Manganese homeostasis modulates glucan and chitin unmasking in the opportunistic yeast Candida albicans
Manon Henry, Maria Khouas, Gabriel Théberge-Julien, Antony T. Vincent, Louis Villeneuve, Éric Rhéaume, Jean-Claude Tardif, Adnane Sellam

TL;DR
This study shows how manganese affects the cell wall of Candida albicans, influencing immune recognition and stress resistance.
Contribution
The novel role of manganese in modulating fungal cell wall antigens and stress resistance is revealed.
Findings
Manganese deficiency alters cell wall structure and unmasking of β-glucan and chitin in Candida albicans.
Manganese modulates glucanase activity to control β-glucan exposure, independent of the calcineurin pathway.
Unmasking of cell wall components reduces phagocytosis by macrophages, impacting immune evasion.
Abstract
Candida albicans is a commensal fungus and also the most prevalent human fungal pathogen. The ability of this opportunistic yeast to acquire and maintain homeostatic levels of manganese (Mn), particularly in the metal-limited host environment, is an important determinant of its fitness. Recent studies have underscored the importance of Mn acquisition through members of Smf transporters, in C. albicans virulence and its ability to withstand various stresses. In the present study, we undertook transcriptional profiling in the mutant of the Mn transporter Smf12 under restricted Mn availability to identify processes that are directly impacted by impaired Mn uptake. Our analysis revealed that smf12 displayed a transcriptional pattern suggestive of a cell wall defect, with many transcripts associated with cell wall biogenesis being differentially regulated. smf12 together with smf11, a mutant…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
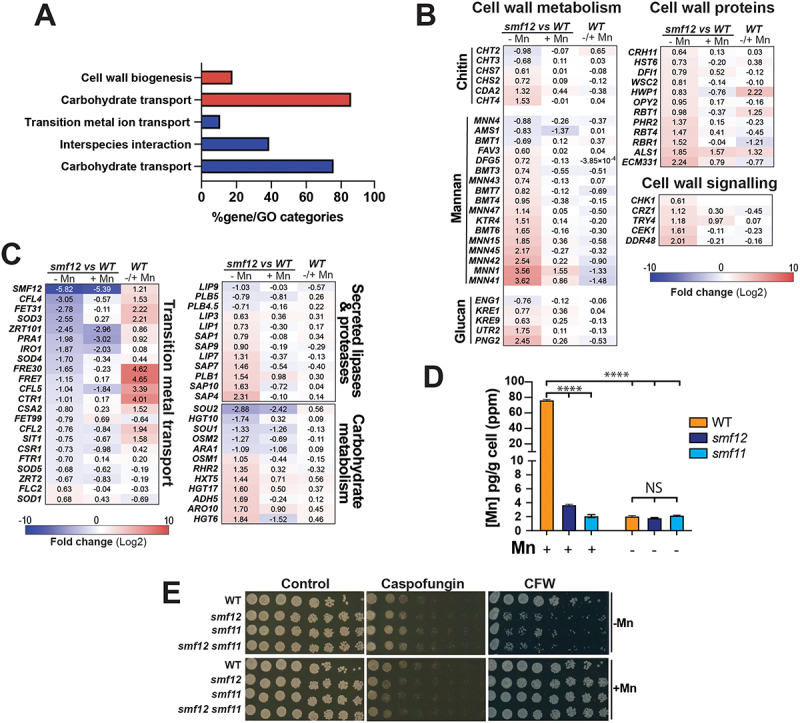 Figure 1
Figure 1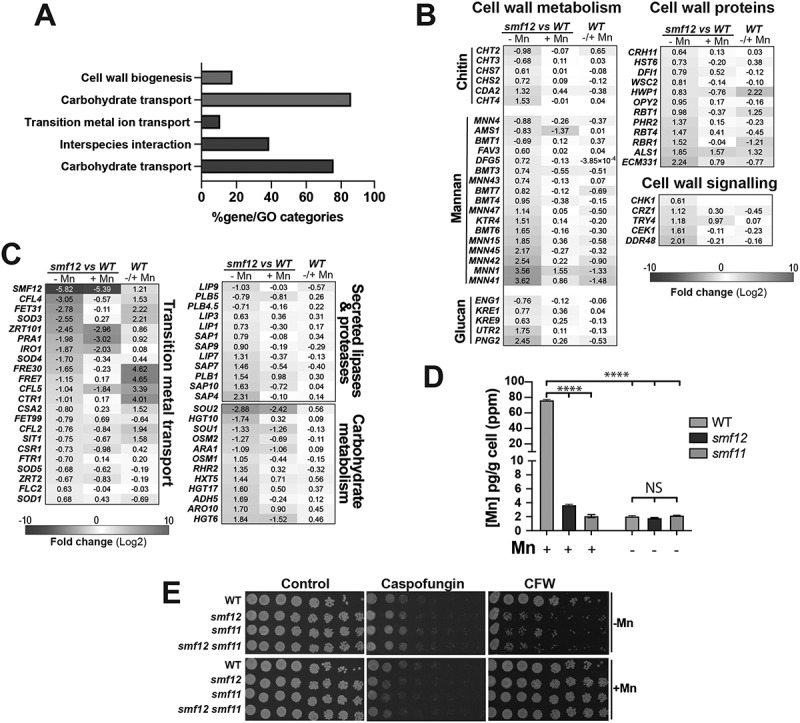 Figure 2
Figure 2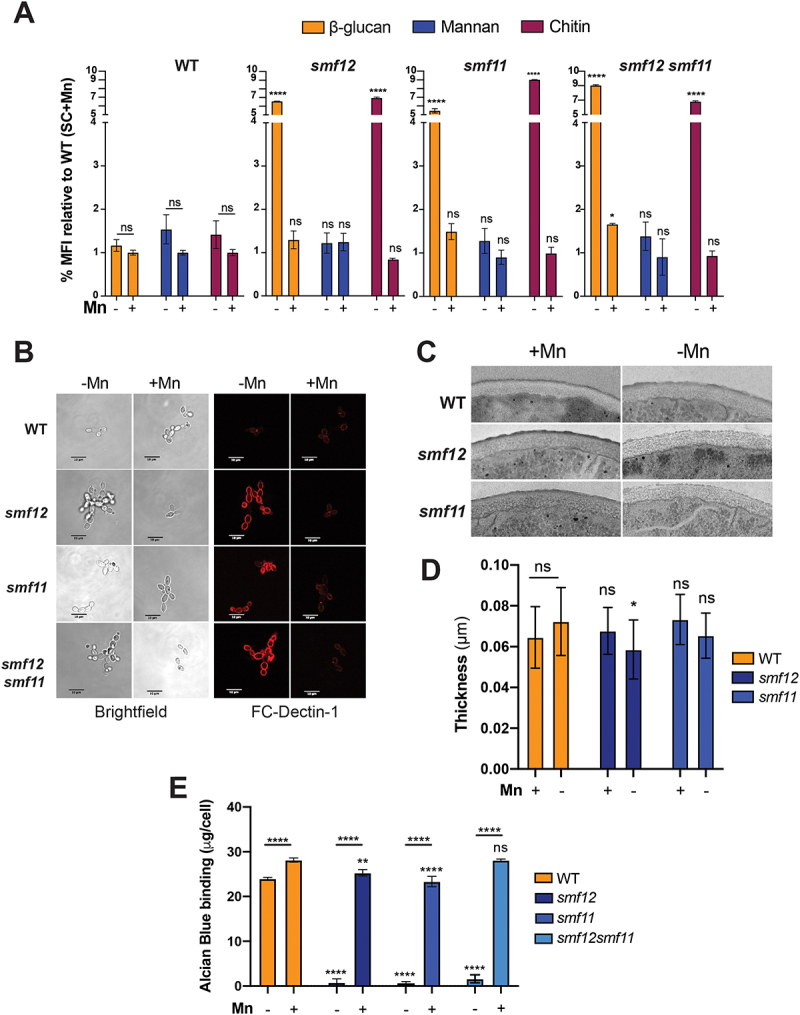 Figure 3
Figure 3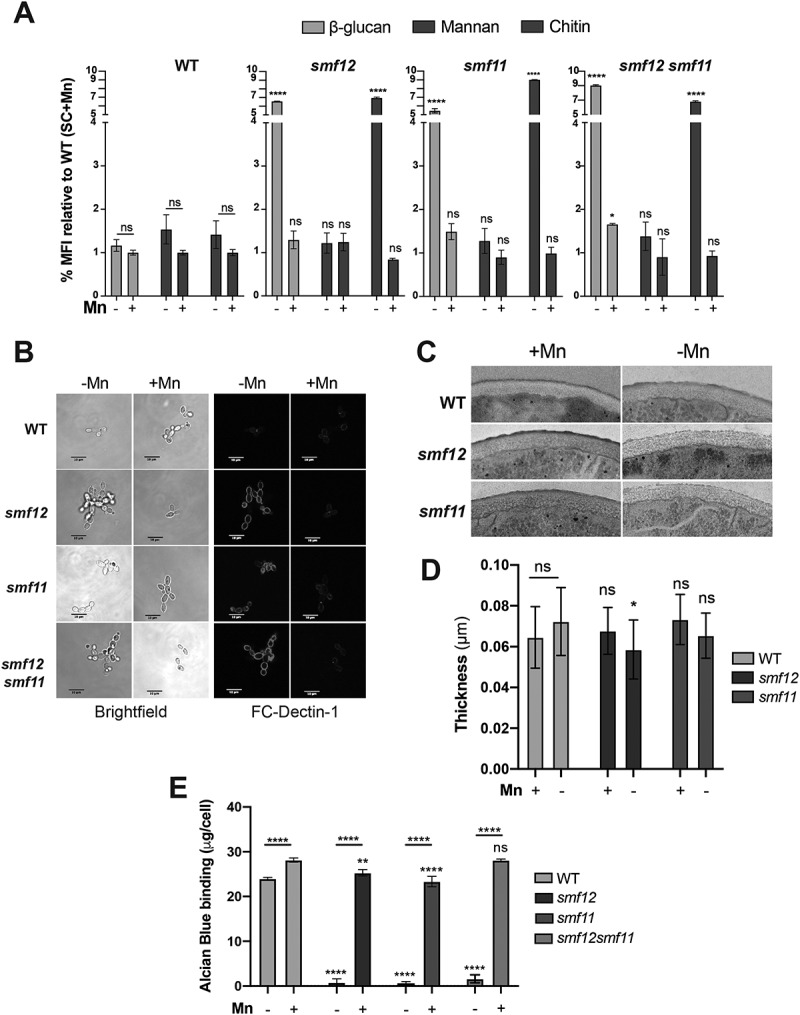 Figure 4
Figure 4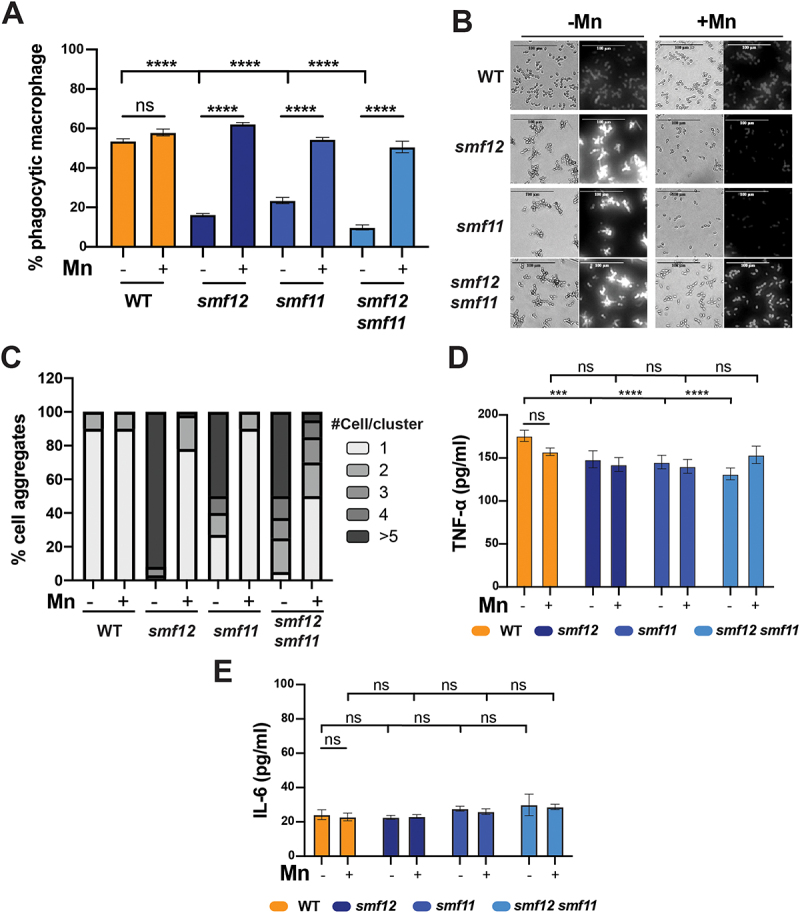 Figure 5
Figure 5 Figure 6
Figure 6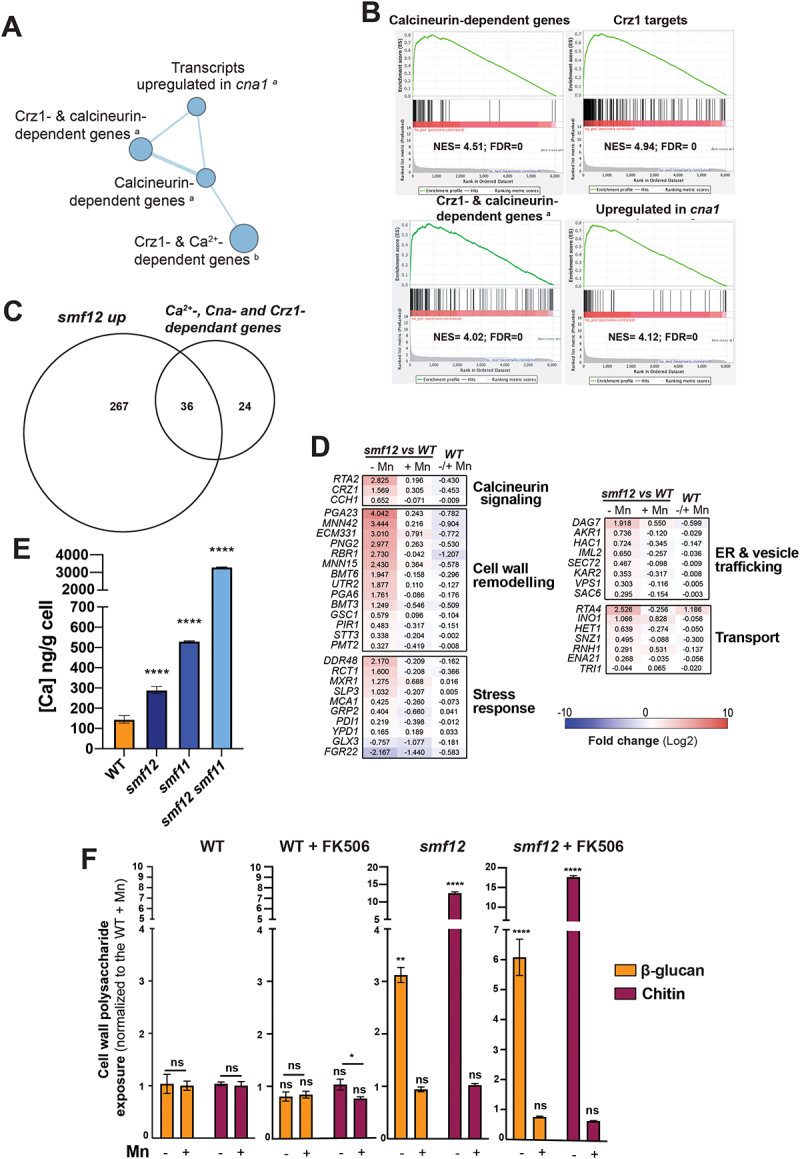 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9 Figure 10
Figure 10- —Canadian Institutes of Health Research10.13039/501100000024
- —Canada Foundation for Innovation10.13039/501100000196
- —Natural Sciences and Engineering Research Council of Canada10.13039/501100000038
- —Natural Sciences and Engineering Research Council of Canada10.13039/501100000038
- —Fondation Institut de Cardiologie de Montréal10.13039/501100012651
- —Fonds de Recherche du Québec - Santé10.13039/501100000156
