Acquired tick resistance in Peromyscus leucopus alters Ixodes scapularis infection
Elis A. Fisk, Cassie J. Leonard, Kristin L. Rosche, Elisabeth Ramirez-Zepp, Jeffrey R. Abbott, Jeb P. Owen, Dana K. Shaw

TL;DR
Mice that develop resistance to ticks reduce tick feeding success and pathogen spread.
Contribution
The discovery of acquired tick resistance in Peromyscus leucopus and its impact on tick-borne pathogen transmission.
Findings
Repeated tick infestations in mice lead to immune-mediated rejection of ticks.
Ticks feeding on resistant mice ingest more host immune cells and pathogens.
Acquired resistance could reduce tick populations and pathogen circulation.
Abstract
Ticks are obligate hematophagous parasites and pathogen vectors responsible for morbidity and mortality worldwide. Ixodes scapularis is a vector for at least seven pathogens relevant to human and animal health, including the Lyme disease microbe, Borrelia burgdorferi, and the causative agent of anaplasmosis, Anaplasma phagocytophilum. Tick-host interactions affect the maintenance of tick-borne pathogens in a population. Here, we report that repeated I. scapularis larval infestations on the wild host species Peromyscus leucopus lead to immune-mediated rejection of the tick, a phenomenon termed acquired tick resistance (ATR). On previously infested mice, we observed that larval feeding success was reduced by over 50%, and fed larvae had decreased blood meal weights compared to larvae fed on naïve hosts. Over sequential infestations, mice exhibited increasingly severe inflammation at tick…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
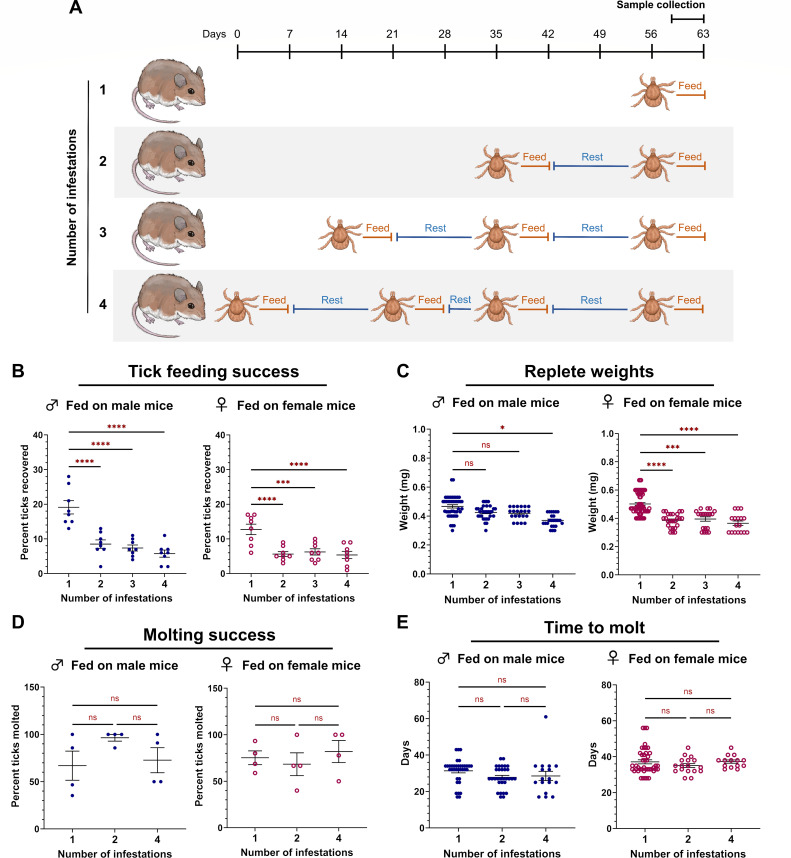 Fig 1
Fig 1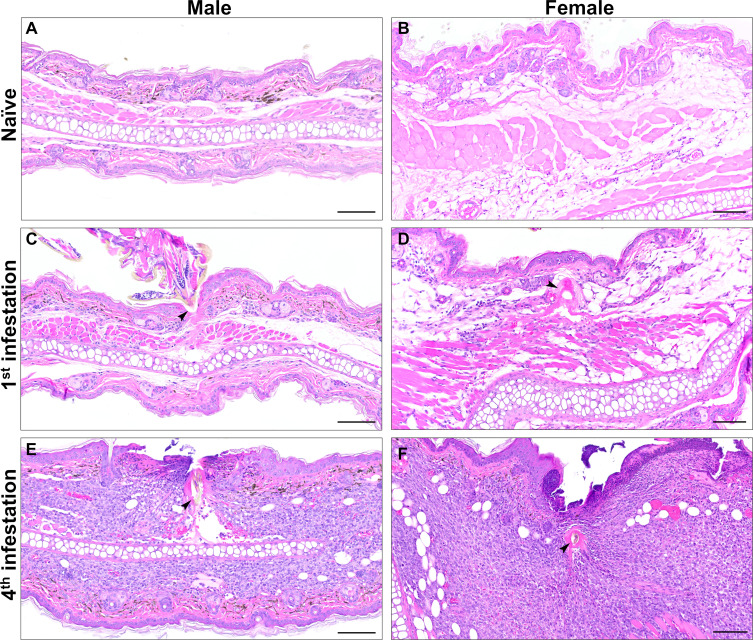 Fig 2
Fig 2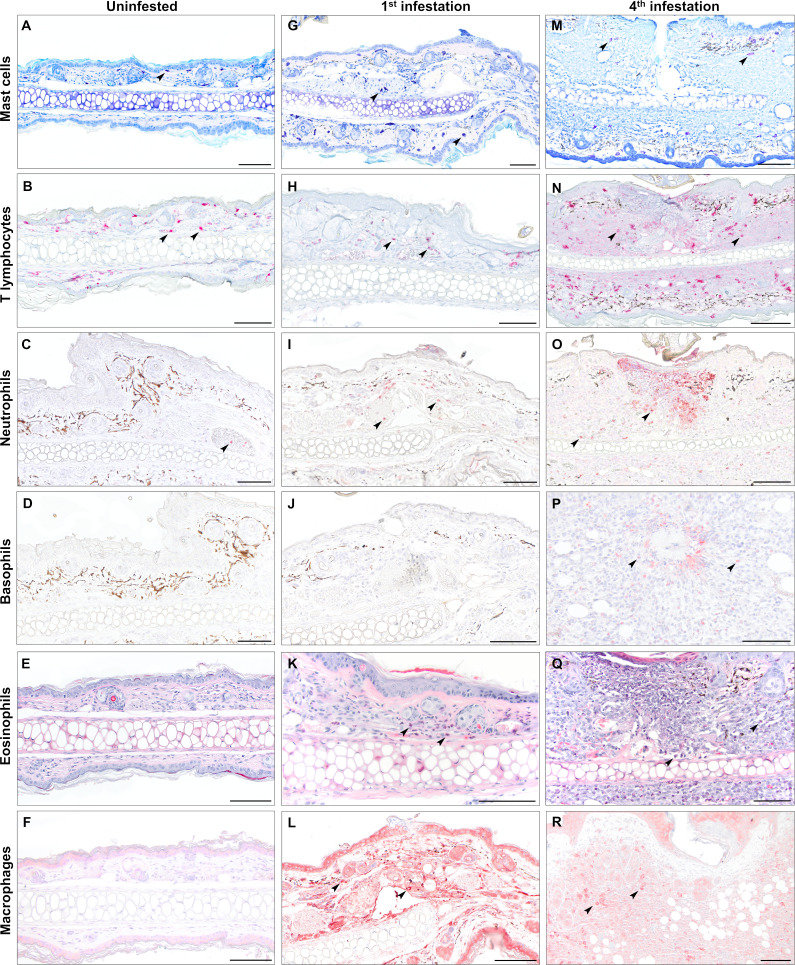 Fig 3
Fig 3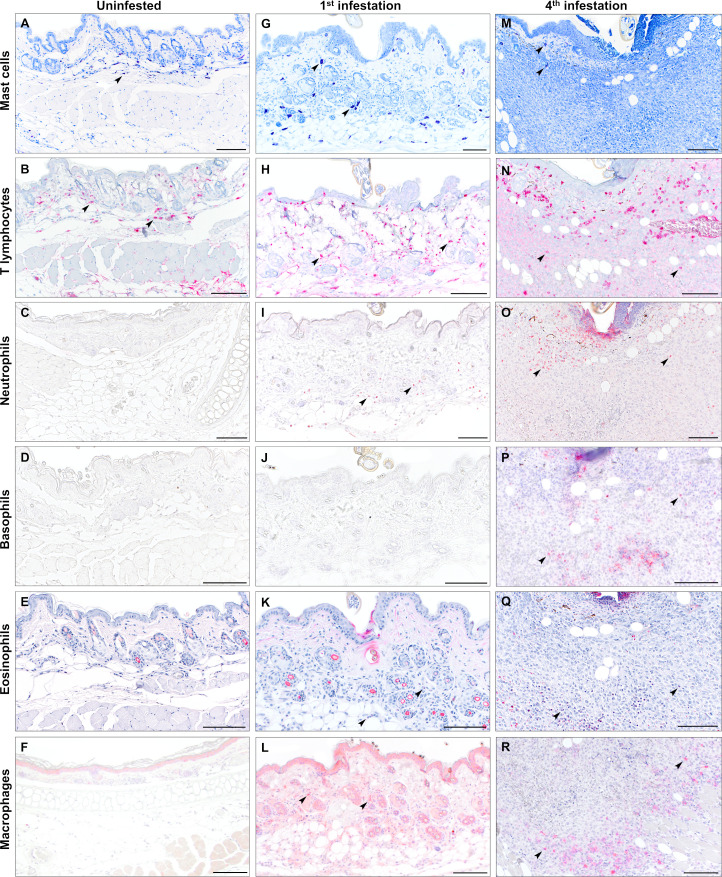 Fig 4
Fig 4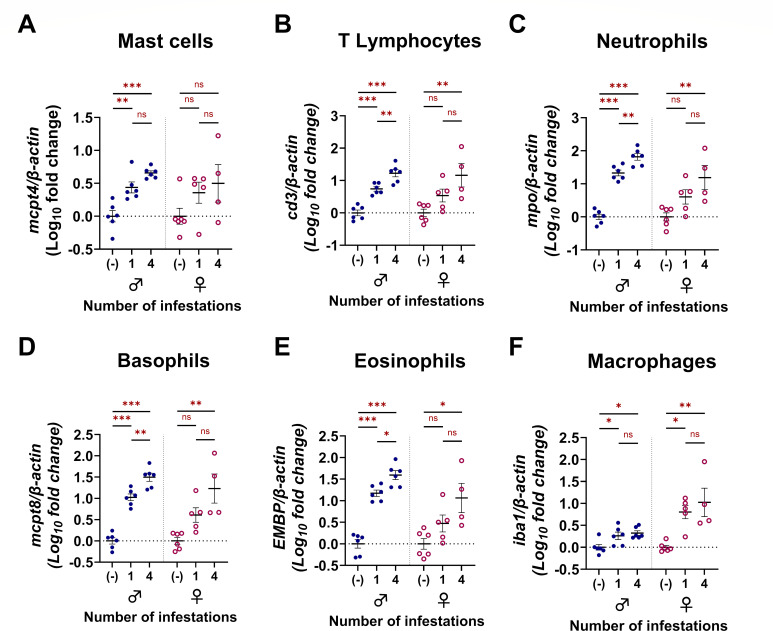 Fig 5
Fig 5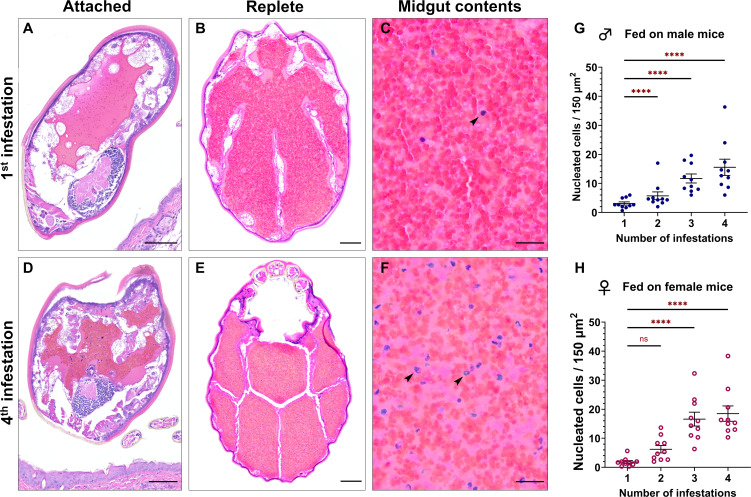 Fig 6
Fig 6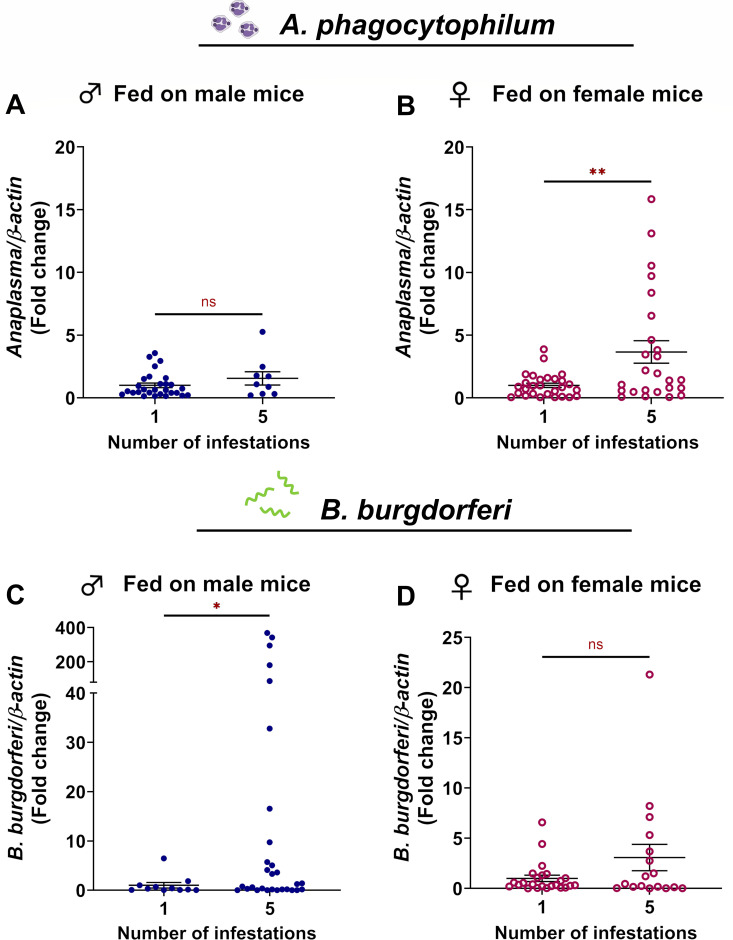 Fig 7
Fig 7- —National Institute of Biomedical Imaging and Bioengineeringhttp://dx.doi.org/10.13039/100000070
- —Division of Intramural Research, National Institute of Allergy and Infectious Diseases
- —Achievement Rewards for College Scientists Foundationhttp://dx.doi.org/10.13039/100008227
- —G. Caroline Engle Distinguished Professor in Infectious Diseases Award
- —Washington State University, College of Veterinary Medicine
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Viral Infections and Vectors · Mosquito-borne diseases and control
INTRODUCTION
The incidence of tick-borne disease in the United States has been on the rise over the last decade, with 50,865 cases reported in 2019 increasing to 73,384 cases in 2022 (1). However, passive surveillance may underestimate the true clinical burden, as insurance claims data estimate that 476,000 patients annually were diagnosed and treated for Lyme disease alone (2). Ixodes scapularis, more commonly known as the deer tick, is capable of transmitting at least seven pathogens relevant to human health (3), including Borrelia burgdorferi, the causative agent of Lyme disease, and Anaplasma phagocytophilum, the causative agent of anaplasmosis. How the host and arthropod vector interact is a driving force influencing the maintenance of tick-borne pathogens in natural systems.
