Transcriptome Analysis of Potential Genes Involved in Innate Immunity in Mudflat Crab (Helice tientsinensis)
Lulu Chen, Ming Wang, Mengdi Zhou, Youkun Fang, Tingting Ji, Ruyang Xia, Menglu Bai, Zhengfei Wang, Jiafei Shen

TL;DR
This study explores how mudflat crabs respond to Vibrio infection by analyzing gene expression changes in their gills and hepatopancreas.
Contribution
The study identifies differentially expressed genes and immune pathways in mudflat crabs during Vibrio infection, offering new insights into crustacean immunity.
Findings
Vibrio parahaemolyticus infection causes structural damage in gill and hepatopancreatic tissues of H. tientsinensis.
Transcriptome analysis identified thousands of differentially expressed genes linked to ribosomal subunits and energy transport.
Key immune pathways and amino acid metabolism are involved in the crab's defense against Vibrio infection.
Abstract
Vibrio species are widely recognized as highly pathogenic bacteria in aquaculture, causing extensive mortality among cultured organisms and resulting in substantial economic losses. H. tientsinensis, a dominant crustacean species in the Yellow River Delta, has been the subject of limited investigation regarding its susceptibility to Vibrio infection. In this study, we examined structural alterations and differential gene expression in the gill and hepatopancreatic tissues of H. tientsinensis following infection with Vibrio parahaemolyticus, and further explored the associated metabolic and immune responses. The findings provide valuable insights into the defense mechanisms of crustaceans against Vibrio pathogens and offer a scientific foundation for aquaculture management and targeted disease prevention strategies. The mudflat crab (H. tientsinensis) is a dominant species in coastal…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
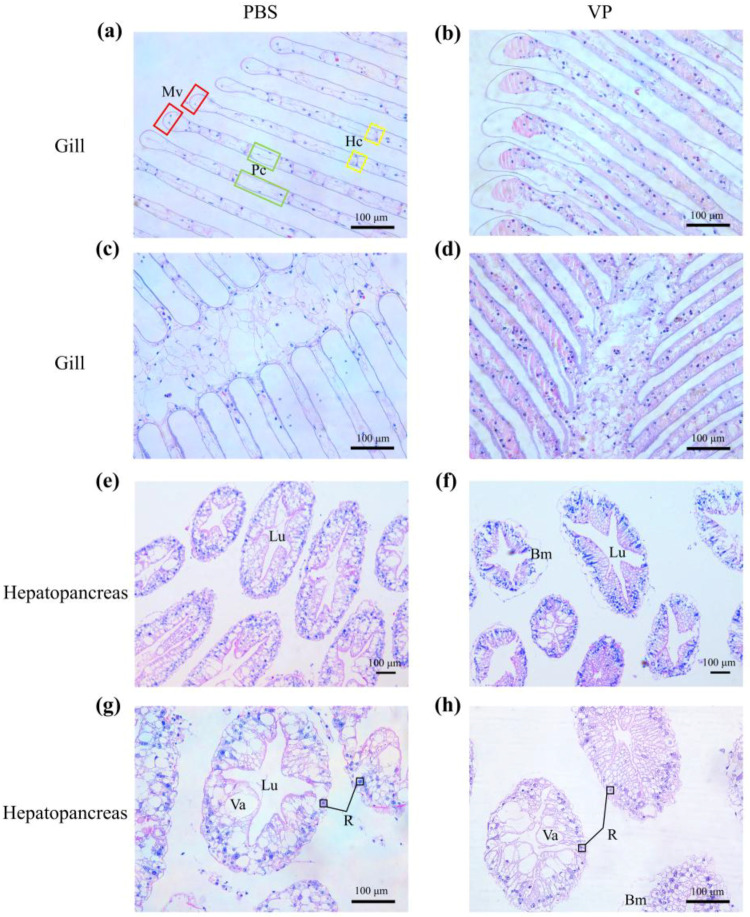 Figure 1
Figure 1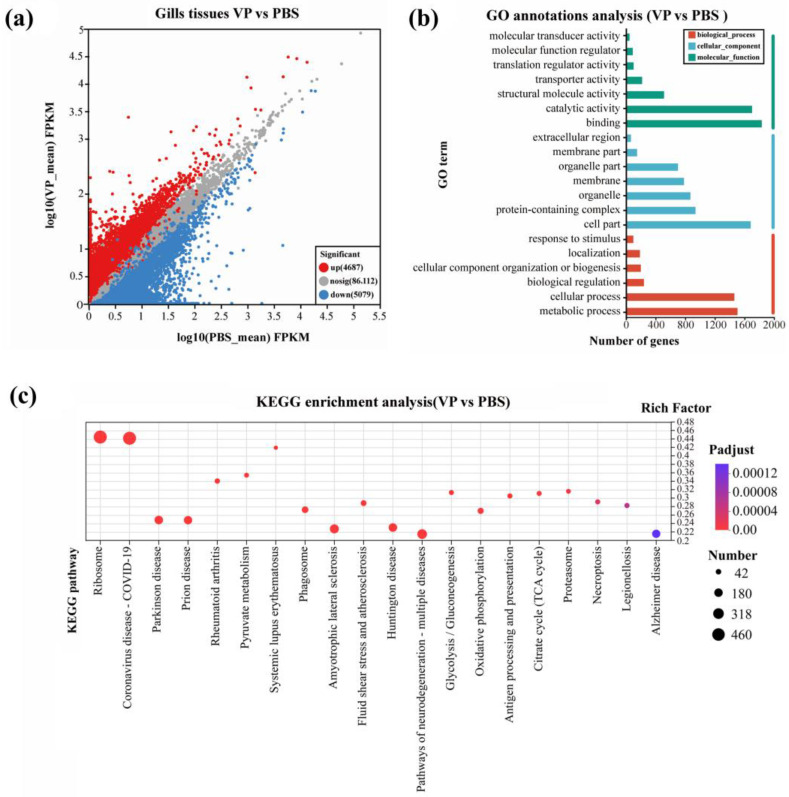 Figure 2
Figure 2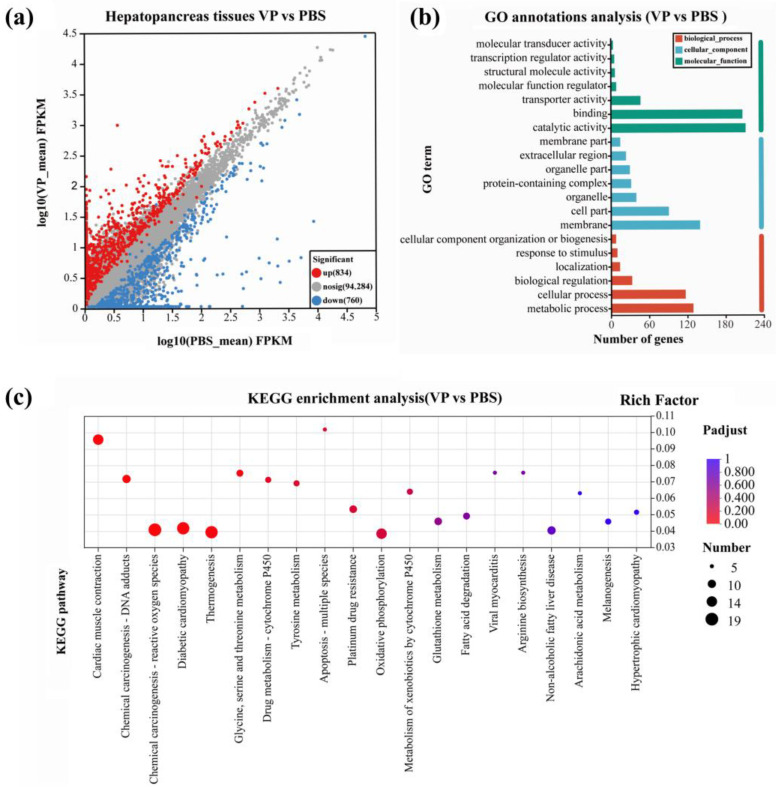 Figure 3
Figure 3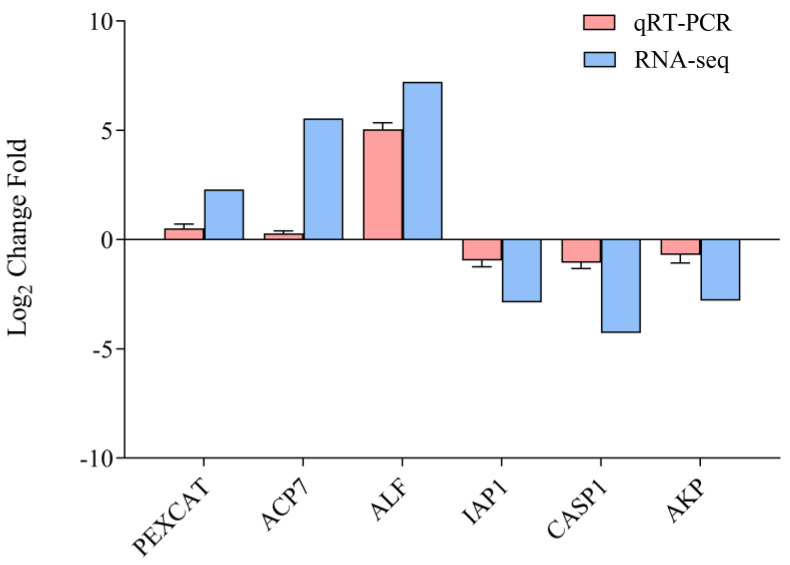 Figure 4
Figure 4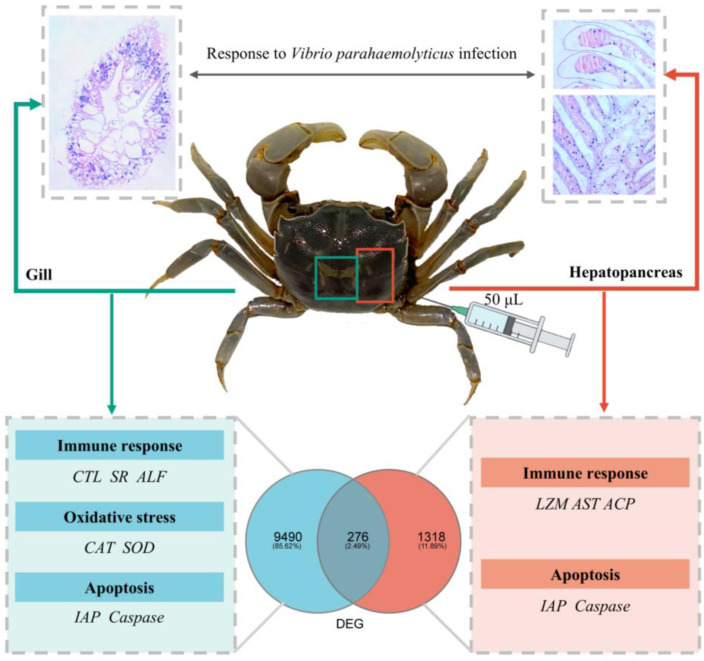 Figure 5
Figure 5- —National Natural Science Foundation of China
- —Qinglan project of Jiangsu Province to Wang Zhengfei
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInvertebrate Immune Response Mechanisms · Vibrio bacteria research studies · Aquaculture disease management and microbiota
1. Introduction