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntifungal resistance and susceptibility · Fungal Infections and Studies · Trace Elements in Health
Introduction
The availability of transition metals, such as iron (Fe), copper (Cu), and zinc (Zn), is tightly regulated within the human host. To limit the proliferation of microbial pathogens, the host employs a strategy known as nutritional immunity, which involves sequestering these essential metals or directing their toxicity against pathogens [1]. However, many pathogens have evolved diverse strategies to bypass nutritional immunity, allowing them to survive and infect the host. Fungal pathogens, in particular, have developed sophisticated regulatory networks to modulate both metal acquisition and detoxification, thereby maintaining homeostasis of these essential micronutrients. For instance, in Candida albicans and other human fungal pathogens, Fe acquisition is facilitated by the high-affinity reductive and siderophore transport systems, as well as hemoglobin-Fe utilization, all of which are essential for virulence [2–5]. Similarly, Cu and Zn are internalized by specific transporters regulated at the transcriptional levels by the transcription factors Mac1 and Csr1, respectively, which are also essential for fungal fitness in vivo [6–8]. Although significant attention has been given to the mechanisms by which fungi acquire Fe, Cu, and Zn, the role of manganese (Mn) in infectious processes and the cellular mechanisms underlying Mn homeostasis in fungal cells remain partially understood.
C. albicans is a commensal fungus and an important member of the human intestinal and vaginal microbiota. Under certain conditions, including immune suppression and dysbiosis, this opportunistic yeast can cause serious infections, such as fungal sepsis, which is associated with high mortality rates [9]. While significant attention has been given to the mechanisms of Fe and Cu acquisition and their role in fungal fitness, less focus has been devoted to Mn. Recently, two independent studies have highlighted the role of Mn homeostasis in the biology and the fitness of this prevalent human fungal pathogen [10,11]. Mn uptake via the NRAMP family transporters Smf12 and Smf13 has been shown to be essential for the development of invasive hyphae and to fully support the activity of Mn-dependent superoxide dismutases, which are critical enzymes for protecting C. albicans cells from oxidative stress [10–12]. Furthermore, Mn has been shown to modulate antifungal sensitivity by regulating antifungal efflux and ergosterol homeostasis [11]. Given the critical role of Mn in these virulence-related processes, both smf12 and smf13 mutants displayed an altered virulence in various infection models [10,11].
