Mono‐ and Co‐Culture Biofilms of Candida auris or Candida albicans With Staphylococcus aureus Regulate Cell Viability and Pro‐Inflammatory Cytokine Expression Differently in Oral Cancer Cell Lines
Wan NurHazirah Wan Ahmad Kamil, Munirah Mokhtar, Madia Baizura Baharom, H. M. H. N. Bandara, Mohammad S. Alrashdan, Nicola Cirillo, Mohd Hafiz Arzmi

TL;DR
The study shows that co-cultures of Candida and Staphylococcus aureus affect oral cancer cells more than mono-cultures, increasing inflammation and reducing cell viability.
Contribution
The novel finding is that co-cultures of Candida auris or Candida albicans with Staphylococcus aureus uniquely influence oral cancer cell behavior compared to mono-cultures.
Findings
Co-cultures of C. auris and S. aureus significantly increased IL-6 and IL-8 expression in oral cancer cells compared to C. albicans.
Co-culturing Candida species with S. aureus reduced cell viability and increased pro-inflammatory IL-8 expression in oral cancer cell lines.
Abstract
Candida species and Staphylococcus aureus are nosocomial pathogens associated with immunocompromised individuals, especially oral cancer patients. This study elucidates the effects of mono‐ and co‐culture biofilms of Candida albicans , Candida auris and S. aureus on cell viability and pro‐inflammatory cytokine expression in healthy epithelial cells (hTERT TIGKs) and oral cancer (ORL‐48) cell lines. Mono‐ and co‐culture biofilms of C. albicans , C. auris and S. aureus , developed using static biofilm for 72 h, were collected and filter sterilized (biofilm filtrate). Test cell growth medium (TCGM) was prepared for hTERT TIGKs and ORL‐48 by mixing 20% (v/v) biofilm filtrate with 80% serum‐free media. The cells were seeded in 96‐well plates, and TCGM was added as treatment. The unstimulated media (UM), made of 100% serum‐free media, served as control. After 24 h, cell viability…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
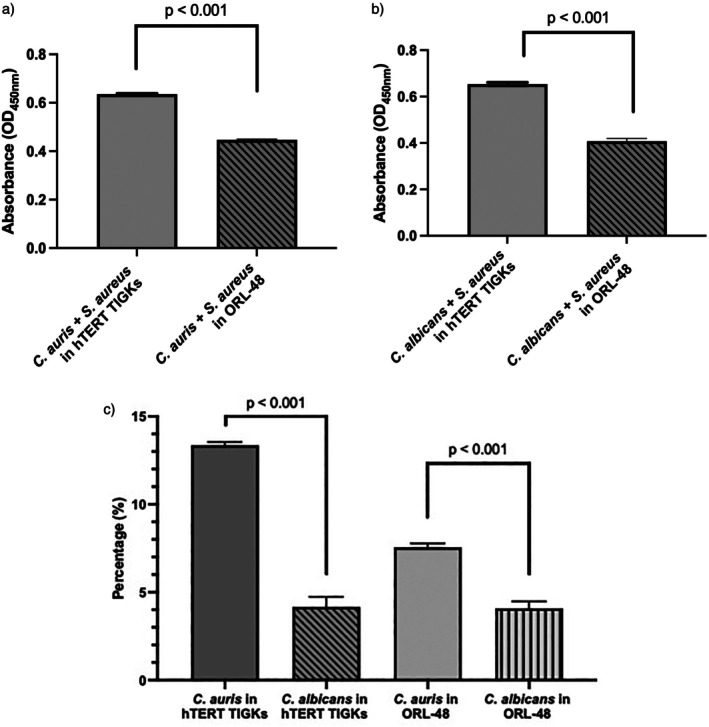 FIGURE 1
FIGURE 1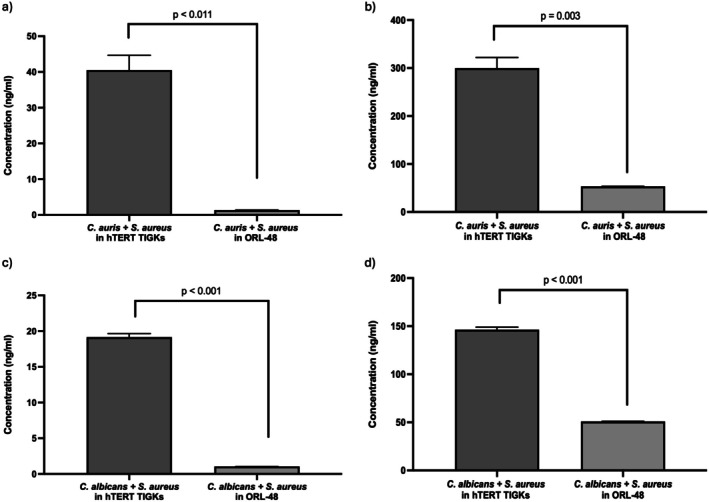 FIGURE 2
FIGURE 2| 95% CI | |||||||
|---|---|---|---|---|---|---|---|
| Cell lines | Multiple comparisons (Post hoc Dunnet T3) | Mean difference | Lower | Upper |
| ||
|
| |||||||
| hTERT TIGKs | Mono‐culture | vs. |
| 0.085 | 0.081 | 0.089 | < 0.001 |
| Mono‐culture | vs. | Unstimulated media (UM) | 0.126 | 0.122 | 0.130 | < 0.001 | |
|
| vs. | Unstimulated media (UM) | 0.041 | 0.036 | 0.046 | < 0.001 | |
| Mono‐culture | vs. |
| 0.027 | 0.016 | 0.039 | < 0.001 | |
| Mono‐culture | vs. | Unstimulated media (UM) | 0.087 | 0.077 | 0.097 | < 0.001 | |
|
| vs. | Unstimulated media (UM) | 0.041 | 0.036 | 0.461 | < 0.001 | |