Some host species will develop protective immunity against ticks after repeated infestations, a phenomenon termed “acquired tick resistance” (ATR) (4). Tick salivary antigens elicit a T helper 2 (Th2) response (5), which stimulates IgE and/or IgG antibody production by B cells (6, 7). This IgE enters the bloodstream and arms the host’s circulating basophils and/or mast cells against tick antigens (7–11). Primed CD4 + memory T cells within the draining lymph nodes will then migrate to the skin. Upon subsequent tick bites, they secrete interleukin 3 (IL-3), which recruits basophils from the vasculature into the bite site (12). The recruited basophils, along with resident mast cells, will release histamine and other mediators that trigger local edema, itching, and epidermal hyperplasia (13–20). This type of immune response detrimentally impacts ticks by thwarting attachment success and reducing feeding weights, molting success, fecundity, and survival (4, 21–25).
Tick resistance has been well studied in laboratory animals, such as guinea pigs (Cavia porcellus), which develop ATR responses that reject over 80% of feeding ticks (21, 26, 27). For this reason, there is significant interest in developing translational strategies that will block tick feeding and pathogen transmission to humans and/or animals. In contrast, interactions with wild animal species are less understood. The white-footed mouse, Peromyscus leucopus, is considered the reservoir host for B. burgdorferi and A. phagocytophilum and is often assumed to not mount ATR responses that reject Ixodes ticks. However, some studies have reported varying degrees of resistance that appear to depend on life stage of the tick (28–31). While repeated nymphal infestations cause an increasing severe inflammatory response in P. leucopus, it does not hamper tick feeding (31). In contrast, repeated larval infestations result in lower feeding success, decreased weight, and reduced fecundity (28–30). Since nymph and adult life stages are primarily responsible for transmitting disease-causing microbes (32–34), these have been the main focus of ATR research (6, 35, 36). Larvae do not transmit B. burgdorferi and A. phagocytophilum (37, 38), but are the first life stage to become infected, which makes them essential for pathogen maintenance in the population (32). The type of immunity elicited by P. leucopus in response to repeated larval infestation, and whether this impacts pathogen acquisition, is not known.
In this study, we demonstrate that white-footed mice mount ATR responses against I. scapularis larvae, which impacts tick feeding and pathogen ingestion. We found that previously infested hosts exhibited severe inflammation at sites of larval attachment predominated by basophils, eosinophils, and neutrophils with epidermal ulceration and hyperplasia. In contrast, tick-naïve mice exhibited only mild to moderate inflammation predominated by macrophages, eosinophils, and neutrophils. The severe inflammation observed in sensitized mice correlated with a significant reduction in larval feeding success, fewer larvae reaching repletion, and decreased blood meal volumes. Additionally, the quantity of leukocytes ingested by feeding larvae increased proportionately with the number of prior infestations experienced by the host. We found that larvae ingested a greater number of B. burgdorferi from sensitized male mice, whereas A. phagocytophilum ingestion was enhanced with sensitized female mice. Altogether, we demonstrate that wild host species develop ATR against larval I. scapularis, which hampers tick feeding and alters pathogen acquisition.
RESULTS
Larvae feed less successfully on tick-sensitized P. leucopus
Both larval and nymph life stages are important for the cycle and maintenance of pathogens in a wild population. However, little is known about wild host responses to larval infestation. We therefore sought to quantify how previous tick exposure in P. leucopus influenced larval feeding success. P. leucopus mice were infested with 100 I. scapularis larvae one to four times (Fig. 1A), and ticks were allowed to feed to repletion over seven days. For mice infested more than once, a two-week waiting period was observed between infestations to allow an adaptive immune response to develop. After each infestation, the proportion of larvae feeding to repletion and the replete larval weight were quantified. During primary infestations, an average of 19.1% (17.2%–21.0%) of larvae successfully fed on male mice, whereas only 5.8% (4.7%–6.8%) fed on sensitized mice, representing a 70% reduction in feeding success (Fig. 1B). Similar decreases in feeding success were observed in larvae fed on female mice, with up to 58% fewer larvae reaching repletion on sensitized mice (statistics summarized in Tables S1 and S2). We also noted that larvae fed more successfully on naïve males compared to naïve females. However, following at least one tick sensitization, both sexes supported similar numbers of larvae.
