Impact of Helicobacter pylori Virulence Genotypes cagA, vacA, oipA, and babA2 on Severity of Gastropathies in Brazilian Patients
Diogo Nery Maciel, Hellen Christina de Oliveira Santos-Dutra, Viviane Lopes Rocha, Lucas Trevizani Rasmussen, Mônica Santiago Barbosa

TL;DR
This study examines how specific Helicobacter pylori gene combinations affect the severity of stomach diseases in Brazilian patients.
Contribution
The study identifies a protective effect of combined virulence genes in H. pylori against certain gastropathies.
Findings
The cagA/vacA/oipA/babA2 gene combination was protective against erosive esophagitis and duodenitis.
No significant link was found between the gene combination and severe gastric diseases.
A trend toward protection against gastric atrophy was observed.
Abstract
Helicobacter pylori (H. pylori) is a Gram-negative, spiral-shaped bacterium that colonizes the human stomach and is linked to various gastroduodenal diseases. The severity of different clinical outcomes may be determined by the combination of virulence genes. The aim of this study was to assess the combinations of the cytotoxin-associated gene A (cagA), the vacuolating cytotoxin A gene (vacA), the outer inflammatory protein A gene (oipA), and the blood group antigen-binding adhesin gene (babA2) genotypes in H. pylori and their associations with the clinical outcomes of infection in patients from Central Brazil. This cross-sectional study included 106 patients who underwent endoscopy or gastrectomy. The presence and genotypes of H. pylori were confirmed using Polymerase Chain Reaction (PCR). Gastropathies were classified according to established severity criteria. Multivariate logistic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
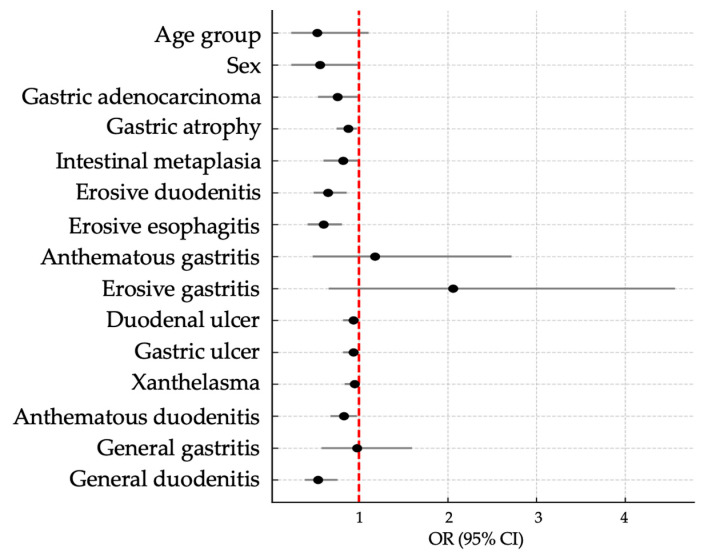 Figure 1
Figure 1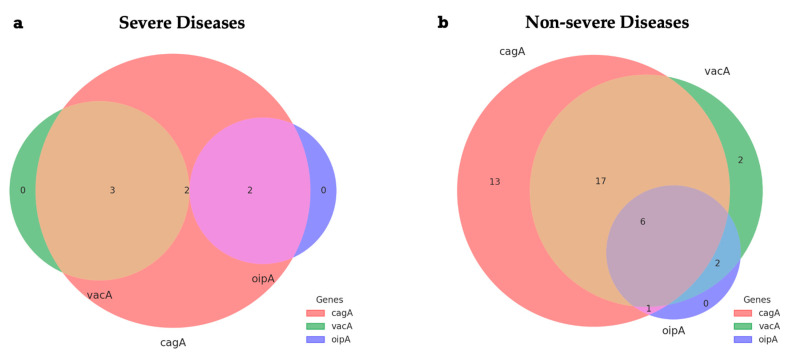 Figure 2
Figure 2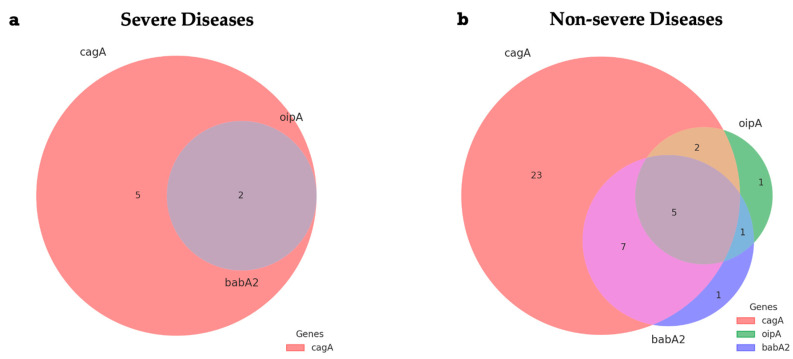 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHelicobacter pylori-related gastroenterology studies · Galectins and Cancer Biology · Eosinophilic Esophagitis
1. Introduction
Helicobacter pylori (H. pylori) harbors a range of virulence genes that are associated with various gastric pathologies, including gastritis, peptic ulcers, Mucosa-Associated Lymphoid Tissue (MALT) lymphoma, and gastric cancer [1,2]. It is estimated that more than 50% of the global population is infected. However, prevalence rates vary substantially, as factors such as lifestyle and socioeconomic status directly influence the infection rate, which varies across different settings, being significantly higher in developing countries (50.8%), such as Brazil, compared to developed countries (34.7%) [1,3,4].
The infection often persists asymptomatically for long periods, which favors the progression of these gastropathies. It is a Gram-negative, microaerophilic, flagellated bacterium, whose adaptive capacity helps in the colonization of the human gastrointestinal tract of approximately half of the global population [5,6,7]. This successful adaptation of the bacterium to the hostile environment of the stomach is favored by a series of virulence factors, which allow its adhesion to the gastric epithelium and evasion of the host’s immune system [8,9].