H. tientsinensis (Brachyura: Grapsoidea: Varunidae) is widely distributed along the coast of the West Pacific, primarily inhabiting intertidal mudflats, swamps, salt marshes, and estuarine areas, and it is a key component of the Yellow River Delta ecosystem, possessing significant ecological and economic value [1,2]. H. tientsinensis plays a crucial role in ecological restoration and maintaining ecological balance. H. tientsinensis mainly feeds on the Phragmites australis, Spartina alterniflora, and Suaeda salsa in the mudflat [1,3]. The invasion of S. alterniflora has increased the amount of plant organic debris, caused an increase in the number of macrobenthos feeding on plant organic debris, reduced the biomass of benthic microalgae, affected the survival of small benthos feeding on microalgae, and destroyed the biodiversity of the mudflat [4,5]. H. tientsinensis helps suppress the growth of the invasive species S. alterniflora by consuming its seedlings [6,7]. Additionally, H. tientsinensis continuously scrapes the surface of the soil in salt marsh habitats, removing S. alterniflora detritus. This activity significantly reduces the organic matter content in the soil, limits the density of macrobenthos, and prevents S. alterniflora from occupying the ecological niches of other species, thereby contributing to the stability of the ecosystem [8,9,10]. As an integral part of the coastal ecosystem, H. tientsinensis not only consumes plants growing in the intertidal zone, such as Suaeda salsa, Salicornia europaea, Phragmites australis, and S. alterniflora, as well as microorganisms in seawater, but it also serves as an important food source for various migratory bird species, including gulls, curlews, cranes, and carnivorous fish in the coastal wetlands, at their stopover or wintering sites. H. tientsinensis is known for its delicious meat, which is highly favored by coastal populations and holds considerable economic value [11].
Crustaceans lack the acquired immune system found in vertebrates and primarily rely on innate immunity to defend against various pathogenic microorganisms in their environment [12,13]. Bacteria are the most widespread and abundant pathogens causing diseases in marine shrimp and crab populations, with species of Vibrio serving as the primary bacterial pathogens responsible for infections in crustaceans [14,15]. Pathogenic vibrios, such as V. parahaemolyticus, Vibrio harveyi, and Vibrio alginolyticus, are significant contributors to these infections [16,17]. V. parahaemolyticus, a naturally occurring halophilic Gram-negative bacterium, is widely distributed in marine and estuarine environments and poses a serious threat to economically important shrimp and crab species. Infections caused by V. parahaemolyticus in Litopenaeus vannamei include hepatopancreatic damage and are characterized by clinical symptoms such as hepatopancreatic damage, lesions in the stratum corneum or epithelial tissues, disruption