*Reduced I. scapularis larvae feeding success on previously sensitized P. leucopus. (A) Schematic depicting the infestation schedule. Each larva is representative of one infestation event in which 100 larvae were manually placed on an anesthetized mouse. Larvae were allowed seven days to feed to repletion. Animals used for histopathology and leukocyte analysis followed a similar infestation schedule with the final sample collection occurring 72 hours post-attachment. (B) Percent feeding success, (C) repletion weights, (D) molting success, and (E) time to molt were quantified for all conditions. Each data point represents either the proportion of larvae feeding to repletion on a single animal (B, D) or a single replete larva (C, E). A negative binomial generalized linear mixed-effects model was used for statistical analysis of larval feeding success and time taken to molt. Larval replete weight was analyzed using a linear mixed-effects regression model. Molting success was analyzed using a generalized linear mixed-effects model. **P < 0.01; ***P < 0.001, ***P < 0.0001. ns, not significant.
To quantify blood meal volume, replete larvae were weighed. Larvae fed on naïve female mice had significantly larger blood meal volumes (0.502 mg ± 0.009) compared to those fed on sensitized mice (Fig. 1C). Larvae fed on mice during the fourth infestation ingested 0.365 mg (± 0.016), representing a 27% reduction in blood meal volume. For larvae fed on male mice, an 18% drop in blood meal volume was observed between the first infestation (0.456 mg ± 0.010) and fourth infestation (0.376 ± 0.009). Although ticks fed on naïve mice had larger bloodmeals, there were no differences in the proportion of larvae molting successfully (Fig. 1D) or the time taken to molt (Fig. 1E). Bloodmeal volumes and molting success were also comparable between ticks fed on either male or female mice. Taken together, these data demonstrate that I. scapularis larvae feed less successfully on sensitized male and female P. leucopus and acquire less blood. Our findings are consistent with previous reports describing fewer larvae reaching repletion (29) and decreased replete weight (28) in larvae fed on sensitized P. leucopus.
Tick sensitization is associated with severe inflammation at the larval attachment site
We next histologically characterized the host’s skin response to larval attachment. P. leucopus mice were infested with larvae one to four times as shown in the experimental timeline (Fig. 1A). During the final round of infestations, mice were euthanized at 72 hours, and punch biopsies were taken at larval attachment sites. Histologic findings from naïve mouse skin and from larval attachment sites are summarized in Table S3. When compared to skin biopsies from uninfested P. leucopus (Fig. 2A and B; Fig S1A and B), mice infested once showed a mild focal inflammatory response at the bite site centered around the larval hypostome/cement cone (arrow) (Fig. 2C and D; Fig S1C and D). Inflammation was composed of only a small number of inflammatory cells which aggregated within the dermis and did not extend significantly into the underlying subcutis or skeletal muscle. Although the epidermis adjacent to larval attachment sites was mildly hyperplastic (25.2 µm ± 2.0 for males, 36.8 ± 6.5 for females), epidermal thickness did not differ significantly from uninfested mice (12.4 µm ± 1.2 for males, 17.2 ± 0.7 for females). In contrast, mice infested four times showed a moderate to severe inflammatory response at the bite site centered around the larval hypostome (arrow) (Fig. 2E and F; Fig S1E and F). Moderate to large numbers of inflammatory cells infiltrated the dermis with extension into the underlying subcutis and skeletal muscle. In some biopsies, inflammation crossed the boundary of the auricular cartilage and was present within the connective tissue of both the concave and convex pinna. The epidermis abutting larval mouthparts was eroded or ulcerated with serocellular crusting, and the intact epidermis at the bite site exhibited significant hyperplasia (57.7 µm ± 15.7 for males, 88.0 ± 15.2 for females) compared to uninfested skin (Fig. 2E and F; Fig. S2; Tables S1 to S3). These findings demonstrate that the inflammatory response to larval attachment is more severe in P. leucopus that were previously sensitized.
Increasing inflammation at larval attachment sites with previous tick exposure. Histological examination of male and female mouse biopsies from (A and B) naïve pinna and (C–F) larval attachment sites from (C and D) a primary infestation and (E and F) a quaternary infestation. (C–F) Lymphocyte, macrophage, and eosinophil infiltrates in the dermis are centered around the hypostome and cement cone. The larval hypostome (C, E and F) and cement cone (D) are indicated by arrows. (E and F) The epidermis adjacent to the larval attachment site is ulcerated with serocellular crusting. H&E stain. Bar = 100 µm.