| Mono‐culture | vs. | Mono‐culture | 0.039 | 0.029 | 0.049 | < 0.001 | |
|
| vs. |
| 0.018 | 0.009 | 0.028 | < 0.001 | |
| ORL‐48 | Mono‐culture | vs. |
| 0.034 | 0.031 | 0.037 | < 0.001 |
| Mono‐culture | vs. | Unstimulated media (UM) | 0.082 | 0.079 | 0.085 | < 0.001 | |
|
| vs. | Unstimulated media (UM) | 0.048 | 0.044 | 0.052 | < 0.001 | |
| Mono‐culture | vs. |
| −0.014 | −0.036 | 0.007 | 0.372 | |
| Mono‐culture | vs. | Unstimulated media (UM) | 0.023 | 0.004 | 0.042 | < 0.001 | |
|
| vs. | Unstimulated media (UM) | 0.009 | −0.007 | 0.024 | 0.500 | |
| Mono‐culture | vs. | Mono‐culture | 0.059 | 0.034 | 0.078 | < 0.001 | |
|
| vs. |
| 0.040 | 0.024 | 0.055 | < 0.001 | |
|
| |||||||
| hTERT TIGKs | Mono‐culture | vs. |
| −25.481 | −25.503 | −25.459 | < 0.001 |
| Mono‐culture | vs. | Unstimulated media (UM) | 16.491 | 16.383 | 16.600 | < 0.001 | |
|
| vs. | Unstimulated media (UM) | 41.973 | 41.868 | 42.077 | < 0.001 | |
| Mono‐culture | vs. |
| 58.717 | 58.550 | 58.884 | < 0.001 | |
| Mono‐culture | vs. | Unstimulated media (UM) | 76.136 | 76.019 | 76.253 | < 0.001 | |
|
| vs. | Unstimulated media (UM) | 17.419 | 17.298 | 17.540 | < 0.001 | |
| Mono‐culture | vs. | Mono‐culture | −59.645 | −59.668 | −59.621 | < 0.001 | |
|
| vs. |
| 24.554 | 24.396 | 24.711 | < 0.001 | |
| ORL‐48 | Mono‐culture | vs. |
| 0.601 | 0.428 | 0.774 | 0.03 |
| Mono‐culture | vs. | Unstimulated media (UM) | 0.410 | 0.277 | 0.544 | 0.04 | |
|
| vs. | Unstimulated media (UM) | 1.011 | 0.875 | 1.147 | < 0.001 | |
| Mono‐culture | vs. |
| 1.037 | 1.006 | 1.067 | < 0.001 | |
| Mono‐culture | vs. | Unstimulated media (UM) | 1.740 | 1.601 | 1.879 | < 0.001 | |
|
| vs. | Unstimulated media (UM) | 0.703 | 0.573 | 0.834 | 0.001 | |
| Mono‐culture | vs. | Mono‐culture | 1.330 | 1.302 | 1.357 | < 0.001 | |
|
| vs. |
| 0.308 | 0.138 | 0.478 | 0.14 | |
|
| |||||||
| hTERT TIGKs | Mono‐culture | vs. |
| 35.227 | 16.718 | 53.736 | 0.014 |
| Mono‐culture | vs. | Unstimulated media (UM) | 59.500 | 40.361 | 78.629 | 0.005 | |
|
| vs. | Unstimulated media (UM) | 24.268 | 21.688 | 26.849 | < 0.001 | |
| Mono‐culture | vs. |
| 2.478 | 0.145 | 4.811 | 0.043 | |
| Mono‐culture | vs. | Unstimulated media (UM) | 10.280 | 7.282 | 13.279 | 0.004 | |
|
| vs. | Unstimulated media (UM) | 7.802 | 6.393 | 9.212 | 0.002 | |
| Mono‐culture | vs. | Mono‐culture | 49.214 | 30.907 | 67.523 | 0.006 | |
|
| vs. |
| 16.466 | 14.528 | 18.404 | < 0.001 | |
| ORL‐48 | Mono‐culture | vs. |
| 30.165 | 29.622 | 30.707 | < 0.001 |
| Mono‐culture | vs. | Unstimulated media (UM) | 14.759 | 14.515 | 15.002 | < 0.001 | |
|
| vs. | Unstimulated media (UM) | 44.923 | 44.267 | 45.580 | < 0.001 | |
| Mono‐culture | vs. |
| 27.600 | 27.123 | 28.067 | < 0.001 | |