Prevalence studies conducted in latin american populations have found rates between 60% and 91% [10,11,12]. The choice of the central-west region of Brazil for this study was due to the geographic diversity of the pathogen and the scarcity of data related to the molecular epidemiology of H. pylori in this area. This region is characterized by significant socioeconomic contrasts, the coexistence of urban and rural areas with different levels of infrastructure and access to health care, and weaknesses in epidemiological surveillance. It is believed that these factors collectively influence both the prevalence of infection and the genetic variability of circulating strains, highlighting the importance of conducting specific regional studies.
Among the most studied virulence factors of H. pylori are the cagA, vacA, oipA, and babA genes, which play fundamental roles in the pathogenesis of the infection. The CagA (cytotoxin-associated gene A) protein is one of the main proteins responsible for inducing inflammatory responses and altering cellular signaling in the host. Its activity, particularly in association with the type IV secretion system, is strongly implicated in the genesis of epithelial lesions and gastric carcinogenesis [13]. VacA (vacuolating cytotoxin A) plays an important role in modulating the immune response and vacuolation of gastric cells, inducing cellular damage and stimulating an environment conducive to pathogen persistence [14].
The OipA (outer Inflammatory Protein A) plays a key role in modulating gastric inflammation by inducing the expression of pro-inflammatory cytokines, like interleukin-8 (IL-8), and enhancing bacterial adhesion to the stomach mucosa, which exacerbates the inflammatory response [15]. OipA expression triggers apoptotic pathways and cytoskeletal changes in gastric epithelial cells, contributing to the progression of severe gastropathies [16]. On the other hand, the BabA protein (blood-group antigen-binding adhesin) acts as a specialized adhesin, mediating the attachment of H. pylori to glycosylated structures expressed on the gastric mucosa. This interaction is a critical step in establishing and maintaining the host-pathogen relationship [17].
H. pylori’s extensive genetic plasticity influences the clinical variability observed in infected individuals. Co-expression of certain genes can amplify inflammation, promote infection persistence, and increase the risk of severe gastric complications [18]. Studies indicate that the association between certain alleles of cytotoxin-associated gene A (cagA) and vacuolating cytotoxin A gene (vacA), for example, is directly related to the progression to gastric cancer, while the concomitant expression of outer inflammatory protein A gene (oipA), and the blood group antigen-binding adhesin gene (babA2) may confer a greater capacity for adhesion and colonization of the gastric epithelium [19,20].