Mn also serves as a cofactor for several metalloenzymes, including mannosyltransferases, which are essential for mannan biosynthesis and the generation of cell wall mannoproteins [13,14]. Inactivation of Mn transporters, such as Smf12 or the P-type ATPase Mn^2+^/Ca^2+^transporter Pmr1, results in a shortening of glycosyl residues linked to proteins [11,13]. Since mannoproteins are immunogenic epitopes that decorate the outer fungal cell wall, Mn homeostasis likely plays a pivotal role in modulating immune recognition. Indeed, under Mn deprivation, C. albicans cells unmask β-glucan, a major polysaccharide of the inner cell wall layer and another important fungal antigenic determinant [15]. This unmasking is likely due to the reduced density of mannan fibrils in the outer layer of the cell wall [13]. In contrast, Fe starvation leads to the opposite effect, where β-glucan is masked, reducing consequently the recognition by phagocytic macrophages [15]. This Fe-mediated masking is regulated by the cAMP-Protein Kinase A signaling pathway, along with the Fe transceptor Ftr1 and the Fe-responsive transcription factor Sef1 [15]. In contrast to Fe, however, the mechanism by which β-glucan is unmasked under Mn limitation and whether this enhances immune cell recognition remains unexplored.
In the current study, we conducted transcriptional profiling of the smf12 mutant in C. albicans under restricted Mn availability to identify processes that are directly affected by a defect in Mn internalization. smf12 mutant exhibited a transcriptional pattern reminiscent of a cell wall defect, with many transcripts related to cell wall biogenesis being differentially regulated. Accordingly, smf12 together with smf11, a mutant of the closest paralog of Smf12 [11], and their corresponding double mutant (smf12 smf11), exhibited hypersensitivity to cell wall stressors and altered cell wall ultrastructure. Consistent with the observation that Mn depletion led to increased exposure of β-glucan [15], the smf12 mutant also exhibited this effect, along with unmasking of the basal chitin layer of the inner cell wall. Intriguingly, unmasking of either β-glucan or chitin in smf mutants led to a decreased rate of phagocytosis suggesting a reduced recognition or internalization by macrophages. Moreover, we showed that Mn-mediated unmasking of β-glucan required the modulation of the glucanase activity and was not signaled through the calcineurin pathway.