| Mono‐culture | vs. | Unstimulated media (UM) | 22.730 | 22.362 | 23.098 | < 0.001 | |
|
| vs. | Unstimulated media (UM) | 42.354 | 41.768 | 42.939 | < 0.001 | |
| Mono‐culture | vs. | Mono‐culture | −7.971 | −8.256 | −7.686 | < 0.001 | |
|
| vs. |
| 2.569 | 2.064 | 3.075 | < 0.001 | |
- —Ministry of Higher Education, Malaysia10.13039/501100003093
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntifungal resistance and susceptibility · Peptidase Inhibition and Analysis · Antimicrobial Peptides and Activities
Introduction
1
Oral squamous cell carcinoma (OSCC) is the most pervasive form of head and neck cancer and is characterised by a high mortality rate, a tendency for recurrence and a potential for metastasis [1]. The main risk factors for OSCC include tobacco use, alcohol consumption, betel quid chewing and microbial infections regarded as an emerging risk factor [2]. However, the role of oral microbial infections in the development of oral cancer and its biological mechanisms remains poorly understood. The potential carcinogenic mechanisms influenced by oral infections include cytokine production and the induction and exacerbation of chronic inflammation [2]. Microbial infections also regulate cellular viability, migration and apoptosis by disrupting the cell cycle and activating anti‐apoptotic signalling pathways [2, 3].
The interactions between Candida auris or Candida albicans and Staphylococcus aureus contribute to the development of nosocomial infections. They can also promote chronic inflammation, modulate immune responses and induce a resistant and pro‐migratory phenotype, which, in turn, contributes to tumour progression. Furthermore, these pathogens can form biofilms that modify the production of inflammatory cytokine molecules, such as interleukin (IL)‐6 and IL‐8, in both healthy and cancer cell lines [4].
Although several studies have reported interactions between C. albicans and S. aureus in oral cell models [5], the effects of intra‐ and interkingdom biofilms of the emerging nosocomial fungal pathogen, C. auris, on cell viability and pro‐inflammatory cytokines expression from oral cancer cell lines remain unknown. Therefore, this study aimed to determine the effect of mono‐ and co‐culture biofilms of Candida spp. and S. aureus in normal (hTERT TIGKs) and oral cancer (ORL‐48) cell lines, with the hypothesis that interkingdom biofilms modulate cell viability and pro‐inflammatory cytokines in oral keratinocytes.
Materials and Methods
2
Cell Lines and Culture
2.1
Normal human gingival keratinocytes immortalised with hTERT (hTERT TIGKs) were purchased from the American Type Culture Collection (ATCC) (CRL‐3397, Manassas, VA).