Larval bite sites reveal an influx of inflammatory cells with repeated exposure
We next sought to characterize the immune cell population mediating the inflammatory response at larval bite sites. Skin samples were taken from male (Fig. 3; Fig. S3) and female (Fig. 4; Fig. S4) mice infested either one or four times for immunohistochemistry and differential staining. Mast cells and eosinophils were visualized with toluidine blue and Luna stains, respectively. Neutrophils (MPO), basophils (MCPT8), T lymphocytes (CD3), and macrophages (IBA1) were visualized by immunohistochemistry with antibodies against specific cell type markers. We found that skin from uninfested male mice had small numbers of resident mast cells (Fig. 3A) and T lymphocytes (Fig. 3B). Few neutrophils were present but confined to the vasculature (Fig. 3C). Naïve skin lacked basophils (Fig. 3D), eosinophils (Fig. 3E), and macrophages (Fig. 3F). During the primary larval infestation in male mice, small numbers of neutrophils (Fig. 3I), eosinophils (Fig. 3K), and macrophages (Fig. 3L) infiltrated the dermis around the larval bite site. In contrast, male mice infested four times showed a robust inflammatory infiltrate composed of T lymphocytes (Fig. 3N), neutrophils (Fig. 3I), basophils (Fig. 3P), eosinophils (Fig. 3Q), and a moderate number of macrophages (Fig. 3R). Sensitized mice showed neutrophils and basophils clustered around and extending into the cement cone of larval ticks (Fig. 3O and P). Similar changes were present in biopsies from female mice (Fig. 4). Naïve skin from female mice contained small numbers of resident mast cells (Fig. 4A) and T lymphocytes (Fig. 4B), with an absence of neutrophils (Fig. 4C), basophils (Fig. 4D), eosinophils (Fig. 4E), and macrophages (Fig. 4F). The inflammatory response to primary infestations was characterized by small numbers of neutrophils (Fig. 4I), eosinophils (Fig. 4K), and macrophages (Fig. 4L). Female mice infested four times developed marked inflammatory infiltrates composed of numerous T lymphocytes (Fig. 4N), neutrophils (Fig. 4O), basophils (Fig. 4P), eosinophils (Fig. 4Q), and macrophages (Fig. 4R).
Serial infestation of male P. leucopus with larval I. scapularis elicits eosinophil, neutrophil, basophil, T lymphocyte, and macrophage infiltrates. Leukocyte characterization in naïve skin and at primary and quaternary larval attachment sites in male mice using special stains and immunohistochemistry. (A, G, M) Mast cells are visualized with toluidine blue stain (arrows). (B, H, N) T lymphocytes were visualized by immunohistochemistry against CD3 (cluster of differentiation 3) (arrows). (C, I, O) Neutrophils are visualized by immunohistochemistry against myeloperoxidase (arrows). (D, J, P) Basophils were visualized by immunohistochemistry against Mcpt8 (mast cell protease 8) (arrows). (E, K, Q) Eosinophils are visualized by Luna stain (arrows). (F, L, R) Macrophages were visualized by immunohistochemistry against ionized calcium-binding adaptor molecule 1 (IBA1) (arrows). Bars = 100 µm.
Serial infestation of female P. leucopus with larval I. scapularis elicits eosinophil, neutrophil, basophil, T lymphocyte, and macrophage infiltrates. Leukocyte characterization in naïve skin and at primary and quaternary larval attachment sites in female mice using special stains and immunohistochemistry. (A, G, M) Mast cells are visualized with toluidine blue stain (arrows). (B, H, N) T lymphocytes were visualized by immunohistochemistry against CD3 (cluster of differentiation 3) (arrows). (C, I, O) Neutrophils are visualized by immunohistochemistry against myeloperoxidase (arrows). (D, J, P) Basophils were visualized by immunohistochemistry against Mcpt8 (mast cell protease 8) (arrows). (E, K, Q) Eosinophils are visualized by Luna stain (arrows). (F, L, R) Macrophages were visualized by immunohistochemistry against ionized calcium-binding adaptor molecule 1 (IBA1) (arrows). Bars = 100 µm.
Punch biopsies were collected from the pinnae of naïve mice and from larval attachment sites on the pinnae of mice infested once or four times. Inflammatory cell markers were quantified in pinnal biopsy samples using quantitative reverse transcriptase PCR (qRT-PCR) with primers specific for mast cells (mcpt4), T lymphocytes (cd3), neutrophils (mpo), basophils (mcpt8), eosinophils (embp), and macrophages (iba1) (Fig. 5). Specific oligonucleotide primers used are summarized in Table S4. This approach revealed that, for male mice, expression of all leukocyte markers was increased in infested skin compared to naïve skin, regardless of infestation number. Between one and four infestations, only markers for T lymphocytes (Fig. 5B), neutrophils (Fig. 5C), basophils (Fig. 5D), and eosinophils (Fig. 5E) were increased in males. In female mice, markers for T lymphocytes (Fig. 5B), neutrophils (Fig. 5C), basophils (Fig. 5D), eosinophils (Fig. 5E), and macrophages (Fig. 5F) increased in mice infested four times when compared to naïve samples; however, statistically significant increases were not detected for any leukocyte markers between one and four exposures. Mast cell and macrophage markers were not increased between one and four infestations in either sex. The mast cell quantification matched our observations with toluidine blue staining (Fig. 3A, G and M; Fig. 4A, G and M), showing that a robust mast cell infiltrate was not observed. Similarly, for macrophages, immunohistochemistry against IBA1 did not show increased macrophage influx to the larval bite site between the first ([Fig. 3L and 4L](#F3 F4)) and fourth ([Fig. 3R and 4R](#F3 F4)) infestations. Taken together, these findings suggest that basophils, eosinophils, neutrophils, and T lymphocytes may be important contributors in driving ATR against I. scapularis larvae.