Materials and methods
Strains and growth conditions
The fungal strains and the PCR primers used in this study are listed in Supplementary Table S1. C. albicans strains were routinely maintained at 30°C on synthetic complete (SC; 1.7% yeast nitrogen base, 0.5% ammonium sulfate, 2% dextrose, 0.2% amino acid, with 50 µg/ml uridine). smf deletion mutants (smf12, smf11 and the double mutant smf12 smf11) were constructed from SN148 strain [16] by replacing the entire ORF with a PCR-disruption cassette amplified from the pFA plasmids [17]. A wild-type control strain (SN148-CIp20) was created by reintroducing URA3 and HIS1 in SN148 using the integrative Cip20 plasmid [18].
Cell wall stress assays
Overnight cultures of Mn-free synthetic complete growth medium (SC-Mn) were diluted to an OD_600_ of 0.1 and 5-fold serial dilutions were prepared using the same medium. A total of 4 μl of each dilution was spotted on either SC-Mn or SC+Mn (SC-Mn with 1 mM MnCl_2_) agar medium containing 0.2 µg/ml caspofungin or 25 µg/ml calcofluor white. Plates were incubated at 30°C for 2 days.
Expression analysis by RNA-seq
RNA-seq was conducted as previously described by Henry et al. [11]. Briefly, overnight cultures of smf12 strain were diluted to an OD_600_ of 0.1 in 60 ml of fresh SC-Mn medium and grown at 30°C under agitation to early logarithmic phase (OD_600_ = 0.4). Cultures were then either supplemented (SC+Mn) or not with 1 mM MnCl_2_ and incubated at 30°C for 90 min. Cell pellets were collected by centrifugation and total RNA was extracted using an RNAeasy purification kit (Qiagen) as previously described [19]. cDNA libraries were prepared using the NEBNext Ultra II RNA Library Prep Kit and sequencing was performed using an Illumina NovaSeq 6000 sequencing system. Gene ontology (GO) analysis was performed using GO Term Finder of the Candida Genome Database [20]. The GSEA (Gene Set Enrichment Analysis) pre-ranked tool (http://www.broadinstitute.org/gsea/) was used to determine the statistical significance of correlations between the C. albicans smf12 transcriptome and different omics datasets as previously described [21]. Differentially expressed transcripts in Supplementary Table S2 were identified using Welch’s t-test with a false-discovery rate (FDR) of 5% and 1.5-fold enrichment cutoff. All RNA-seq data are available at the GEO database (https://www.ncbi.nlm.nih.gov/geo/) with the accession number GSE283948.
Quantification of β-glucan, chitin, and mannan by flow cytometry
C. albicans cells were grown in SC-Mn and SC+Mn until reaching the exponential phase and were centrifuged and fixed for 20 min using 4% paraformaldehyde (w/v, final concentration). Cells were washed three times in FACS buffer (PBS, 0.5 mM EDTA, 0.1% BSA). Three independent cell batches (each consisting of 100 µl of cell suspension at an OD_600_ = 1) were treated as follows. For β-glucan exposure, cells were stained using 5 ng Dectin-1 conjugated to IgG1 Fc domain (fc-hdec1a-2, InvivoGen) for one hour at 4°C. Cells were then washed three times in FACS buffer and incubated for one hour at 4°C with IgG Fc secondary antibody conjugated to TRITC (Invitrogen, A18822). For mannan and chitin exposures, cells were incubated for one hour at 4°C with Concanavalin A conjugated to Alexa-488 (Invitrogen, C11252) and wheat germ agglutinin (WGA) conjugated to Alexa-488 (Invitrogen, W11261), respectively. Cells were then washed three times with FACS buffer and analyzed on a BD FACSymphony A1 Cell Analyzer. To discriminate large aggregates that might affect fluorescence measurements, small size particles were detected and gated in SSC vs. FSC plots using unstained cells to adjust voltages. The mean fluorescence intensity (MFI) of stained cells was measured for the TRITC and Alexa Fluor 488 channels. Data were collected for three biological replicates for each condition and analyzed with the BD FACSDiva Software v.9.0.2. Graphs present the percentage of the MFI of 10,000 events normalized to the WT in SC+Mn.
Alcian blue binding assay
Alcian Blue binding assay was performed as previously described [13]. Briefly, cells were grown in SC+Mn or SC-Mn until reaching the exponential phase and washed with 0.02 M HCl at pH 2.5. A total of 3 × 10[7] cells were resuspended in 1 ml of Alcian Blue (30 mg/ml) diluted in 0.02 M HCl. After 10 min incubation at room temperature, cells were pelleted and the OD_620 nm_ of the supernatant was measured using a Biotek® Cytation 5 plate reader.
Phagocytosis assay and cytokines production quantification
Phagocytosis was quantified as described by Carneiro et al. [22]. Briefly, yeast cells were grown in either SC-Mn or SC+Mn to reach the exponential phase and were then fixed for 20 min using 4% formaldehyde and 0.2% Triton mixture. Fixed cells were stained with SytoxGreen 1 mM for one hour and co-cultured with macrophages J774A.1 MOI (5:1) in DMEM growth medium at 37°C and 5% CO_2_. To discriminate engulfed yeasts from those attached to macrophages, we take benefit of SYTOX-green quenching by propidium iodide (PI) (6 mg/ml) for 10 min. While phagocytosed yeasts are protected from quenching by macrophage membrane, non-phagocyted or attached yeast are stained with PI. After one hour incubation, non-phagocyted yeast were stained with propidium iodide (6 mg/ml) for 10 min and quantification was made by flow cytometry. J774A.1 cells were gated by FSC/SSC and fluorescence minus one controls (FMOC) were used to define the quadrants identifying phagocytic populations in a PI/SytoxGreen dot plot [22]. A total of 10,000 events per condition were measured and three biological replicates were considered for each condition. TNF-α and IL-6 levels in the co-culture media were measured using the mouse TNF-α (Quantikine, MTA00B-1) and IL-6 (Quantikine, RM6000B-1) ELISA kits, respectively, according to the manufacturer’s instructions.
Microscopy
Transmission electron microscopy (TEM) was used for cell wall ultrastructure imaging (Imaging- Microscopy Platform, IBIS, Université Laval, Quebec City). Yeast cells were grown in SC+Mn or SC-Mn liquid media to reach the exponential phase and were then fixed overnight at 4°C using a mixture of 2% glutaraldehyde and 1.6% paraformaldehyde. Cells were washed three times in cacodylate buffer and were immobilized with 3% agarose, cut in a stick of 0.5 by 0.5 by 5 mm, washed twice in cacodylate buffer, and fixed in 1% OsO_4_ for 90 min at room temperature. Samples were dehydrated with grading ethanol (30%, 50%, 70%, 95%, 100%, 15 min each) and washed three times for 15 min with 100% ethanol. Samples were mixed in EPON-812 and hardener (BDMA) in a silicone mold and dried for 1 day at 37°C followed by 3 days at 60°C. Sections were cut with a Reichert-Jung Ultracut E Ultramicrotome using a 35° DiATOME diamond blade and then stained for 5 min in 2% uranyl acetate. To enhance uranyl acetate contrast, we proceed with a second stain for 3 min in 3% lead citrate. Grids were imaged on a JEM-2100plus TEM. Graph in Figure 2D represents the thickness of the inner layer in a total of 20 cells per condition and 100 measurements per cell using ImageJ [23]. Total chitin was quantified using epi-fluorescence microscopy. C. albicans cells were stained with calcofluor white (10 µg/ml) and images were taken using a Biotek ® Cytation 5 high-content microscope. For each condition, fluorescence intensities of 100 cells were measured using ImageJ [23].