The oral cancer cell line ORL‐48, derived from malignant keratinocytes of the gingiva of an oral cancer patient, was obtained from Cancer Research Malaysia, Subang Jaya Medical Centre (CRM, Selangor, Malaysia). Specific culture conditions are reported in the Supporting Information S1.
Preparation of Test Cell Growth Medium (TCGM)
2.2
The mono‐ and co‐culture biofilms were developed previously, prior to the collection of TCGM as described by Wan Ahmad Kamil et al. [6]. Details for the production of biofilms are reported in the Supporting Information S1. After 72 h of biofilm formation, the biofilm filtrate was collected and filter‐sterilised using a 0.22 μM Millex Millipore syringe filter (MERCK, Germany). To prepare TCGM for hTERT TIGKs and ORL‐48, 20% (v/v) biofilm filtrate was mixed with dermal cell basal medium (PCS‐200‐030; ATCC, Manassas, VA) and DMEM/F12 (Thermo Fisher Scientific, USA), respectively.
Cell Viability Assay Using Cell Counting Kit‐8 (CCK‐8)
2.3
hTERT TIGKs and ORL‐48 cells were incubated with TCGM or unstimulated media, and the cell viability assay was undertaken as detailed in the Supporting Information S1. Absorbance was measured at 450 nm using an Infinite 200 Pro microplate reader (TECAN, Switzerland).
Pro‐Inflammatory Cytokines Expression
2.4
ELISA assay was conducted to assess the expression of pro‐inflammatory cytokines. In brief, the samples are prepared using the same method as for the CCK‐8 assay. Following the procedure, the conditioned medium was collected for ELISA and transferred into an ELISA plate to measure the concentrations of IL‐6 and IL‐8 using an ELISA (Elabscience, USA) based on the manufacturer's instructions. The procedure is detailed in the Supporting Information S1.
Statistical Analysis
2.5
All data were analysed using GraphPad Prism version 10.0 and IBM SPSS Statistics for Mac, version 29.0 (IBM Corp., Armonk, NY, USA). A two‐tailed paired t test was used to compare co‐culture in different cell lines. A one‐way analysis of variance (ANOVA) with Dunnett's post hoc test was performed to compare cell viability and cytokine expression amongst the biofilm‐conditioned media groups. A p value < 0.05 was considered statistically significant.
Results
3
Effect of Biofilm on the Cell Viability of hTERT TIGKs and ORL‐48 Oral Cell Lines
3.1
The largest number of viable cells (hTERT TIGKs and ORL‐48) was observed within 24 h treatment with mono‐culture C. auris biofilm‐conditioned media, whereas the lowest was observed when treated with mono‐culture S. aureus biofilm‐conditioned media (Table S1). After 24 h treatment with co‐culture biofilm of C. auris and S. aureus, the cell viability of hTERT TIGKs and ORL‐48 was significantly decreased as compared to mono‐culture of both C. auris (p < 0.001) and C. albicans (p < 0.001) (Table 1).
The cell viability of hTERT TIGKs when treated with co‐culture of C. auris or C. albicans with S. aureus was significantly higher than ORL‐48 cells (p < 0.001; Figure 1a,b). In addition, a significantly higher percentage reduction in the number of viable cells was observed when hTERT TIGKs and ORL‐48 cell lines were treated with C. auris co‐cultured with S. aureus compared to co‐culture of C. albicans (p < 0.001; Figure 1c).
The cell viability of hTERT TIGKs and ORL‐48 based on the absorbance at OD450 nm when treated with co‐culture of Candida with Staphylococcus aureus . Significant differences were observed between the co‐culture of Candida auris (a) or Candida albicans (b) with S. aureus in different cell lines. (c) The percentage reduction was measured by comparing biofilm‐conditioned media of S. aureus and Candida with respective mono‐culture in hTERT TIGKs and ORL‐48 cell viability. The data represent the means from three biological replicates, each consisting of three technical replicates (n = 9). The data were analysed using a two‐tailed paired t test. A significant difference in cell viability between Candida spp. in respective cell lines was observed (p < 0.05).