Recent research indicates that the expression of OipA is associated with other H. pylori virulence markers, such as cagA, vacA s1/m1 and babA2. In strains with preserved cag-PAI pathogenicity island, there is greater coexpression of oipA, which suggests a possible synergistic effect between these genetic determinants. The geographic variability of these virulence factors may represent an adaptive mechanism to the gastric microenvironment, which interacts with host elements and environmental factors, and influences the clinical manifestations of the infection [2,21].
Therefore, the combination of multiple virulence factors plays a critical role in the progression of infection and the severity of associated gastric lesions. Integrated analysis of these genetic profiles is essential for understanding the pathogenesis of H. pylori [15].
The advent of multi-omic approaches, integrating genomics, transcriptomics, proteomics, and metabolomics, has decisively expanded the understanding of the Helicobacter pylori–host relationship. The simultaneous analysis of multiple molecular contexts can yield consistent associations between virulence factors and clinical outcomes [22,23,24]. Within this framework, the present study employs a combinatorial gene analysis approach, aligning with contemporary trends in systems biology to provide an integrated perspective on H. pylori virulence.
This study aimed to investigate the combinations between the cagA, vacA, oipA, and babA2 genotypes of H. pylori and their associations with different clinical outcomes in patients from Central Brazil.
2. Results
2.1. Demographic and Clinical Characteristics of Participants
A total of 106 patients with dyspepsia, who underwent upper gastrointestinal endoscopy or gastrectomy as indicated for diagnostic purposes, were included in the study. Of these, 69 tested positive for H. pylori, representing an approximate prevalence of 65.1% in this sample. The population consisted of 82 females and 24 males, aged between 18 and 75 years, with a predominance of individuals over 51 years of age. Infection was more common among participants with primary education and those from families with a monthly income between one and two minimum wages (Table 1).
2.2. Endoscopic Findings and Their Association with H. pylori Infection
Table 2 presents the distribution of clinical–endoscopic findings in the sample of 106 patients with dyspepsia, classified according to the presence or absence of H. pylori. Among the infected patients (n = 69), the most common findings were anthematous gastritis (42.0%) and erosive gastritis (30.4%). Less prevalent findings included gastric adenocarcinoma (2.9%), intestinal metaplasia (2.9%), and gastric ulcer (1.4%). In the non-infected group (n = 37), anthematous gastritis was also the most common finding (37.8%), followed by erosive gastritis (29.7%) and gastric atrophy (18.9%). A significant difference was observed for the diagnosis of gastric atrophy (p = 0.035), which was more prevalent among non-infected patients. Other diagnoses, such as intestinal metaplasia, erosive duodenitis, erosive esophagitis, and general gastritis, did not show statistically significant differences between the groups (p > 0.05). The overall frequency of severe gastric disease was relatively low in both groups, with a predominance of non-severe lesions in the total sample.
The distribution of H. pylori virulence gene combinations according to endoscopic diagnosis is described in Table 3.
2.3. Association Between Virulence Genes and Demographic or Clinical Variables
The cagA/babA2 combination had the highest overall frequency (21.7%), followed by cagA/oipA (13.0%) and cagA/oipA/babA2 (10.1%). No gene combination showed a statistically significant association with the severity of gastric disease (p > 0.05). The analysis of H. pylori virulence gene combinations is essential for understanding the impact of these genetic variants on the pathogenesis of gastric diseases. However, the lack of significant association with clinical outcomes observed in this study may reflect the complexity of interactions among microbial virulence factors, host genetics, and other factors that influence disease progression.
The cagA/vacA/oipA/babA2 gene combination did not show significant differences regarding age group (p = 0.120) or sex (p = 0.100), despite the predominance of this gene combination in the female group (80%). The association with gastric adenocarcinoma and intestinal metaplasia was not statistically significant, but a protective trend against gastric atrophy was observed (p = 0.049; Odds Ratio (OR) = 0.88; 95% Confidence Interval (CI): 0.75–0.98). Among non-severe diseases, the presence of this gene combination was correlated with a lower risk of erosive duodenitis (p = 0.003; OR = 0.65; 95% CI: 0.49–0.86) and erosive esophagitis (p = 0.002; OR = 0.60; 95% CI: 0.42–0.81). Additionally, individuals with the cagA/vacA/oipA/babA2 combination had a lower likelihood of developing general duodenitis (p < 0.001; OR = 0.54; 95% CI: 0.39–0.76). These findings suggest a potential protective role for this gene combination against inflammatory lesions in the gastrointestinal tract (Table 4).