Exo-glucanase and exo-chitinase assays
Cells were grown in SC+Mn or SC-Mn until reaching the exponential phase and washed with 50 mM sodium acetate pH 5.5. Exo-glucanase activity was assessed according to the method previously described by Gonzalez et al [24]. Cell pellets were incubated in 200 µl sodium acetate buffer containing 1% laminarin (w/v; Sigma, L9634-1 G) for 4 h at 37°C. Reduced glucose released after degradation of laminarin polymer by glucanase was quantified using DNS (Sigma, D0550-100 G) assay as previously described [25]. DNS mixed with reducing sugar releases colorimetric 3-amino-nitrosalicylic acid proportionally to reduced sugar concentration. Reduced sugar concentrations in samples were determined based on a glucose standard curve (0 - 4 mg/ml). Samples were mixed in a 1:20 (sample: DNS reagent (10 mg/ml)) ratio and heated for 5 min at 100°C to catalyze the reaction between sugar and DNS, and measurements were made at OD_540 nm_ using Cytation 5. Exo-chitinase activity was quantified as previously described by McCreath and Gooday [26]. Cell pellets were incubated in 200 µl sodium acetate buffer containing 1% 4-methylumbelliferyl N-acetyl-β-D-glucosaminide (4MU) (w/v; Sigma) for two hours at 37°C. Fluorescence was measured at λ_ex_ 360 nm and λ_em_ 440 nm.
Inductively coupled plasma-mass spectrometry
The total content of cell-associated calcium and manganese was quantified using inductively coupled plasma-mass spectrometry (ICP-MS) as previously described by Henry et al. [11].
Statistical analyses
GraphPad Prism 10 was used for statistical analyses. Data were generated from at least three independent biological replicates and then expressed as means ± standard deviation. Statistical difference between two sets of data with a non-parametric distribution was assessed using one-way ANOVA (Tukey’s multiple comparison test). The following p-values were considered: *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001.
Results
Transcriptomic analysis of smf12 under Mn starvation reflects a cell wall defect
In our previous work, we generated a mutant of the Mn NRAMP transporter, SMF12, to investigate the impact of Mn scarcity on the biology of C. albicans. We found that inactivation of SMF12 resulted in attenuated virulence, increased sensitivity to azoles, and activation of the unfolded protein response (UPR) [11]. To investigate further cellular processes affected in smf12, we carried out RNA-seq profiling under manganese (Mn) starvation. The transcriptional profile of smf12 growing in the Mn-free SC medium (SC-Mn) was compared to that of the WT strain growing under similar conditions. Upregulated transcripts in smf12 were mainly enriched in processes related to the synthesis and the processing of the three core polysaccharides of the fungal cell wall namely chitin (CHS2, 7, CDA2, CHT4), glucan (KRE1, 9, UTR2, PNG2) and phosphomannan (MNN1, 15, 41, 42, 43, 45, 47, BMT3, 4, 6, 7, KTR4, DFG5, FAV3) (Figure 1(A-B) and Supplementary Table S2). Transcripts related to cell wall signaling proteins and transcriptional control including the Cek1 MAP kinase, the histidine kinase Chk1 and the transcription factor Crz1 were also upregulated (Figure 1(B)). This transcriptional signature is reminiscent of a situation in which the cell wall is undergoing remodeling in response to a specific perturbation. Downregulated genes were mainly enriched in the acquisition of metals including iron (SIT1, CFL2, 4, 5, IRO1, FRE7, 30, FET31, CSA2, FTR1), zinc (ZRT101, PRA1, CSR1, ZRT2) and copper (CTR1) (Figure 1(C)). Genes of carbohydrate metabolism and both secreted lipases and proteases were differentially modulated in smf12 mutant suggesting a reprogramming of metabolism in response to the deletion of this Mn transporter (Figure 1(C)). When Mn was supplemented to the growth medium, the differential modulation of most of the smf12 transcripts described above was significantly attenuated (Figure 1(B-C)). This suggests that the transcriptional profile of smf12 is primarily a result of impaired Mn homeostasis in this mutant. Figure 1.smf12 exhibits a transcriptional signature reflecting cell wall perturbation.(A) GO enrichment of upregulated (red) and downregulated (blue) transcripts in smf12 mutant growing under Mn limitation. The transcriptional profile of smf12 growing in SC-Mn was compared to that of the WT strain (SN148-CIp20) growing under similar conditions. (B-c) Modulation of cell wall associated genes (B) and transcripts enriched in metals utilization and metabolism (C) in smf12 mutant. Heatmaps represent differentially modulated transcript in smf12 mutant under both Mn limitation (-Mn) and repletion (+Mn) as well as in WT in response to Mn starvation [11]. (D) Mn uptake in the smf11 mutant. Intracellular levels of Mn were determined by ICP-MS in the WT strain (SN148-CIp20) as well as in the smf11 and smf12 mutants. (E) smf mutants are sensitive to cell wall perturbers in an Mn-dependant manner. Spot assay was used to assess the impact of caspofungin (0.2 µg/ml) and calcofluor white (CFW; 25 µg/ml) on the growth of WT (SN148-CIp20) and smfs mutants (smf12, smf12 and smf12 smf11 double mutant). Strains were serially diluted, spotted on SC-Mn and SC + Mn, and incubated for 2 days at 30°C.
To test the potential cell wall perturbation in smf12, we assessed the sensitivity of this mutant as well as smf11, a mutant of Smf12 paralog Smf11, to various cell wall-perturbing agents, including caspofungin, which inhibits glucan synthesis, and calcofluor white (CFW), which binds and perturbs the chitin layers. Similar to the smf12 mutant, smf11 showed a marked defect in internalizing Mn, confirming its role in Mn uptake in C. albicans (Figure 1(D)). smf12 together with smf11 and their corresponding double mutant smf12 smf11 exhibited similar degree of hypersensitivity to CFW under Mn limitation as compared to their parental WT strain (Figure 1(E)). Supplementing the growth medium with Mn alleviated the CFW defect in all tested mutants, suggesting that the cell wall perturbation is a consequence of impaired Mn homeostasis. Intriguingly, in response to caspofungin, smfs mutants showed a mild growth defect exclusively under Mn sufficiency while Mn depletion rescued this phenotype (Figure 1(E)). Together, this underlined that Mn homeostasis mediated by Smf12 and Smf11 is required for cell wall integrity in C. albicans.