Effect of Biofilm‐Conditioned Media on Pro‐Inflammatory Cytokine Expression in hTERT TIGKs and ORL‐48 Cell Lines
3.2
The expression of pro‐inflammatory cytokines in hTERT TIGKs and ORL‐48 cell lines varied under different biofilm culture media conditions. Overall, both mono‐cultures and co‐culture of C. auris or C. albicans with S. aureus significantly increased IL‐6 and IL‐8 expression in hTERT TIGKs and ORL‐48 as compared to UM (Tables S2 and S3, respectively).
The expression of IL‐6 in hTERT TIGKs and ORL‐48 significantly increased when treated with co‐culture C. auris and S. aureus as compared to mono‐culture C. auris (p < 0.001; p = 0.03; Table 1), whereas the opposite was true when C. albicans was used (p < 0.001; p < 0.001; Table 1). The expression of IL‐8 in hTERT TIGKs significantly decreased during co‐culture with either C. auris or C. albicans with S. aureus as compared to mono‐culture of respective Candida spp. (Table 1). In addition, significant differences in IL‐6 and IL‐8 cytokine expression were observed between the co‐culture of C. auris or C. albicans with S. aureus in different cell lines (p < 0.05; Figure 2).
(a, c) IL‐6 and (b, d) IL‐8 cytokine expression of hTERT TIGKs and ORL‐48 based on the absorbance at OD450 nm when treated with co‐culture of Candida with Staphylococcus aureus . The data were analysed using a two‐tailed paired t test. Significant differences were observed between the co‐culture of Candida auris or Candida albicans with S. aureus in different cell lines. Data were considered significantly different when (p < 0.05).
Discussion
4
The present study revealed mono‐ and co‐culture biofilms of Candida spp. and S. aureus promote cell growth and modulate cytokine production in oral keratinocytes in a paracrine, interkingdom and cell‐specific manner.
Mono‐culture biofilms of C. auris exhibited the highest growth‐stimulating capacity in both hTERT TIGKs and ORL‐48 cell lines. In contrast, treatment with co‐culture biofilms of C. auris or C. albicans and S. aureus significantly reduced cell viability in both cell lines compared to mono‐culture of the respective Candida. This suggests that S. aureus, which produces exotoxins such as α‐hemolysin and leukocidins, can paracrinally compromise the cell viability of hTERT TIGKs [5].
The expression of IL‐6 in hTERT TIGKs and, to a lesser extent, in ORL‐48, was increased when treated with co‐culture biofilm‐conditioned media of C. auris with S. aureus compared to mono‐culture C. auris . Conversely, a reduction in the expression of IL‐6 was observed in both cell lines when treated with co‐culture of C. albicans and S. aureus , compared to mono‐culture C. albicans . The results highlight that oral biofilms formed by nosocomial pathogenic bacteria and fungi may contribute to oral mucosal inflammation and are consistent with research indicating that co‐infections and microbial interactions can exacerbate chronic diseases and could be involved in the progression of oral inflammatory diseases and oral cancer [7, 8, 9].
The present study also showed an increase of IL‐8 expression in malignant ORL‐48 cells, but not in hTERT TIGKs, when treated with co‐culture biofilms of C. auris or C. albicans with S. aureus as compared to mono‐culture of the yeast. Overall, the findings are significant in oral oncology in that they demonstrate a specific increase in IL‐8 expression, which is potentially a biomarker for oral cancer [5], when coinfected by C. auris and S. aureus biofilms. As previous studies consistently reported a higher level of IL‐8 in carcinomas directly linked with tumour angiogenesis and disease severity [10], our results suggest that interkingdom interaction might increase tumour vascularity and disease progression in oral cancer patients.
In conclusion, this study demonstrates for the first time that C. auris biofilm creates a favourable environment for cell survival and growth and that co‐culture of C. auris or C. albicans with S. aureus increases pro‐inflammatory IL‐8 expression in oral cancer cell lines, but not in non‐malignant cells. Thus, interkingdom interaction may regulate oral carcinogenesis.
Author Contributions
W.N.W.A.K.: data curation, formal analysis. M.M. and M.H.A.: methodology, project administration, resources, review and editing. M.B.B.: investigation, methodology. H.M.H.N. and M.S.A.: review and editing. W.N.W.A.K. and M.H.A.: conceptualisation, investigation, methodology, supervision, resources, visualisation. W.N.W.A.K., N.C., and M.H.A.: writing (original draft), review and editing.
