complex migration history. Gene flow and potential founder effects in this population may explain the observed enrichment of the allele and the deviation from HWE.
Importantly, children with the IGHG330 allele had fewer Plasmodium spp infections, particularly P vivax asymptomatic infections, as compared to children without IGHG330. This effect was particularly evident for children heterozygous for IGHG3*30. Asymptomatic infections not only serve as a reservoir for continued malaria transmission and pose challenges for malaria elimination, but they can also progress to severe disease [30]. The strong association with P vivax asymptomatic infections may reflect their high incidence in this cohort. Children in this cohort acquired immunity to P vivax infections more quickly than to P falciparum [20].
It is not clear why IGHG330 is more protective in heterozygous form, but several factors might explain this observation. First, in our earlier study, IGHG330 homozygotes had lower total and antigen-specific IgG3 levels but showed enhanced ADCP via increased FcγRIIa binding. It is possible that protection peaks in heterozygotes due to a balance between expression levels and effector function, while homozygotes might experience diminishing returns or altered subclass switching. Second, some form of balancing selection could be occurring. Co-inheritance of IGHG330 with IGHG101, for example, may contribute to immune modulation that differs between heterozygotes and homozygotes. Last, the difference in cohorts may also play a role. The current study focused on young children, whereas our previous work involved pregnant women. Age, immune status, and exposure history could influence the functional impact of this allele. Moreover, the distinct B-cell populations in heterozygotes undergo allelic exclusion and are committed to expressing a single allele. Thus, distinct B cells produce IgG3 with His433 and Glu433, which may result in an additive effect.
Several studies have examined the associations between IgG allotypes and malaria immunity [28, 31, 32]. The distribution of IgG allotypes differs among populations and has been associated with malaria resistance or susceptibility [28]. A study reported that Beninese children with IGHG301 had significantly higher ADCP of merozoites compared to those with other IgG3 allotypes [32]. Certain IgG allotypes mediate increased opsonic phagocytosis, reflecting their increased affinity to one or the other FcγR [33, 34]. Previously, monoclonal antibodies with the IGHG330 sequence showed high affinity to FcγRIIa [18], which is present on classical monocytes and neutrophils, and women with the IGHG330 allele showed greater antibody-dependent phagocytosis of infected erythrocytes. Given the importance of FcγRIIa in antibody-dependent phagocytosis [35, 36], we postulate that enhanced ADCP could contribute to the observed protective association. FcγRIIa-mediated phagocytosis by monocytes and neutrophils has been linked to protection in several malaria studies, and the increased ADCP we observed in IGHG330 carriers aligns well with this mechanism. However, other immune pathways are involved in protection from blood-stage infection. For example, FcγRIIIa-mediated ADCC or complement activation, both of which are less efficient with IGHG330, may play a more prominent role in protection from severe or symptomatic disease. Our current data suggest that IGHG330 may enhance one pathway, ADCP, while having a more limited effect on others.
A long hinge region increases IgG3 flexibility and increases opsonic phagocytosis [37]. In Ghana, a medium hinge length was associated with greater odds of cerebral malaria compared to a long hinge length [26]. All the children homozygous for the IGHG330 allele (n = 77) in our study had a long IgG3 hinge region (Supplementary Table 2). Notably, we also found that most children with IGHG314 (n = 32/35) had a long IgG3 hinge, while 3 had a small hinge and 3 were heterozygous (LS) (Table 2). To our knowledge this is the first study to report this novel hinge length polymorphism in IGHG314. Previously, 2 IgG3 variants IGHG301 (long hinge) and IGHG304 (small hinge) under G3m5 mosaic were shown to have the same sequence and to vary in hinge length [16].