Inactivation of SMF12 alters cell wall ultrastructure and increases β-glucan and chitin exposures
The regulation of the exposure of cell wall polysaccharides, a process known as masking [27], is crucial for the outcome of an infection, as it enables fungal pathogens to evade host immune surveillance. To assess whether the observed cell wall perturbation in smf12 and smf11 affects cell wall masking, we quantified by flow cytometry the exposure of β-glucan, chitin, and mannans under both Mn deprivation and sufficiency conditions. While the exposure of the three polysaccharides in WT cells was not affected by Mn availability, smf12 and smf11 and the corresponding double mutant exhibited a drastic increase of both β-glucan and chitin exposures when Mn was depleted (Figure 2(A-B)). The unmasking of β-glucan and chitin in these mutants was reverted by Mn supplementation suggesting that Mn homeostasis is important for the cell wall masking in C. albicans. Figure 2.Altered cell wall ultrastructure and increased β-glucan and chitin exposures in smf mutants.(A) Quantification of β-glucan, mannan, and chitin exposure by flow cytometry in WT (SN148-CIp20) and smf mutants growing under either Mn scarcity or sufficiency. Cells were collected and incubated with FC-Dectin-1, WGA-Alexa-488, and Concanavalin A-Alexa-488 to assess the β-1,3-glucan, chitin, and cell wall mannan at the cell periphery, respectively. Median fluorescence intensity (MFI) was quantified by flow cytometry. (B) Representative confocal microscopy image showing β-glucan exposure in smf mutants and WT strains growing in the presence or the absence of Mn. (C-D) Effects of genetic inactivation of SMF12 and SMF11 on C. albicans cell wall architecture. TEM was used to analyze the effect of Mn availability on cell wall ultrastructure in WT (SN148-CIp20) and smf mutants (smf12 and smf11). Representative images are shown for each strain and condition (C). (D) Quantification of the thickness of the inner cell wall layer. Data represent the mean ± SEM of 100 measurements from 20 cells per condition. (E) smf mutants exhibit decreased Alcian blue binding. WT and smf mutants were stained with Alcian blue and the amount of dye bound to the cell wall was estimated by absorbance at OD_620 nm_. Data represent the mean ± SEM from three independent replicates.
The impact of smf mutations on the ultrastructure of the cell wall was also investigated using transmission electron microscopy (TEM). The thickness of the inner cell wall, which contains both glucan and chitin, was measured. While Mn availability has no significant effect on the inner cell wall thickness of the WT and smf11 cells, smf12 mutation led to a weak, but significant, decrease in the inner layer thickness under Mn limitation (Figure 2(C-D)). Furthermore, as the characteristic fibrillar outer layer of mannoproteins was not clearly observed by TEM, we employed the Alcian Blue binding assay to estimate the thickness of the cell wall phosphomannan. Under Mn sufficiency, both WT and smf mutants showed comparable high levels of staining (Figure 2(E)). However, upon Mn depletion, smf mutants exhibited a drastic reduction in staining intensity as compared to the WT suggesting a significant reduction of the mannan fibrillar outer layer which might explain the unmasking of β-glucan and chitin phenotype. This supports the previous finding highlighting the crucial role of Mn homeostasis in protein mannosylation and glycosylation in C. albicans [11,13]. Together, these findings support that Mn uptake deficiency led to cell wall structure perturbation and unmasking of both chitin and β-glucan.
Cell wall unmasking in smf12 and smf11 under Mn scarcity affects host-pathogen interaction
Different studies have highlighted the impact of glucan and chitin exposure on the phagocytosis rate of immune cells. In general, unmasking either glucan or chitin in certain mutants or in response to specific environmental cues has been shown to enhance immune recognition and increase phagocytosis by macrophages and neutrophils [27]. Based on this, we investigated whether the increased exposure of glucan and chitin on the surface of smf12 and smf11 cells led to enhanced phagocytosis by macrophages. J774A.1 murine macrophages were exposed to either WT or smfs mutant cells grown in an Mn-depleted or repleted medium. C. albicans WT strain was phagocytosed at the same rate regardless of Mn availability in the growth medium (Figure 3(A)). A significant decrease of phagocytosis rate was perceived when smf12, smf11, or smf11smf12 mutants were grown under Mn limitation (Figure 3(A)). This phagocytosis pattern was reversed when Mn was supplemented to a level comparable to that of the WT strain. This suggests that the unmasking phenotype of smf mutants under Mn starvation is linked to a reduced recognition or internalization by macrophages. Figure 3.Phagocytosis of smf mutants by murine macrophages and the subsequent inflammatory response.(A) smf mutants and WT (SN148-CIp20) cells pre-grown in an Mn-depleted or -repleted medium were co-incubated with J774A.1 macrophages at an MOI of 5:1 for one hour, and the rate of phagocytosis determined by flow cytometry. For each strain and each growth condition, data represent the percentage of phagocytosis as compared to the WT in SC+Mn. (B-C) Altered smf mutants’ morphology is reverted by Mn supplementation. (B) Representative images of WT (SN148-CIp20) and smf mutants’ morphologies showing cells stained with CFW. (C) Distributions of the number of yeast cells per cluster for each strain under Mn limitation or repeltion. (D-E) Inflammatory responses in macrophage incubated with smf mutants. ELISA results are shown for TNF-α and IL-6 in supernatants of co-culture media.
The reduced phagocytosis rate in smf mutants under Mn starvation may be linked to their morphological defects. Most of these cells grew as larger multicellular aggregates, which could enhance resistance to phagocytosis, as previously reported for different mutants with similar morphological defect [13,28–32]. Indeed, Mn depletion induced a clumping morphology in smf mutants, likely resulting from a cell separation defect, which was rescued by Mn supplementation (Figure 3(B-C)). Alternatively, the decreased phagocytosis could result from the overexposure of chitin in smf mutants, potentially blocking Dectin-1. Previous studies have shown that chitin interferes with Dectin-1 receptor activity, inhibiting phagocytosis and leading to reduced cytokine production and macrophage engulfment [33–35]. Consistent with this, the overexposure of either glucan or chitin on the surface of smf mutants under Mn limitation did not significantly affect the immune response, as assessed by the levels of tumor necrosis factor alpha (TNF-α) and IL-6, secreted in media, compared to Mn-repletion conditions (Figure 3(D-E)). Taken together, these findings suggest that although smf mutants exhibit increased exposure of glucan and chitin, their reduced susceptibility to phagocytosis is likely due to their altered morphology and/or the masking of Dectin-1.
smf12 unmasking is not mediated by the calcineurin pathway
Gene Set Enrichment Analysis (GSEA) was used to identify similarities between the smf12 transcriptional profile and those of potential signaling pathways that could mediate the observed unmasking phenotype. We found that smf12 profile significantly correlates with different transcriptomics datasets of mutants of the calcineurin pathway, including cna1 mutant of the calcineurin catalytic subunit and its major transcription factor effector, crz1 (Figure 4(A-B) and Supplementary Table S3). A total of 36 transcripts that are activated by calcium and regulated by both calcineurin and Crz1 (60 in total) [37] were upregulated in smf12 (Figure 4(C)). This set of common genes were enriched mainly in biological processes linked to cell wall remodeling, calcineurin signaling (CRZ1, CCH1, RTA2) stress responses and transport (Figure 4(D)). Again, supplementing the growth medium with Mn attenuated the calcineurin activation signature of smf12 mutant suggesting a causality between Mn uptake defect and calcineurin signaling activation. Under Mn limitation, smf12, as well as smf11 and smf11smf12 mutants, exhibited elevated calcium levels compared to the WT cells, corroborating the activation of the calcineurin transcriptional output in smf12 (Figure 4E). Figure 4.Activation of the calcineurin pathway in smf12.(A) GSEA analysis indicates a transcriptional signature reflecting calcineurin pathway activation in smf12 mutant. The diameter of the circle reflects the number of modulated transcripts in each dataset. Images were generated using Cytoscape [36] with the Enrichment Map plug-in. (B) GSEA graphs of significant correlations between smf12 transcriptome and mutants of the calcineurin pathway (cna1 and crz1). (C-D) Overlap of significantly upregulated transcripts in smf12 mutants with the set of genes defining the calcineurin regulon in C. albicans [37]. (E) smf mutants exhibit high intracellular calcium (Ca) content. Ca levels were determined by ICP-MS in exponentially grown WT (SN148-CIp20) and smf strains in SC-Mn medium at 30°C. (F) smf12 unmasking of β-glucan and chitin is not mediated by the calcineurin pathway. Quantification of β-glucan, mannan, and chitin exposure by flow cytometry in WT and smf12 growing under either Mn scarcity or sufficiency in the presence or the absence of the calcineurin inhibitor, FK506.