Ethics Statement
This study did not involve human participants or animal subjects, and therefore, ethical approval was not required.
Consent
The authors have nothing to report.
Conflicts of Interest
The authors declare no conflicts of interest.
Peer Review
The peer review history for this article is available at https://www.webofscience.com/api/gateway/wos/peer‐review/10.1111/jop.70043.
Supporting information
Data S1: Supporting Information.
Table S1: Descriptive analysis of cell viability in hTERT TIGKs and ORL‐48 cells based on the absorbance at OD_450nm_. when treated with biofilm‐conditioned media of mono‐and co‐culture C. auris or C. albicans with S. aureus .
Table S2: Descriptive analysis of pro‐inflammatory cytokines interleukin‐6 (IL‐6) expressed in hTERT TIGKs and ORL‐48 cell lines when treated with biofilm‐conditioned media of mono‐and co‐culture C. auris or C. albicans with S. aureus .
Table S3: Descriptive analysis of pro‐inflammatory cytokines interleukin‐8 (IL‐8) expressed in hTERT TIGKs and ORL‐48 cell lines when treated with biofilm‐conditioned media of mono‐and co‐culture C. auris or C. albicans with S. aureus .
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Y. Tan , Z. Wang , M. Xu , et al., “Oral Squamous Cell Carcinomas: State of the Field and Emerging Directions,” International Journal of Oral Science 15, no. 1 (2023): 44, 10.1038/s 41368-023-00249-w.37736748 PMC 10517027 · doi ↗ · pubmed ↗
- 2X. Wang , W. Zhang , W. Wu , S. Wu , A. Young , and Z. Yan , “Is Candida albicans a Contributor to cancer? A Critical Review Based on the Current Evidence,” Microbiology Research 272 (2023): 127370, 10.1016/j.micres.2023.127370.37028206 · doi ↗ · pubmed ↗
- 3P. N. Deo and R. Deshmukh , “Oral Microbiome and Oral Cancer—The Probable Nexus,” Journal of Oral and Maxillofacial Pathology 24, no. 2 (2020): 361, 10.4103/jomfp.JOMFP_20_20.33456248 PMC 7802843 · doi ↗ · pubmed ↗
- 4A. A. Al‐Qahtani , F. S. Alhamlan , and A. A. Al‐Qahtani , “Pro‐Inflammatory and Anti‐Inflammatory Interleukins in Infectious Diseases: A Comprehensive Review,” Tropical Medicine and Infectious Disease 9, no. 1 (2024): 13, 10.3390/tropicalmed 9010013.38251210 PMC 10818686 · doi ↗ · pubmed ↗
- 5Z. Zhu , Z. Hu , S. Li , R. Fang , H. K. Ono , and D. L. Hu , “Molecular Characteristics and Pathogenicity of Staphylococcus aureus Exotoxins,” International Journal of Molecular Sciences 25, no. 1 (2023): 395, 10.3390/ijms 25010395.38203566 PMC 10778951 · doi ↗ · pubmed ↗
- 6W. N. H. Wan Ahmad Kamil , M. Zainal , M. Mokhtar , H. M. H. N. Bandara , S. G. Dashper , and M. H. Arzmi , “Aggregation and Biofilm Formation of Mono‐ and Co‐Culture Candida Species and Staphylococcus aureus Are Affected by Nutrients in Growth Media,” Biofouling 41, no. 6 (2025): 634–648, 10.1080/08927014.2025.2518281.40528537 · doi ↗ · pubmed ↗
- 7L. Huang , F. Luo , M. Deng , and J. Zhang , “The Relationship Between Salivary Cytokines and Oral cancer and Their Diagnostic Capability for Oral cancer: A Systematic Review and Network Meta‐Analysis,” BMC Oral Health 24, no. 1 (2024): 1044, 10.1186/s 12903-024-04840-3.39237889 PMC 11378403 · doi ↗ · pubmed ↗
- 8M. H. Arzmi , N. Cirillo , J. C. Lenzo , et al., “Monospecies and Polymicrobial Biofilms Differentially Regulate the Phenotype of Genotype‐Specific Oral cancer Cells,” Carcinogenesis 40, no. 1 (2019): 184–193, 10.1093/carcin/bgy 137.30428016 · doi ↗ · pubmed ↗