of structural barriers, softening of the shell, intestinal atrophy or discontinuity, loss of internal contents, and pale discoloration, often resulting in mortality rates as high as 100% [18,19]. Previous studies have demonstrated that V. parahaemolyticus accelerates energy consumption in the hepatopancreas [20,21], causing significant damage to the organ, leading to intestinal atrophy, and resulting in the permeation of endotoxins from bacterial cell walls into tissues [22]. Ultimately, this results in hepatopancreatic failure and necrosis, as evidenced by pathological changes in Eriocheir sinensis [20]. Additionally, V. parahaemolyticus impedes the growth of Portunus trituberculatus, and causes difficulties during the molting process [23]. Moreover, the synthase activity regulating the amino acid metabolic pathway in Scylla paramamosain is inhibited following V. parahaemolyticus infection [24], leading to significant economic losses in coastal aquaculture and fisheries [25,26].
Transcriptome analysis is a highly effective method for conducting systematic studies of gene function and is also a valuable approach for exploring gene expression levels, providing a comprehensive overview of complex biosynthesis processes [27]. While studies on the genomics of V. parahaemolyticus in common economic crustaceans have been reported, the immune response mechanisms of H. tientsinensis against V. parahaemolyticus remain unclear. This study aims to investigate the tissue morphology and differential gene expression in the gills and hepatopancreas of H. tientsinensis following V. parahaemolyticus infection, through pathological tissue sections and transcriptome analysis, to identify key genes associated with V. parahaemolyticus infection in H. tientsinensis. Investigating the changes in metabolism and immune responses of V. parahaemolyticus infection in H. tientsinensis will contribute to a deeper understanding of the innate immune system. This research will enrich the knowledge of crustacean mechanisms against Vibrio infections and provide theoretical guidance for scientific aquaculture practices.
2. Materials and Methods
2.1. Experimental Samples
H. tientsinensis (Carapace length: 2.5–2.9 cm, Carapace width: 3.2–3.7 cm, body weight: 18.1–25.0 g) male samples (n = 20) were collected from the coastal tidal flats (33.27578964° N, 120.79251467° E) of Dafeng Port in Yancheng, Jiangsu Province, China, along with soil and water samples from the surrounding area. The samples were temporarily maintained in a simulated tidal flat environment at 25 °C in the laboratory. During the maintenance period, water and food were supplemented as needed, and deceased samples were promptly removed.