2.4. Distribution Patterns of Virulence Gene Combinations
A more detailed analysis of the cagA/vacA/oipA/babA2 gene combination and predictive variables, such as sociodemographic characteristics, risk factors, and the severity of gastropathies, is presented in the Forest Plot shown in Figure 1, which displays the Odds Ratio values.
The Venn diagrams represented in Figure 2 show the distribution of combinations of the H. pylori virulence genes cagA, vacA, and oipA in patients diagnosed with severe or non-severe gastric disease. The intersections indicate the coexistence of two or more genes in the same sample. In the group with severe disease, seven samples exhibited the cagA gene, of which three had vacA coexisting, two had oipA coexisting, and two had cagA, vacA, and oipA simultaneously. The isolated presence of vacA or oipA was not observed in this group.
Figure 3 shows two Venn diagrams that illustrate the distribution of combinations of the H. pylori virulence genes cagA, oipA, and babA2 in patients diagnosed with severe or non-severe gastric disease. The intersections indicate the coexistence of two or more genes in the same sample. Among the cases classified as severe, five samples presented only the cagA gene, while two samples simultaneously exhibited cagA, oipA, and babA2. No cases of coexistence between cagA and babA2, or isolated manifestation of oipA or babA2, were identified in this group. In the group with non-severe disease, the predominance of samples positive only for cagA was observed (n = 23). In addition, seven samples presented coexistence of cagA and babA2, five simultaneously exhibited the three genes (cagA, oipA, and babA2), and two cases were positive for cagA and oipA. Three unique observations were also detected: one for oipA, one for babA2, and a combination of oipA and babA2 without cagA.
3. Discussion
This study evaluated the combinations of the cagA, vacA, oipA, and babA2 virulence genes and their associations with the severity of gastrointestinal lesions in a dyspeptic population infected with H. pylori in the Central region of Brazil. The prevalence of infection was 65.1. The distribution of the studied genes and sociodemographic variables, such as age and sex, did not show significant differences. These findings support observations that the pathogenicity of H. pylori seems to be more related to bacterial and immunological factors than to isolated sociodemographic factors [6,25].
The frequency of the cagA (79.7%) and vacA (53.6%) genes was higher in this study, followed by babA2 (15.9%) and oipA (26.1%). Although cagA was more common among cases of severe disease, its presence did not reach statistical significance, similar to the other genes. Previous studies suggest that the isolated presence of these genes does not absolutely predict the severity of clinical manifestations, but certain combinations can enhance the microorganism’s infection capability [26,27].
In this study, the cagA/vacA/oipA/babA2 combination showed a significant protective association against inflammatory lesions, such as erosive duodenitis, erosive esophagitis, and general duodenitis. These findings support the idea that the simultaneous interaction of multiple virulence factors can modulate the host’s inflammatory response, reducing tissue damage [28]. Despite the low frequency of subgroups expressing the cagA/vacA/oipA/babA2 gene combination, the complete absence of certain inflammatory lesions among these individuals suggests a potential factor associated with clinical protection. Although numerically limited, the asymmetric distribution led to statistically significant associations, raising the hypothesis of a possible biological effect.
The combined analysis of the cagA, vacA, oipA, and babA2 genes in this study was based on both their functional roles, described in the literature, and the frequency with which these genes are investigated as key H. pylori virulence markers. While some studies have not found a direct association between these genes and specific clinical outcomes [29,30], others have suggested trends within subgroups, such as the significant association observed for the cagA+/vacA s1/m1 genotype [31]. Furthermore, Barhoine et al. (2025) [2] reported that although co-occurrence among several virulence genes was statistically significant in their study, oipA, babA, and iceA showed no direct correlation with histopathological parameters, reinforcing the complexity of gene–gene interactions and the importance of contextual analysis.
This complex interaction between pathogen and host mechanisms, reinforced by the combination of virulence genes, allows for successful bacterial colonization through evasion of the immune system [32].