Former study underlined the contribution of the calcineurin signaling to the unmasking phenotype of C. albicans [38,39]. As our RNA-seq data reflect an activation of this signaling pathway, we hypothesized that smf12 unmasking is mediated by the calcineurin pathway. To test this, we assessed both glucan and chitin exposures of smf12 mutant in the presence of the calcineurin inhibitor FK506. Under Mn limitation, as described above, smf12 exhibited an unmasking of both β-glucan and chitin (Figure 4F). However, inhibition of the calcineurin signaling in this mutant increased further the exposition of both β-glucan and chitin as compared to the control (Figure 4F). Mn repletion reverted the unmasking of smf12 in the presence or the absence of the calcineurin inhibitor (Figure 4F). Thus, activation of the calcineurin signaling under Mn starvation is unlikely modulating the unmasking phenotype of smf12.
Smf mutants exhibit a decreased Mn-dependant exo-glucanase activity
Glucanases such as the endo-glucanase Eng1 and the exo-glucanase Xog1 were shown to mediate shaving of exposed β-glucan and contribute to the hiding of fungal pathogen from the immune system recognition [40–42]. As our RNA-seq showed a decreased transcript level of the endo-glucanase Eng1, we hypothesized that overexposure of β-glucan on the surface of smf12 might be a consequence of a decreased shaving capacity of this epitope. We measured β-glucanase activity of intact cells of smf mutants using laminarin as a substrate. All smf mutants exhibited a significant decrease in glucanase activity under Mn limitation as compared to the WT strain and the Mn-repletion conditions, suggesting a reduction of the β-glucan “shaving” capacity of smf mutants (Figure 5A). Figure 5.Smf mutants exhibit a decreased glucanase activity and a high chitin content.(A) Exo-glucanase activity of WT (SN148-CIp20) and smf (smf12, smf11, and smf12 smf11) cells was determined in each strain under either Mn limitation or repletion. (B) Chitinase activity of WT (SN148-CIp20) and smf mutants in response to Mn availability. (C) Total chitin assessment in WT (SN148-CIp20) and smf mutant strains. Total chitin was quantified using microscopy and CFW staining. For each condition, fluorescence intensities of 100 cells were measured using ImageJ.
Chitin synthesis, but not processing, is altered in smf12
Previous work have shown that chitin unmasking in response to low pH resulted from a reduced expression of the cell wall chitinase Cht2 leading to decreased processing of chitin fibrils [43]. As our RNA-seq data showed a downregulation of CHT2 and also CHT3, we hypothesized that a similar mechanism causes de-cloaking of chitin at the cell periphery of smf12 under Mn limitation. When comparing chitinase activity of the WT and smf mutants under either Mn scarcity or repletion no significant difference was noticed (Figure 5B). Furthermore, other studies highlighted the role of increased chitin biosynthesis and deposition at the unmasked foci of the cell wall in altering its architecture, which consequently leads to the overexposure of glucan [39,44]. Indeed, smf12 exhibited elevated transcript levels of the chitin synthase CHS2 and an increased cellular chitin content, particularly under Mn limitation (Figure 5C). A similar increase in chitin content was observed in smf11, and to a lesser extent, smf11smf12 double mutant. However, smf11smf12 was not able to recover the WT chitin content as did smf11 and smf12 when Mn was supplemented (Figure 5C). Collectively, these findings indicate that the chitin unmasking observed in smf12 is likely due to enhanced chitin biosynthesis and deposition at the cell wall, rather than a defect in chitin cleavage.
Discussion
Recent studies have highlighted the critical role of Mn homeostasis in regulating various biological processes that influence the fitness and virulence of C. albicans [10–12]. Mn has been shown to play an important role in C. albicans’ ability to cope with both endoplasmic reticulum and plasma membrane stress, and it is essential for the functionality of superoxide dismutase enzymes. In this study, we uncovered a novel function of Mn in maintaining cell wall integrity and modulating the exposure of fungal antigenic determinants, further emphasizing the central role of this metal in supporting the opportunistic nature of this yeast.
The sensitivity of smf mutants to CFW underscores the critical role of Mn metabolism in maintaining the integrity of the C. albicans cell wall. This defect is reflected in the transcriptome of smf12, with differential regulation of many genes involved in cell wall biogenesis, as well as transcriptional signatures indicative of the activation of cell wall integrity (CWI) pathways, including calcineurin and the MAPK Cek1. The cell wall defect observed in smf12 could be primarily attributed to alterations in the abundance of chitin on the cell surface. Notably, high chitin content in C. albicans and other Candida species has previously been linked to hypersensitivity to CFW as shown here for smf mutants [45]. Conversely, increased chitin content has been associated with enhanced tolerance to caspofungin [45–47]. This might explain why smf mutants exhibit sensitivity to this echinocandin only when Mn is available, whereas under Mn-limited conditions, the elevated chitin content may help mitigate their sensitivity to caspofungin. Furthermore, the mannosylation defect observed in smf mutants, as previously reported [10,11], together with the reduced thickness of the internal cell wall layer reported here, could lead to increased chitin levels, similar to what was seen in mannosyltransferase mutants in C. albicans [48].
In this study, we demonstrate that impairing C. albicans cells’ ability to uptake Mn results in the unmasking of both chitin and β-glucan. This phenotype is likely the result of a combination of various alterations in the smf mutants. First, the Alcian blue binding assay indicated a reduction of the mannan fibrillar outer layer which might primarily contribute to the unhiding of β-glucan from the cell surface as previously observed in C. albicans mannosylation- and mannoprotein-deficient mutants [49–51]. Second, increased chitin content and deposition at the cell surface may drive β-glucan unmasking in smf12. Indeed, β-glucan unmasking in C. albicans, whether triggered by antifungal drugs, environmental or host signals, is typically accompanied by increased chitin deposition at the cell periphery [27]. Thus, abnormal deposition of chitin layer at the cell wall of smf12 might push β-glucan to emerge at the cell periphery. Third, while chitin processing by chitinases at the surface of the cell wall was unaffected in smf mutants, glucanase activity was significantly impaired, leading to a reduced capacity for β-glucan “shaving” in these mutants. Notably, the transcript levels of the endo-glucanase Eng1 were reduced in smf12, suggesting that the unmasking of β-glucan could result from improper modulation of these enzymes and an inability to cleave exposed β-glucan from the cell surface [40]. Overall, it is likely that one or more of these mechanisms, or a combination thereof, explain the unmasking of β-glucan and chitin in smf mutants.