2.2. V. parahaemolyticus Infection and Sample Collection
The V. parahaemolyticus strain (ATCC 17802) used in this study was obtained from the China General Microbiological Culture Collection Center (CGMCC) and stored in a −80 °C ultra-low temperature freezer (Thermo Fisher Scientific, Waltham, MA, USA). V. parahaemolyticus was revived by culturing in 3% NaCl nutrient agar liquid medium (10 g peptone, 5 g beef extract, 30 g NaCl dissolved in 1 L ddH_2_O, pH = 7.0) until an OD_600_ reached 0.6. Then, the bacterial cells were centrifuged at 4000 rpm for 10 min and suspended in phosphate-buffered saline (PBS) for collection. Mature crabs were randomly divided into two groups: the control group (PBS group) and the experimental group (V. parahaemolyticus infection group, VP group). Each group consisted of three biological replicates. In the experimental group, 50 μL of V. parahaemolyticus (2.76 × 10^7^ CFU/mL) was injected into the third walking leg joint membrane of each crab. The control group received the same volume of PBS. 48 h after injection, the gill and hepatopancreas tissues were collected for dissection. One portion was fixed in 4% paraformaldehyde (Biosharp, Chengdu, Sichuan, China) at 4 °C for subsequent histopathological analysis, while the other portion was snap-frozen in liquid nitrogen and stored at −80 °C for RNA extraction and transcriptome sequencing.
2.3. Histopathological Investigation
The fixed tissues of H. tientsinensis were cut into appropriately sized pieces (approximately 1 cm) using a surgical blade in the fume hood (Fuxia, Shaoxin, Zhejiang, China). The tissues were then subjected to a series of graded ethanol dehydration steps followed by paraffin embedding [28]. The embedded paraffin blocks were sectioned into 6 μm-thick sections using a microtome (Leica, Wetzlar, Germany). The paraffin sections were stained with Hematoxylin and Eosin (H&E), and the morphological features and pathological changes in the gill and hepatopancreas tissues were observed and photographed under an optical microscope (Leica, Wetzlar, Germany).
2.4. RNA Extraction, cDNA Library Construction, and Sequencing
Total RNA from the gill and hepatopancreas tissues was extracted using the RNAprep Pure Tissue Kit (Tiangen, Beijing, China) according to the manufacturer’s procedure. The RNA concentration and purity were measured using a Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), and RNA integrity was verified by 1.5% agarose gel electrophoresis. The statistical power for our RNA-seq data was calculated using the RNA-Seq Power Calculator [29]. Three biological replicates were employed to achieve the required statistical power. cDNA libraries were generated using the Illumina Truseq™ RNA Sample Prep Kit (Illumina, San Diego, CA, USA). Subsequently, all sample libraries were sequenced on the Illumina HiSeq/MiSeq platform (Illumina, San Diego, CA, USA), with sequencing conducted by Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China).
2.5. Transcriptome Assembly and Functional Annotation
After high-quality RNA-seq sequencing data were obtained, reads containing poly-N reads (>10% reads) or low-quality reads (<20 bp reads, q-value < 20) were removed using SeqPrep (version 0.1) (https://github.com/jstjohn/SeqPrep, accessed on 5 May 2023) and Sickle (version 1.33) (https://github.com/najoshi/sickle, accessed on 10 May 2023) to obtain the clean reads. De novo assembly of all clean data was performed using Trinity software (version 2.15.2) (https://github.com/trinityrnaseq/trinityrnaseq/wiki, accessed on 3 June 2023). The assembly results of the initial reads were then evaluated, and redundant reads were removed using TransRate software (version 1.0.3) (http://hibberdlab.com/transrate/, accessed on 10 June 2023) and CD-HIT software (version 4.6.2) (https://github.com/weizhongli/cdhit/archive/V4.6.2.tar.gz, accessed on 12 June 2023). The transcriptome was evaluated using BUSCO software (version 6.0.0) (Benchmarking Universal SingleCopy Orthologs, http://busco.ezlab.org, accessed on 20 June 2023) to ensure the completeness of the assembly. All transcripts obtained from RNA-seq sequencing were compared against six major databases: non-redundant protein sequence (NR, https://www.ncbi.nlm.nih.gov/public/, accessed on 4 July 2023); Swiss-Prot (https://ucrania.imd.ufrn.br/~pitagoras/protein_dimension_db/release_1/uniprot_sorted.fasta.gz, accessed on 5 July 2023); Pfam (https://pfam.sanger.ac.uk/, accessed on 6 July 2023); EggCOG (http://eggnogdb.embl.de/#/app/home, accessed on 10 July 2023); Gene Ontology database (GO, https://www.geneontology.org, accessed on 19 July 2023); and Kyoto Encyclopedia of Genes and Genomes (KEGG, https://www.genome.jp/kegg/, accessed on 5 August 2023). BLAST+ (https://ftp.ncbi.nlm.nih.gov/blast/executables/blast+/LATEST/, accessed on 15 September 2023) was used for obtaining the annotation information from each database, and statistics were compiled for each database’s annotation results.