Our results suggest that the combined presence of the cagA, vacA, and oipA genes is more common in non-severe forms of the disease, although the cagA gene is present in all cases classified as severe. This contrasts with a study conducted in the Southeast region of Brazil, where the combination of the cagA, vacA, dupA, and oipA virulence factors showed high prevalence in patients with chronic gastritis and gastric cancer [31].
The absence of vacA and oipA as isolated factors in severe diseases reinforces the possibility that increased pathogenicity is more associated with the presence of cagA or the combination of multiple genes. In contrast, non-severe lesions demonstrated greater diversity of gene combinations, while severe disease cases showed a more restricted profile, concentrated on the isolated presence of cagA or the triple gene association. In a study conducted in Ecuador, the cagA/oipA combination was associated with an increased risk of developing acute gastric inflammation and lymphoid follicular hyperplasia [24].
The findings of this research reinforce the central role of the cagA gene in the virulence of the microorganism, especially in more aggressive clinical forms. The cag pathogenicity island (cag-PAI) contains genes encoding a secreted effector protein (CagA) and components of a type IV secretion system (Cag T4SS), which are closely related to the pathogenesis of H. pylori [33]. Approximately 60% to 70% of H. pylori strains express CagA, leading to increased synthesis of interleukin-8 (IL-8). The presence of tyrosine-containing motifs in the CagA structure, once phosphorylated, allows for its translocation into gastric epithelial cells via the Cag T4SS. Alterations in the sequence of these motifs have been correlated with gastric mucosal degeneration and increased predisposition to gastric adenocarcinoma [34].
The oipA and babA2 genes, important for bacterial adhesion to the gastric mucosa, showed prevalences of 15.9% and 26.1%, respectively. Despite their recognized role in bacterial adhesion and inflammation induction, their isolated combinations were not associated with lesion severity, suggesting that their influence depends on the genetic context of the strain and host factors. The lower frequency of the oipA genotype observed in this study contrasts with data from populations in Vietnam, where high prevalence and association with pre-neoplastic gastric mucosal alterations have been reported [12]. Conversely, despite the high detection of oipA in studies conducted in China, no significant correlation was found between this genotype and gastroduodenal clinical manifestations [14]. These data may reflect unique characteristics of the studied populations.
Among the main limitations of this study are the exploratory design, including the reduced sample size of specific subgroups, especially the subgroup with severe clinical manifestations, in addition to the absence of functional analyses and the lack of control for host variables. However, the findings reinforce the importance of the molecular characterization of H. pylori strains and serve as a basis for future hypotheses that encourage the development of broader genomic and immunogenetic approaches, with more integrative investigative strategies. Its contribution lies in the identification of preliminary patterns that should be validated by future studies with greater sample representation and statistical power.
4. Materials and Methods
4.1. Population and Samples
This cross-sectional study included 106 individuals with dyspeptic symptoms, aged 18 to 75, who underwent upper gastrointestinal endoscopy or gastrectomy at referral hospitals in the city of Goiânia, Goiás, Brazil. The exclusion criteria ruled out patients currently using proton pump inhibitors, H2 receptor antagonists, immunosuppressive agents, or antibiotics. Additionally, participants with evidence of active gastrointestinal bleeding, pregnant or lactating women, and those deemed ineligible for endoscopy were excluded. All participants signed an informed consent form (ICF) and completed a sociodemographic questionnaire. The confidentiality and privacy of all personal data were strictly protected throughout the study, ensuring the integrity of participant information.
Sample collection followed the guidelines of the IV Brazilian Consensus on Helicobacter pylori Infection. During the endoscopic procedure, two biopsy samples were taken from the antral region and two from the gastric body, as recommended for histological representativeness [35]. Tumor samples from patients with gastric cancer were obtained during gastrectomy, ensuring the preservation of morphological integrity and neoplastic tissue representativeness for analysis [36].
One sample from the antrum and another from the gastric body were sent to the Clinical Pathology Laboratory of the University Hospital at the Federal University of Goiás (HC/UFG) for histopathological analysis. The two remaining fragments were refrigerated and stored at −20 °C in the Helicobacter pylori Research Center (NEHP) of the same institution. These samples were later subjected to molecular analysis following previously established protocols, as described in prior publications by the NEHP [37,38].