*Repeated infestation in male and female P. leucopus is associated with increased expression of eosinophil, neutrophil, basophil, T lymphocyte, and macrophage transcripts. Analysis of leukocyte infiltrate at larval attachment sites in male and female mice using qRT-PCR to quantify gene expression in pinnal biopsies from naïve mice or from larval attachment sites. (A) Mast cell-specific transcript mast cell protease 4 (mcpt4). (B) T lymphocyte-specific transcript cd3. (C) Neutrophil-specific myeloperoxidase (mpo). (D) Basophil-specific transcript mcpt8. (E) Eosinophil-specific eosinophil major basic protein (EMPB). (F) Macrophage-specific transcript iba1. All qRT-PCR data points represent individual bite sites or biopsy samples collected from the pinnae of uninfested mice. Leukocyte marker quantification was analyzed using a linear regression model. *P < 0.05; **P < 0.01; **P < 0.001. ns, not significant.
Larvae fed on tick-sensitized hosts ingest greater numbers of leukocytes
Given the significant reduction in larval feeding success on sensitized P. leucopus, we next asked if tick resistance led to appreciable histologic changes within the midgut of feeding larvae. Larvae were collected from mice that were naïve or that had been serially infested for histologic examination. In larvae fed on mice without prior tick exposure, the midgut contained primarily erythrocytes during attachment (Fig. 6A) and at repletion (Fig. 6B and C), with only small numbers of nucleated cells admixed with the blood meal (Fig. 6C). However, in larvae fed on a mouse previously infested three times, the midgut contents contained greater numbers of nucleated cells during attachment (Fig. 6D) and repletion (Fig. 6E and F). Automated counting of nucleated cells within a defined area of the midgut revealed that the quantity of nucleated cells within the blood meal increases significantly with each infestation previously experienced by the host (Fig. 6; Fig. S5; Tables S1 and S2). Average nucleated cells within a 150 × 150 µm area of the midgut increased from 1.2 ± 0.2 to 18.5 ± 1.7 for larvae fed on males and from 3.1 ± 0.4 to 15.6 ± 1.7 for larvae fed on females between first and fourth infestations (Fig. 6G and H).
*Larvae fed on tick-sensitized hosts ingest more host leukocytes. Histologic images of attached and replete larvae. (A–C) Histology of larval cross sections from ticks fed on a naïve mouse (1st infestation) at 3-days post-attachment (A) and after repletion (B). (C) Nucleated cells shown in the midgut (arrow) at high magnification. (D–F) Histology of larval cross sections from ticks fed on a sensitized mouse (4th infestation) at three days post-attachment (D) and after repletion (E). (F) Nucleated cells with multilobulated or fragmented nuclei are shown in the midgut (arrows) at high magnification. H&E stain. A–B, D–E: bar = 100 µm; C, F: bar = 20 µm. (G–H) Nucleated cell counts within the midguts of replete larvae fed on male (G) or female (H) mice. Each data point represents the mean nuclear count from three sites in the midgut of a single replete larva. Nucleated cell counts were analyzed using a negative binomial generalized linear mixed-effects model. **P < 0.01; ***P < 0.0001. ns, not significant.
The majority of the nucleated cells were characterized by segmented nuclei (polymorphonuclear) and were morphologically distinct from the mononuclear midgut epithelial cells and hemocytes of the tick (39–41), indicating that they are of host origin. We were unable to detect host cell populations by immunohistochemistry or qRT-PCR, possibly due to degradation of the blood meal during digestion. Our findings demonstrated that tick resistance is associated with a shift in bloodmeal contents characterized by increased numbers of ingested host leukocytes.
ATR enhances larval acquisition of A. phagocytophilum and B. burgdorferi
A. phagocytophilum is an obligate intracellular pathogen that infects and propagates in granulocytes, particularly neutrophils, during mammalian infection (42). In contrast, neutrophils can kill B. burgdorferi through phagocytosis, oxidative bursts, hydrolytic enzymes, and neutrophil extracellular traps (43, 44). Since we observed a significant neutrophil component as part of the severe inflammation at the bite site on sensitized mice (Fig. 3 to 5) and significant changes in blood meal composition within replete larvae (Fig. 6), we asked whether these changes could change pathogen acquisition by I. scapularis. To address this, we infected tick-naïve and tick-sensitized mice with either A. phagocytophilum or B. burgdorferi by needle inoculation. This approach was chosen to control for variables that are associated with tick-bite transmission, including undefined infectious dose and the potential for unsuccessful tick attachment due to mouse grooming behavior (45, 46). Naïve larvae were allowed to feed to repletion on infected mice, and pathogen burdens were quantified. We found that larvae fed on previously sensitized female P. leucopus ingested 3.66 ± 0.90-fold more A. phagocytophilum than those fed on tick-naïve mice. Ticks that fed on male mice showed a similar trend (Fig. 7A and B); however, this difference was not statistically significant. When mice were infected with B. burgdorferi, the opposite trend was observed. We found that larvae fed on previously sensitized male mice ingested 46.93 ± 19.80-fold more B. burgdorferi than those fed on tick-naïve mice. While no difference was observed in larvae fed on naïve or sensitized female mice, there was a similar but non-significant increasing trend (Fig. 7C and D; Tables S1 and S2). Taken together, these data demonstrate that ATR impacts how many pathogens are ingested by naïve I. scapularis larvae from infected P. leucopus. Notably, although sex did not impact larval feeding success, it did influence pathogen load in the tick, suggesting that undefined variables between male and female mice exhibiting ATR influence A. phagocytophilum and B. burgdorferi transmission dynamics.