2.6. Analysis of DEGs
The expression levels of genes/transcripts were analyzed using RSEM (RNA-Seq by Expectation-Maximization) software (version 1.3.3) (https://github.com/deweylab/RSEM, accessed on 5 April 2024) to define the differential expression between samples. Meanwhile, the functional annotation of transcripts was integrated to uncover the regulatory mechanisms of genes. TPM (Transcript per million) data for transcripts and genes were obtained by correcting gene length variations across samples, caused by differential transcript usage, using the tximport 1.10.0 R package and the “length Scaled TPM” tool. Additionally, the corrected read count data for each gene was used to evaluate gene expression levels in terms of fragments per kilobase of transcript per million fragments mapped (FPKM). The corrected read count data for each gene were then imported into EdgeR (version 4.7.5) (http://bioconductor.org/packages/stats/bioc/edgeR/, accessed on 10 September 2024) for statistical comparison between experimental and control groups. DEGs were considered when the false discovery rate (FDR) was ≤0.05 and the FPKM was ≥1. GO functional and KEGG pathway enrichment analyses were performed to map all DEGs to different categories, aiming to understand the functions and biological pathways involved. GO functional enrichment analysis was conducted using Goatools (version 1.4.12) (https://github.com/tanghaibao/Goatools, accessed on 5 October 2024), with a p-value threshold of 0.05. KEGG pathway enrichment for candidate genes was analyzed using KOBAS (version 3.0) (http://bioinfo.org/kobas/, accessed on 15 October 2024), and statistical significance was considered when FDR ≤ 0.05.
2.7. qRT-PCR Validation
To validate the RNA-seq results by qRT-PCR, six DEGs were randomly selected from the PBS and VP groups, including peroxisomal catalase-like (PXCAT), acid phosphatase type 7-like (ACP7), anti-lipopolysaccharide factor-like (ALF), inhibitor of apoptosis protein 1 (IAP1), caspase-1-like (CASP1), and alkaline phosphatase-like (AKP). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was employed as the reference gene. The same RNA samples used for RNA-Seq library construction were reverse transcribed into cDNA libraries. All qPCR primers were designed using Primer Premier 5.0 software and listed in Table S1. The qRT-PCR reactions were performed in a 20 µL reaction volume, consisting of 10 µL of 2 × FastReal qPCR PreMix (TIANGEN, Beijing, China), 0.6 µL of forward and reverse primers, 2 µL of 50 × cDNA, 0.4 µL of ROX, and 6.4 µL of RNase-free ddH_2_O. The reactions were then run on an ABI QuantStudio 3 quantitative PCR instrument (Applied Biosystems, Foster City, CA, USA). The thermal cycling conditions were set as follows: 95 °C for 2 min, 45 cycles at 95 °C for 5 s, 55 °C for 10 s, and 72 °C for 30 s. The relative expression levels of different genes were calculated according to the 2^−ΔΔCT^ method [30]. Three biological replicates were included for each gene.
3. Results
3.1. Histopathological Evaluation
After 48 h of V. parahaemolyticus infection, the gills of H. tientsinensis displayed distinct changes. In the PBS control group, the gill filaments were neatly arranged, with no signs of gill cavity dilation or rupture. The blood cells of the gill filaments were separated by pillar cells, and the marginal vessel was obvious (Figure 1a,c). In the VP experimental group, the gill cavity was dilated, gill filaments were ruptured, the structure of the blood cells and the pillar cells became blurred, the blood cells were aggregated in the gill lobules, the number of blood cells was increased, and the end was enlarged (Figure 1b,d). In the hepatopancreas tissue, the lumen morphology of the hepatopancreas tubules in the PBS control group was normal, maintaining a stellate shape, with well-defined connective tissue and evenly distributed nuclei located near the outer edge of the hepatopancreas tubules (Figure 1e,g). In the hepatopancreas tubules of the VP experimental group, nuclei collapsed, and the detachment of the basement membrane occurred. The star-shaped structure of the hepatopancreas expanded, the folds contracted, and in some cases, the lumen space of the hepatopancreas tubules increased. Epithelial cells atrophy, disappearance of the stellate shape, as well as the enlargement and increase in vacuoles in hepatopancreas tubules were seen (Figure 1f,h).
3.2. De Novo Assembly and Annotation of Unigenes
The total number of raw reads from both the control and experimental groups was confirmed through RNA sequencing (Table S2). After the removal of poly-N sequences and low-quality reads, the average percentage of clean reads obtained from each sample was 98.47%. Mapping to the reference genome, the average mapping ratio at the transcript level for each sample was 83.39%. Subsequently, de novo assembly of the clean reads was performed using Trinity software, resulting in a total of 95,878 reads, of which 13,014 reads exceeded 1500 bp in length (Table S3). PCA analysis was conducted on transcriptome sequencing data from gill and hepatopancreatic tissues. The four groups (PBS group of gills, VP group of gills, PBS group of hepatopancreas, VP group of hepatopancreas) exhibited a completely separated quadrant distribution (Figure S1). A total of 39,743 non-redundant unigenes were annotated in public databases. Of these, 20,962 were assigned to the GO database, 19,668 to the KEGG, 24,074 to eggNOG, 36,689 to the NR protein database, 19,375 to Swiss-Prot, and 20,793 to Pfam (Table 1).
In GO database, those unigenes were classified into three categories: biological process (BP), cellular component (CC), and molecular function (MF). The most prevalent terms in the BP category were cellular process (8266 unigenes) and metabolic process (7817 unigenes). In the CC category, cell part (8315 unigenes) and membrane (6031 unigenes) were the dominant terms. For the MF category, binding (10,675 unigenes) and catalytic activity (11,024 unigenes) were the most abundant subcategories (Figure S2a). A total of 19,668 unigenes, primarily involved in seven categories including Metabolism, Genetic Information Processing, Environmental Information Processing, Cellular Processes, Organismal Systems, Human diseases, and Drug Development, were mapped to 46 pathways in the KEGG database, with the three most enriched pathways being Infectious disease: Viral, Translation, and Signal transduction (Figure S2b).