4.2. Histology
Gastric mucosa samples obtained through endoscopy or gastrectomy were fixed in 10% buffered formalin solution. The tissue fragments were then stained with hematoxylin-eosin and Giemsa. Histopathological evaluation was performed according to the Sydney System, which standardizes the classification of morphological changes in the gastric mucosa [39].
4.3. DNA Extraction
DNA extraction followed the manufacturer’s protocol for the QIAamp DNA Mini Kit^®^ (Qiagen, Valencia, CA, USA). The quantification and assessment of the purity of the extracted genomic DNA were conducted using optical density measurements with a NanoDrop^®^ ND-1000 UV-Vis spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA), ensuring the integrity and quality of the genetic material for subsequent analyses.
4.4. DNA Amplification of H. pylori and Virulence Genes cagA, vacA, oipA, and babA2
The PCR technique was used for the molecular diagnosis of H. pylori infection, targeting the amplification of a fragment of the 16S ribosomal RNA (rRNA) gene. Samples that tested positive for H. pylori were then further analyzed for the detection of the virulence genes cagA, vacA, oipA, and babA2.
The amplification of the 16S rRNA, cagA, vacA, oipA, and babA2 genes was carried out using a BioRad^®^ thermal cycler (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Each PCR reaction contained the following components: 0.5 µL of Taq DNA polymerase (2.5 units), 5 µL of CoralLoad 10X PCR buffer (QIAamp, Qiagen) containing 1.5 mM MgCl_2_, 2 µL of dNTPs (2.5 mM), and 33.5 µL of milli-Q water. After mixing, 5 µL of DNA from each sample (150 ng) was added, bringing the total reaction volume to 50 µL. DNA from H. pylori-positive samples was used as a positive control in the PCR reactions. DNA-free water was used as a negative control to ensure the absence of contamination and validate the specificity of the amplification process.
Details regarding the oligonucleotides used for the amplification of each gene, along with the PCR conditions and expected fragment sizes, are provided in Table 5 [40,41,42,43].
The PCR products were stained with BlueGreen nucleic acid dye (Lab Biotechnology^®^ Co., Ltd., Beijing, China) and subjected to electrophoresis on a 1.5% agarose gel. The visualization of the fragments was performed under ultraviolet (UV) light using a transilluminator, confirming the presence and size of the generated amplicons.
4.5. Diagnosis of Gastroduodenal Diseases and Severity Criteria
The stratification of patients with gastroduodenal lesions, based on the degree of severity, was conducted by the gastroenterology services and clinical pathology laboratories of the recruiting hospitals. This classification relied on the analysis of endoscopic reports and histopathological examinations. Clinical outcomes were grouped into two categories: non-severe, including duodenitis, esophagitis, gastritis, xanthelasma, and ulcers; and severe, comprising gastric adenocarcinoma, gastric atrophy, and intestinal metaplasia [44].
4.6. Data Analysis
Descriptive statistics were used to characterize the demographic and clinical profiles of the patients. Categorical variables were expressed as absolute frequencies (n) and relative frequencies (%), while continuous variables were reported as means and standard deviations. To compare the distribution of patient characteristics according to H. pylori infection status (positive vs. negative), Pearson’s chi-square test was applied.
To investigate the associations of individual virulence genes (cagA, vacA, dupA, oipA, babA2) and their combinations (e.g., cagA/vacA/oipA/babA2, cagA/oipA, cagA/babA2, cagA/oipA/babA2) with clinical outcomes, multiple logistic regression models were fitted. Each model included one gene or gene combination as the binary outcome variable (presence/absence), and the clinical and demographic features were included as independent predictors. The predictor variables entered into the models were as follows: age group, sex, and the presence or absence of each of the following clinical diagnoses—gastric adenocarcinoma, gastric atrophy, intestinal metaplasia, erosive duodenitis, erosive esophagitis, enanthematous gastritis, erosive gastritis, duodenal ulcer, gastric ulcer, xanthelasma, enanthematous duodenitis, general gastritis, and general duodenitis.
To prevent multicollinearity and overfitting, all regression models were adjusted using ridge penalization. Odds Ratios, 95% Confidence Intervals, Wald statistics, and p-values were reported for each predictor variable.