In C. albicans, β-glucan unmasking is regulated by multiple signaling mechanisms including the calcineurin and the Cek1 MAPK pathways [27,38]. In the current study, although calcineurin was activated in smf12, as evidenced by the activation of its bona fide regulation, inhibition of this signaling pathway failed to restore β-glucan and chitin exposure to wild-type levels. This suggests that the unmasking phenotype in smf12 is independent of the calcineurin pathway. Our RNA-seq data revealed an upregulation of the MAPK Cek1, indicating a potential role for this signaling pathway in mediating the unmasking phenotype observed in smf12. However, GSEA analysis did not reveal a significant similarity between the smf12 profile and mutants of the Cek1 pathway (e.g. mutant of the transcription factor effector, Cph1 and STE11^ΔN467^ hyperactive mutant). Therefore, further investigation is required to determine whether Cek1 or another, yet unidentified, pathway is mediating the unmasking phenotype under Mn deficiency. Additionally, the cause of calcineurin hyperactivation in smf12 remains to be elucidated. In many fungi, the calcineurin pathway is a well-known regulator of both cell wall integrity and UPR [52–54]. Accordingly, one possible explanation is that the calcineurin pathway could be activated as a secondary response to signal cell wall alterations and/or ER stress, given that the UPR is constitutively activated in smf12 [11,55]. Furthermore, calcineurin activation might prevent the formation of non-viable supra-high chitin smf12 cells by modulating chitin biosynthesis as previously reported [56].
In the current study, we found that the unmasking of either chitin or glucan did not lead to an enhanced immune response or increased phagocytosis. The reduced susceptibility of smf mutants to phagocytosis is likely due to their altered morphology, as they formed aggregates, presumably resulting from a defect in glucan processing at the septa [57]. Similar results have been observed in C. albicans phosphomannan-deficient mutants and other mutants of glycosyltransferases, which also exhibit an aggregate phenotype akin to that of smf12 and show resistance to phagocytosis [29,31]. Recent studies on the multi-drug-resistant yeast C. auris have demonstrated that evolved aggregative cells, or cells where aggregation is induced by specific drugs or growth conditions, are more resistant to phagocytosis by macrophages than their yeast-form counterparts [28,58]. Future research will be essential to further clarify the role of Mn homeostasis in the development of this multicellular morphology and to investigate whether Mn-starved niches within the human host may contribute to the immunity-recalcitrant phenotype observed in C. auris. The decreased uptake of smf mutants by macrophages may be a consequence of Dectin-1 masking, likely due to the high chitin content on their cell surface. Interestingly, this fungal survival strategy has been recently described in C. albicans, where increased chitin on the cell surface promotes arginine degradation by host arginase-1, thereby preventing the utilization of this amino acid by nitric oxide synthase to produce the antifungal molecule nitric oxide [33].
In the present work, with few exceptions, the deletion of either SMF12, SMF11, or both resulted in a similar phenotype. This observation suggests that Smf11 and Smf12 are likely paralogs that work together as a heterodimer complex to mediate Mn uptake. Although this has not yet been investigated for NRAMP transporters, previous studies have demonstrated that paralogs of ABC transporters can form heterodimers in both bacteria and higher eukaryotes [59,60]. Notably, the heterodimeric configuration of these transporters has been shown to alter the substrate specificity of the interacting partners [61]. Thus, future research is necessary to determine whether Smf11 and Smf12 can form heterodimers and, if so, whether this affects their substrate specificity, particularly for metals. A recent investigation showed that Smf11 transports ferrous iron in C. albicans and contributes to Fe sensing [62]. Similarly, in S. cerevisiae, the Smf11 ortholog, Smf1, has been shown to transport Fe as well as other cations such as potassium and calcium [63]. Therefore, it is likely that inactivating Smf11 or Smf12 in C. albicans may disrupt the homeostasis of other metals or cations, potentially triggering compensatory uptake mechanisms. This could explain why, in the current study, both smf11 and smf12 mutants exhibited elevated calcium levels [36], likely due to the activation of alternative Ca^2 +^ uptake systems [13].
Supplementary Material
Table S2.xlsx
Table S1.xlsx
Table S3.xlsx
Table S4.xlsx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Murdoch CC, Skaar EP. Nutritional immunity: the battle for nutrient metals at the host–pathogen interface. Nat Rev Microbiol. 2022;20(11):657–15. doi: 10.1038/s 41579-022-00745-635641670 PMC 9153222 · doi ↗ · pubmed ↗
- 2Pijuan J, Moreno DF, Yahya G, et al. Regulatory and pathogenic mechanisms in response to iron deficiency and excess in fungi. Microb Biotechnol [Internet]. 2023 [cited 2024 Dec 2];16(11):2053–2071. doi: 10.1111/1751-7915.1434637804207 PMC 10616654 · doi ↗ · pubmed ↗
- 3Alselami A, Drummond RA. How metals fuel fungal virulence, yet promote anti-fungal immunity. Disease Model Mechanisms [internet]. 2023 [cited 2024 Dec 4];16(10):dmm 050393. doi: 10.1242/dmm.050393 PMC 1062967237905492 · doi ↗ · pubmed ↗
- 4Blatzer M, Latgé J-P. Metal-homeostasis in the pathobiology of the opportunistic human fungal pathogen Aspergillus fumigatus. Curr Opin Microbiol [Internet] 2017 [cited 2024 Dec 4];40:152–159. Available from: https://www.sciencedirect.com/science/article/pii/S 136952741730214 X 29179120 10.1016/j.mib.2017.11.015 · doi ↗ · pubmed ↗
- 5Kornitzer D, Roy U. Pathways of heme utilization in fungi. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research [Internet] Biochim et Biophys Acta (BBA) - Mol Cell Res. 2020 [cited 2021 Jul 12];1867(11):118817. Available from: https://www.sciencedirect.com/science/article/pii/S 016748892030175010.1016/j.bbamcr.2020.11881732777371 · doi ↗ · pubmed ↗
- 6Volkova M, Atamas A, Tsarenko A, et al. Cation transporters of Candida albicans—new targets to fight candidiasis? Biomolecules. 2021;11(4):11. doi: 10.3390/biom 11040584 PMC 807335933923411 · doi ↗ · pubmed ↗
- 7Alves R, Barata-Antunes C, Casal M, et al. Adapting to survive: how Candida overcomes host-imposed constraints during human colonization. PLOS Pathog [Internet]. 2020 [cited 2024 Dec 2];16(5):e 1008478. doi: 10.1371/journal.ppat.100847832437438 PMC 7241708 · doi ↗ · pubmed ↗
- 8Khemiri I, Tebbji F, Sellam A. Transcriptome analysis uncovers a link between copper metabolism, and both fungal fitness and antifungal sensitivity in the opportunistic yeast Candida albicans. Front Microbiol. 2020;11:935. doi: 10.3389/fmicb.2020.0093532508775 PMC 7248230 · doi ↗ · pubmed ↗