3.3. Classification and Analysis of DEGs
To further analyze and characterize the DEGs, a threshold was applied to select DEGs with a significance level of less than 0.05 (Padjust < 0.05) and gene expression changes of at least 2-fold (|log2FC| ≥ 1). After the injection of V. parahaemolyticus into the H. tientsinensis, 9766 DEGs were detected in the gills (4687 upregulated and 5079 downregulated) (Figure 2a; Table 2), while 1594 DEGs were identified in the hepatopancreas (834 upregulated and 760 downregulated) (Figure 3a, Table 3). Among these DEGs, 276 were significantly upregulated or downregulated in both the gill and hepatopancreas tissues.
3.4. GO and KEGG Analysis of DEGs
GO annotation analysis revealed that the 9766 DEGs in the gill were classified into three categories, with a total of 45 functional groups: 19 groups in biological process (predominantly associated with cellular and metabolic processes), 12 in cellular component (primarily linked to cell parts, organelles, and protein-containing complex), and 14 in molecular function (chiefly involved in binding, catalytic activity, and transporter activity) (Figure 2b). The 1594 unigenes identified in the hepatopancreas were classified into three main categories across 39 functional groups. These categories included biological processes (16 functional groups), cellular components (12 functional groups), and molecular functions (11 functional groups) (Figure 3b). Analysis of biological processes identified that a substantial proportion of DEGs were predominantly involved in cellular, metabolic, and signaling processes. Investigation of cellular components revealed that over fifty percent of the DEGs were associated with cell parts, membranes, and components of membrane structures. In terms of molecular functions, DEGs were chiefly linked to binding, catalytic activity, and transporter activity.
The KEGG enrichment analysis revealed significant enrichment of pathways related to ribosome, coronavirus disease COVID-19, and systemic lupus erythematosus in the gill (Figure 2c). In contrast, the DEGs in the hepatopancreas were significantly enriched in pathways associated with cardiac muscle contraction, chemical carcinogenesis DNA adducts, and glycine, serine, and threonine metabolism (Figure 3c).
3.5. Validation of RNA-Seq Data by qRT-PCR
Six genes from the RNA-seq data were selected for qRT-PCR analysis. The qRT-PCR expression patterns of these six genes (PXCAT, ACP7, ALF, IAP1, CASP1, and AKP) showed consistent agreement with RNA-seq results (Figure 4). This indicates that the expression trends of the genes measured through RNAseq were accurate and reliable.
4. Discussion
The gill, serving as the respiratory organ of crustaceans, acts as a direct interface between the organism and its environment. When crustaceans are exposed to pathogens and toxins, the gill, as the first line of defense, directly encounters the pathogens and activates immune-related metabolic pathways [31]. Additionally, the gill plays a crucial role in respiration, osmoregulation, excretion of nitrogenous wastes, and maintenance of acid-base balance, making it a vital organ to consider in toxicological analyses [32,33], the hepatopancreas of crustaceans, analogous to the liver, pancreas, and intestine of vertebrates, plays an essential role in metabolism, nutrient absorption, and immune function [34]. As the primary detoxification organ for exogenous substances, the hepatopancreas also plays a significant role in oxidative stress [35].
This study found that after V. parahaemolyticus infection for 48 h, significant damage was observed in the gill and hepatopancreas tissues of H. tientsinensis. According to the DEGs analysis, the expression of Aspartate aminotransferase (AST) was significantly upregulated in the hepatopancreas of the experimental group of H. tientsinensis following infection with V. parahaemolyticus, while both AST gene and Alanine aminotransferase (ALT) gene expressions were significantly increased in the gill tissue. These findings are consistent with results from E. sinensis infected by V. parahaemolyticus, where changes in the gene expression levels of ALT and AST were also reported. ALT and AST proteins are transferase enzymes [36], and the significant increase in their activities indicates that tissue damage occurred in the crab. DEG enrichment analysis revealed that significantly altered genes were associated with immune response, oxidative stress, and apoptosis. Several GO terms, including transporter activity (GO:0005215), immune system process (GO:0002376), and biological regulation (GO:0065007), were linked to immune responses. Additionally, the KEGG enrichment analysis highlighted significant enrichment of DEGs in immune and apoptosis-related pathways, such as phagosome (K06563) and lysosome (K19363).
4.1. Immune-Related Genes
After 48 h of V. alginolyticus infection in H. tientsinensis, the expression level of Lysozyme gene (LZM) in the gill was significantly upregulated, while a similar molecule of LZM in the hepatopancreas was found to be significantly downregulated. LZM is an alkaline protein capable of lysing bacterial cell walls and exhibiting a strong inhibitory effect on V. alginolyticus and V. parahaemolyticus [37,38]. LZM is also an antimicrobial peptide (AMP) that is capable of killing and clearing invading bacteria [39,40]. The downregulation of LZM gene expression in the hepatopancreas was also observed 12 h after V. parahaemolyticus infection in E. sinensis [20]. Based on these observations, it is speculated that the immune-related mechanisms in the hepatopancreas respond rapidly to pathogen invasion, and LZM activity is suppressed in the later stages of infection. In contrast, the increase in LZM activity in the gill of H. tientsinensis suggests that the immune defense mechanisms in crustaceans may be activated during the early contact phase with pathogens and continue to respond to immune challenges in order to resist the invasion of exogenous substances.
In addition, another antimicrobial peptide, ALF, which plays a crucial role in the innate immunity of crustaceans [41,42], was found to be significantly upregulated in the gill of E. sinensis following microbial infection. In the present study, two types of AMP genes (LZM and ALF) were upregulated after V. parahaemolyticus infection, suggesting that both LZM and ALF may play key roles in the immune defense against microbial infection in H. tientsinensis.