Additionally, the distribution of virulence gene combinations was analyzed using binary coding (presence = 1, absence = 0), and patients were categorized into two clinical outcome groups, “Severe” and “Non-severe” gastropathy. Venn diagrams were used to graphically illustrate the intersections among gene combinations in both groups. These diagrams were created in R (version 12.1) using the VennDiagram and ggVennDiagram packages, applying the venn.diagram() function for clinical group stratification.
All statistical analyses were conducted using SPSS version 26.0 (IBM Corp., Armonk, NY, USA), with a two-tailed significance level set at α = 0.05.
5. Conclusions
In this study, the cagA/vacA/oipA/babA2 combination demonstrated a potential protective effect against the occurrence of erosive esophagitis, erosive duodenitis, and unspecified duodenitis. These findings suggest that the synergistic interaction of multiple bacterial virulence determinants may modulate the host’s inflammatory response, directly influencing the clinical outcome of H. pylori infection. In exploratory studies involving low-prevalence genetic variables, such evidence may support initial hypotheses when interpreted with caution. In this context, the results, despite statistical limitations, may contribute to the understanding of the interactions between virulence genotypes and clinical manifestations of H. pylori infection. Future investigations should aim to further characterize virulence gene variants and explore the immunogenetic profiles of hosts, with the goal of elucidating the molecular mechanisms involved in pathogenesis and refining risk stratification for the development of more severe clinical complications.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wattanawongdon W. Simawaranon Bartpho T. Tongtawee T. Relationship between Helicobacter pylori virulence genes and gastroduodenal disease J. Int. Med. Res.2023510300060523116146510.1177/0300060523116146536924226 PMC 10026102 · doi ↗ · pubmed ↗
- 2Barhoine M. Moustaoui F. Hammani O. Aghrouch M. Lemkhente Z. Belhabib Z. Bajaddoub Z. Touyar A. Aqoudad N. Rherissi B. The Effect of Helicobacter pylori Gene Combinations of cag A, cag E, vir B 11, vac A and bab A on the Outcome of Gastric Disease in a Southern Moroccan Population Pathogens 20251427910.3390/pathogens 1403027940137764 PMC 11944658 · doi ↗ · pubmed ↗
- 3Bashir S.K. Khan M.B. Overview of Helicobacter pylori Infection, Prevalence, Risk Factors, and Its Prevention Adv. Gut Microbiome Res.20232023974702710.1155/2023/9747027 · doi ↗
- 4Zamani M. Ebrahimtabar F. Zamani V. Miller W.H. Alizadeh-Navaei R. Shokri-Shirvani J. Derakhshan M.H. Systematic review with meta-analysis: The worldwide prevalence of Helicobacter pylori infection Aliment. Pharmacol. Ther.2018186887610.1111/apt.1456129430669 · doi ↗ · pubmed ↗
- 5Sedarat Z. Taylor-Robinson A.W. Helicobacter pylori Outer Membrane Proteins and Virulence Factors: Potential Targets for Novel Therapies and Vaccines Pathogens 20241339210.3390/pathogens 1305039238787244 PMC 11124246 · doi ↗ · pubmed ↗
- 6Hooi J.K. Ying Lai W. Khoon Ng W. Suen M.M. Underwood F.E. Tanyingoh D. Malfertheiner P. Graham D.Y. Wong V.W. Wu J.C. Global Prevalence of Helicobacter pylori Infection: Systematic Review and Meta-analysis Gastroenterology 201715342042910.1053/j.gastro.2017.04.02228456631 · doi ↗ · pubmed ↗
- 7Wondmagegn Y.M. Girmay G. Amare G.A. Assefa M. Tamir M. Abriham Z.Y. Setegn A. Prevalence of intestinal parasites and Helicobacter pylori co-infection in people with gastrointestinal symptoms in Africa: A systematic review and meta-analysis BMC Infect. Dis.2025252010.1186/s 12879-024-10432-x 39754070 PMC 11699658 · doi ↗ · pubmed ↗
- 8Teng K.W. Hsieh K.S. Hung J.S. Wang C.J. Liao E.C. Chen P.C. Lin Y.H. Wu D.C. Lin C.H. Wang W.C. Helicobacter pylori employs a general protein glycosylation system for the modification of outer membrane adhesins Gut Microbes 202214213065010.1080/19490976.2022.213065036206406 PMC 9553153 · doi ↗ · pubmed ↗