IgG allotypes could also affect susceptibility to malaria and disease outcomes by influencing the subclass distribution and affinity of antimalarial antibodies [38–41]. Damelang et al reported elevated proportions of IgG1 (measured as % of total IgG) but lower proportions of IgG3 in plasma of women with IGHG330 allele as compared to women with IGHG301 allele [18]. Caucasian individuals with the Gm 3,23;5,13 phenotype showed lower concentrations of IgG1 but higher concentrations of IgG3 in normal human serum [40]. In the present study, levels of IgG to P vivax proteins, previously defined as serological markers of exposure [27], were assessed. IgG levels were significantly lower in children heterozygous for IGHG330 compared to children without IGHG330. Further research is needed to understand the impact of the IGHG330 allele on the acquired immune response to infection, and to assess whether antibodies isolated from these children have different abilities to induce effector functions. As acquisition and boosting of antibodies is impacted by exposure to Plasmodium parasites, and levels of antibodies can change over time, further work is needed to understand the potentially complex interplay of these factors and the IGHG330 itself.
Our study corroborates the discovery of IGHG330 in PNG [18] and extends the finding beyond pregnant women. In this longitudinal study we established relationships between IGHG330 allele and Plasmodium spp infections. A longitudinal study has an advantage over a cross-sectional design such as getting information at multiple time-points, but limitations of our study include the sample size and unequal distribution of Gm allotypes between groups, which affected statistical power. Future studies could investigate antigen-specific subclasses and antibody half-life, to better characterize IGHG330. With few severe malaria cases, associations of IGHG330 allele with severe malaria could not be made. Further studies of IgG3 alleles in other malaria-endemic regions and globally could help elucidate if IGHG330 is found more broadly and should examine potential associations between IGHG330 and severe malaria. Including control antigens like tetanus or influenza could help to show if IgG levels for those antigens follow a different trend as compared to Plasmodium antigens.
In conclusion, our data provide compelling evidence that the IGHG330 allele confers protection against Plasmodium spp infections and is probably under positive selection. Its high prevalence in children from the East Sepik region of PNG underscores its potential functional significance. The strong associations between IGHG330 and lower antibody levels against P vivax antigens further suggest that malaria has been a key evolutionary driver in maintaining this polymorphism within the population.
Supplementary Material
jiaf390_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1World Health Organization . World malaria report 2024. 2024. https://www.who.int/teams/global-malaria-programme. Accessed 7 August 2025.
- 2Carter R, Mendis KN. Evolutionary and historical aspects of the burden of malaria. Clin Microbiol Rev 2002; 15:564–94.12364370 10.1128/CMR.15.4.564-594.2002 PMC 126857 · doi ↗ · pubmed ↗
- 3Kariuki SN, Williams TN. Human genetics and malaria resistance. Hum Genet 2020; 139:801–11.32130487 10.1007/s 00439-020-02142-6PMC 7271956 · doi ↗ · pubmed ↗
- 4Cohen S, Mc Gregor IA, Carrington S. Gamma-globulin and acquired immunity to human malaria. Nature 1961; 192:733–7.13880318 10.1038/192733 a 0 · doi ↗ · pubmed ↗
- 5Weaver R, Reiling L, Feng G, et al The association between naturally acquired Ig G subclass specific antibodies to the Pf RH 5 invasion complex and protection from Plasmodium falciparum malaria. Sci Rep 2016; 6:33094.27604417 10.1038/srep 33094 PMC 5015043 · doi ↗ · pubmed ↗
- 6Aitken EH, Damelang T, Ortega AP, et al Developing a multivariate prediction model of antibody features associated with protection of malaria-infected pregnant women from placental malaria. Elife 2021; 10:e 65776.34181872 10.7554/e Life.65776 PMC 8241440 · doi ↗ · pubmed ↗
- 7Opi DH, Kurtovic L, Chan JA, Horton JL, Feng G, Beeson JG. Multi-functional antibody profiling for malaria vaccine development and evaluation. Expert Rev Vaccines 2021; 20:1257–72.34530671 10.1080/14760584.2021.1981864 · doi ↗ · pubmed ↗
- 8Damelang T, Brinkhaus M, van Osch TL, et al Impact of structural modifications of Ig G antibodies on effector functions. Front Immunol 2024; 14:1304365.38259472 10.3389/fimmu.2023.1304365 PMC 10800522 · doi ↗ · pubmed ↗