Noteworthy, the expression level of acid phosphatase type (ACP) gene, which encoded an enzyme associated with non-specific immune response, was significantly upregulated in the gill of H. tientsinensis in the experimental group. In contrast, the expression levels of the ACP and AKP gene were significantly reduced in the hepatopancreas of the experimental group. ACP plays an essential role in various important immune defense activities [43] and is involved in the transfer and metabolism of phosphate groups [44]. In crustaceans, AKP is also an important metabolic-regulatory enzyme, working in conjunction with other hydrolases to form a crucial detoxification system [45]. Both ACP and AKP regulate cellular metabolism within the immune defense system [46,47]. Following V. alginolyticus infection in L. vannamei, the expression of the AKP gene in the hepatopancreas was found to be significantly lower than in the control group [48]. In E. sinensis, after infection with V. parahaemolyticus, the expression level of ACP in the hepatopancreas was significantly higher in the experimental group compared to the control group [20]. Based on these findings, it is hypothesized that ACP in the gill of H. tientsinensis is involved in immune regulation, accelerating the metabolism and transport of immune-related substances. However, the activity of metabolic-regulatory enzymes and the function of the detoxification system in the hepatopancreas are somewhat inhibited following pathogen infection.
Following V. parahaemolyticus infection in H. tientsinensis, the expression levels of Scavenger receptors (SR) gene and C-type lectin domain family 6 member A-like (CTL) gene were significantly upregulated in the gill of the experimental group. SR is a typical subfamily of pattern recognition receptors (PRRs) [49,50,51], while CTL is a broad family of PRRs [52]. High expression of SR was observed in the hepatopancreas and gill of E. sinensis, where it binds to both Gram-negative and Gram-positive bacteria, enhancing AMP production and phagocytosis during bacterial invasion [53,54]. Following an attack by Gram-negative bacteria, E. sinensis CTLs are significantly upregulated [55]. These findings suggest that SR and CTL in the gill of H. tientsinensis are involved in immune recognition, antimicrobial activity, microbial binding, and immune regulation, playing a critical role in innate immunity. After a pathogen attack, PRRs in the gill of H. tientsinensis activate corresponding immune pathways, inducing the upregulation of AMP expression and contributing to the host’s innate immune defense.
The DEG analysis revealed that the expression level of Lipopolysaccharide-induced TNF-α factor (LITAF) was significantly upregulated in the gill of H. tientsinensis. LITAF is an immune-related gene induced by lipopolysaccharide (LPS) and plays a critical role in immune regulation in response to Vibrio species and viruses [56]. LITAF regulates inflammatory and immune responses and participates in the expression of various immune-related factors [57]. In crustaceans, the activation and upregulation of LITAF were associated with antibacterial and antiviral immune responses. Previous studies have demonstrated that, following V. alginolyticus infection, the expression of LITAF in the gill, intestine, and hepatopancreas of L. vannamei was significantly upregulated [58]. Additionally, after V. parahaemolyticus infection, the expression of LITAF was significantly elevated in L. vannamei [59]. In this study, LITAF was annotated in the lysosome (K19363) signaling pathway. The lysosome pathway integrates various metabolic tasks, including degradation, transport, phagocytosis, and autophagy, to maintain cellular homeostasis [60]. The upregulation of LITAF expression in the gill of H. tientsinensis is suggested to activate its innate immune response. It plays a significant role in regulating inflammatory responses and in antibacterial and antiviral immune defense mechanisms.
4.2. Oxidative Stress-Related Genes
Previous studies have shown that Vibrio infection leads to the dysregulation of the antioxidant system in crustaceans [61], resulting in oxidative stress. Oxidative stress is characterized by an imbalance between oxidation and antioxidation within the organism. Excessive oxidative stress can induce or suppress the expression of related factors, leading to inflammatory responses and causing oxidative damage.
In the experimental group of H. tientsinensis, the gene expression levels of Superoxide dismutase (SOD) and Catalase (CAT) were significantly upregulated in gills. SOD and CAT are key components of the enzymatic antioxidant system and are often selected as indicators of oxidative stress [62,63]. Both SOD and CAT participate in the innate immune response of invertebrates. Specifically, SOD catalyzes the conversion of O_2_ to H_2_O_2_ in response to bacterial challenge [64,65]. Conversely, CAT decomposes H_2_O_2_ into harmless H_2_O and O_2_ [66]. Following V. parahaemolyticus infection, the gene expression level of SOD in the hepatopancreas of E. sinensis was significantly increased [20]. Differences in the expression of non-specific immune-related enzymes, such as AKP and ACP, indicate an imbalance in the antioxidant system in crustaceans [20,37]. The significant upregulation of SOD and CAT expression, along with the notable increase in ACP expression in the gill of H. tientsinensis, suggests that V. parahaemolyticus infection induces oxidative stress in the gill tissue, leading to oxidative damage.
4.3. Genes Related to Apoptosis and Pyroptosis
Caspases are a family of proteases that participate in various critical biological processes, including apoptosis and inflammation [67]. In this study, necrosis and blurred cellular structures were observed in the gill of H. tientsinensis in the experimental group (Figure 1). Furthermore, there was a significant upregulation of Caspase-like (Caspase 3) gene expression in the gill. Caspase 3 is a conserved cysteine protease family that plays a pivotal role in apoptosis. Activated caspase 3 cleaves a series of proteins, ultimately leading to cell apoptosis [68,69]. After infection, a significant upregulation of apoptotic genes was observed in the gill.
Compared to the control group, the gene expression levels of the IAP 1 and baculoviral IAP repeat-containing protein 7-like gene were significantly downregulated in the hepatopancreas of the experimental group in H. tientsinensis. Inhibitors of apoptosis proteins (IAPs) regulate the balance between cell proliferation and cell death by inhibiting caspase activities and mediating immune responses [70,71]. In E. sinensis, following LPS and bacterial stimulation, IAP expression increases in hemocytes, activating Caspase 3/7 and inducing apoptotic pathways [72]. IAP activity was inhibited in the hepatopancreas of H. tientsinensis, whereas Caspase 3/7 exhibited no significant differential expression. This suggested that after V. parahaemolyticus infection in H. tientsinensis, the apoptotic pathways were activated, while the anti-apoptotic pathways were suppressed in the hepatopancreas.
In the hepatopancreas of the experimental group of H. tientsinensis, the expression of CASP1 was significantly downregulated. The expression of CASP1 induces pyroptosis, a form of cell death that promotes pathogen clearance by recruiting immune cells to the site of infection [73,74,75,76]. After infection, the gene expression of CASP1 was significantly reduced in the hepatopancreas of the experimental group. Simultaneously, a significant upregulation of Cytochrome c expression was observed in the hepatopancreas. The release of cytochrome c from mitochondria is a hallmark of apoptosis [77]. As a caspase activator, cytochrome c induces caspase expression and activates apoptosis [78], which is consistent with the phenomenon of blurred tissue structure found in our study of hepatopancreas tissue section. After the host is attacked by V. parahaemolyticus, the epithelial cells atrophy and the nuclei collapse. These findings suggest that after V. parahaemolyticus infection, the hepatopancreas of H. tientsinensis experiences tissue damage, with anti-apoptotic mechanisms being suppressed and apoptotic genes being highly expressed.
5. Conclusions
The results of this study indicate that V. parahaemolyticus infection causes severe tissue damage in the gill and hepatopancreas of H. tientsinensis. In the gill, this is manifested as the proliferation and aggregation of hemocytes, with some gill filaments showing signs of breakage. In the hepatopancreas, vacuolization, basal membrane separation, and cell membrane rupture were observed. V. parahaemolyticus infection leads to the differential expression of immune-related genes (including AST, ALT, LZM, ALF, LITAF, etc.) in the gills and hepatopancreas, thereby triggering immune defense mechanisms (Table 2 and Table 3). Moreover, the expression levels of antioxidant enzymes in the gill were increased, including a rise in the activity of the non-specific immune enzyme ACP and a decrease in AKP activity, suggesting an imbalance in antioxidant system activity. Furthermore, the significant downregulation of the anti-apoptotic gene IAP and the significant upregulation of apoptotic genes, including Caspase-like and Cytochrome c, indicate that cellular programmed death caused by exogenous invaders leads to tissue damage in the gill and hepatopancreas of H. tientsinensis. In summary, we demonstrated the response mechanism of the gills and hepatopancreas of H. tientsinensis under V. parahaemolyticus infection (Figure 5). This study provides both morphological and bioinformatics data to further understand the innate immune defense mechanisms of H. tientsinensis against Vibrio species and offers theoretical insights for large-scale aquaculture practices.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhang D. Ding G. Zhang H. Tang B. Isolation and characterization of 10 microsatellite markers in Helice tientsinensis (Brachyura: Varunidae)Conserv. Genet. Resour.2009132132310.1007/s 12686-009-9074-z · doi ↗
- 2Zhang L. Lan S. Angelini C. Yi H. Zhao L. Chen L. Han G. Interactive effects of crab herbivory and spring drought on a Phragmites australis-dominated salt marsh in the Yellow River Delta Sci. Total Environ.202176614425410.1016/j.scitotenv.2020.14425433421778 · doi ↗ · pubmed ↗
- 3Qiu D. Yan J. Ma X. Luo M. Wang Q. Cui B. Microtopographical modification by a herbivore facilitates the growth of a coastal saltmarsh plant Mar. Pollut. Bull.201914043144210.1016/j.marpolbul.2019.01.06630803664 · doi ↗ · pubmed ↗
- 4Song Y. Yan C. Gao C. Xu H. Hua E. Liu X. Seasonal Distribution of Meiofaunal Assemblages in the Mangrove Tidal Flat of Futian, Shenzhen, China J. Ocean Univ. China 20222195596410.1007/s 11802-022-4869-6 · doi ↗
- 5Chen Z. Li B. Zhong Y. Chen J. Local competitive effects of introduced Spartina alterniflora on Scirpus mariqueter at Dongtan of Chongming Island, the Yangtze River estuary and their potential ecological consequences Hydrobiologia 20045289910610.1007/s 10750-004-1888-9 · doi ↗
- 6Elmer W.H. A Tripartite Interaction Between Spartina alterniflora, Fusarium palustre, and the Purple Marsh Crab (Sesarma reticulatum) Contributes to Sudden Vegetation Dieback of Salt Marshes in New England Phytopathology 20141041070107710.1094/PHYTO-08-13-0219-R 24679153 · doi ↗ · pubmed ↗
- 7Lan S.Q. Zhang L.W. Yi H.P. Xu C.L. Lu F. Feng G.H. Han G.X. Food source and feeding habit of Helice tientsinensis from the common reed vegetation in high marsh of Yellow River Delta, China Chin. J. Appl. Ecol.20203131932510.13287/j.1001-9332.202001.03431957410 · doi ↗ · pubmed ↗
- 8Reinsel K.A. Impact of fiddler crab foraging and tidal inundation on an intertidal sandflat: Season-dependent effects in one tidal cycle J. Exp. Mar. Biol. Ecol.200431311710.1016/j.jembe.2004.06.003 · doi ↗