*P. leucopus ATR against I. scapularis larvae alters A. phagocytophilum and B. burgdorferi acquisition by ticks. Naïve larvae were fed to repletion on A. phagocytophilum-infected (A –B) or B. burgdorferi-infected (C–D) male and female P. leucopus that were either tick-naïve or sensitized with four previous larval infestations. Pathogen burdens were assessed via qRT-PCR and normalized to tick-naïve conditions. Pathogen burdens were analyzed using a parametric Welch’s t-test. Each data point represents a single replete larva. *P < 0.05; *P < 0.01. ns, not significant.
DISCUSSION
How a host species interacts with hematophagous arthropods impacts vector competence and pathogen movement within natural systems (47), both in ticks (48–50) and in other vectors like mosquitoes (51–54). In this study, we demonstrate that repeated exposure to I. scapularis larvae causes an increasingly severe inflammatory response in P. leucopus. This correlates with decreased larval feeding success characterized by fewer replete ticks and lower replete weights, which is consistent with previous reports (28, 30). We found that larvae fed on sensitized mice showed altered blood meal compositions with an increased amount ingested host leukocytes. Moreover, ATR caused increased B. burgdorferi and A. phagocytophilum ingestion by I. scapularis larvae in a sex-dependent manner. To our knowledge, this is the first time that ATR has been implicated in pathogen movement between native reservoir hosts and I. scapularis larvae.
Tick resistance in reservoir hosts has been largely overlooked in favor of using model host organisms (6, 13, 35). For example, guinea pigs exhibit an exaggerated resistance phenotype, with over 80% fewer ticks feeding to repletion (21, 26, 27). For studies aiming to develop tick-targeted vaccines, near-complete cessation of tick feeding on sensitized hosts is desirable, and in these cases, such host organisms are appropriate models. Natural tick-host pairings do not generally elicit such a high degree of resistance (36, 50). Nevertheless, the 58% to 70% reduction in larval feeding success observed in our study when larvae are fed on tick-sensitized hosts would result in fewer ticks within the environment. Tick-borne pathogen transmission within natural systems is a density-dependent phenomenon with greater numbers of infected ticks in the environment being associated with a greater risk of infection (55, 56). Previous studies have correlated reductions in questing nymphs ranging from 68% to 76% following acaricide treatment of reservoir hosts with a 53% to 96% reduction in Lyme disease risk (57–62). It is possible that the degree of larval resistance in reservoir hosts is sufficient to influence pathogen transmission risk. This may be especially true during periods when many tick-naïve hosts are entering the population, such as the bimodal peaks of P. leucopus reproductive activity in spring and autumn (63). Additionally, reduced replete weight is correlated with reduced fecundity (50, 64) and post-molt size (50, 65), which is notable because smaller ticks are more susceptible to desiccation because of their higher surface-to-volume ratio (66, 67). Though these factors were not directly evaluated in our study, they may also influence the number of questing ticks within the environment. Interestingly, the reduction in larval I. scapularis feeding success on tick-sensitized P. leucopus is similar to that seen in Dermacentor andersoni larvae fed on deer mice (Peromyscus maniculatus) and cottontail rabbits (Sylvilagus nuttallii) (50), in Ixodes trianguliceps fed on bank voles (Clethrionomys glareolus) (68), and on larval I. scapularis fed on meadow voles (Microtus pennsylvanicus) (28). This suggests that the phenomenon of tick resistance in native hosts spans across species within natural systems and may have an underappreciated role in the natural tick-pathogen-host dynamics.
Previous work shows that I. scapularis nymphs elicit increasingly severe inflammation at the bite site in tick-sensitized P. leucopus (31). However, this does not correlate with a drop in feeding success, in contrast to what we observed with larvae. Nymphal infestations did not show a basophil influx at bite site until four days post-attachment, and the majority were confined to the blood vessel lumens (31). We found basophil infiltrates at larval bite sites three days post-infestation, centered around the embedded hypostome, suggesting that basophils may play an essential role in mediating ATR against I. scapularis larvae. This is in agreement with previous findings in Mus musculus subjected to repeated infestation with Haemaphysalis longicornis larvae in which ablation of basophils resulted in an inability to develop tick resistance (69). This may suggest that older life stages are better able to suppress basophil infiltrate than immature tick life stages, facilitating their feeding success despite inflammation at the bite site.
Tick saliva plays an important immunosuppressive role during tick feeding (70–81). Qualitative and quantitative differences in saliva may be responsible for the differential impact of ATR on feeding success between tick life stages. Comparisons of the sialome between nymphal and adult H. longicornis revealed 30 proteins produced exclusively in nymph saliva, including the protease inhibitors serpin and cystatin, and the immunomodulatory alpha-1-acid glycoprotein 1. Seventy-four proteins were exclusively found in adult saliva, including the antioxidant catalase and immunomodulatory galectin-1 (82). Additionally, the volume of saliva produced by a feeding tick correlates with the volume of both the hemolymph and blood meal (83, 84). It is possible that the smaller volume and altered salivary content produced by I. scapularis larvae are not sufficient to abrogate the negative effects of the P. leucopus immune response, including infiltration and histamine release by basophils. Histamine and serotonin produced in sensitized hosts in response to tick attachment decrease both saliva production and feeding success in adult Dermacentor andersoni (85). The negative effects of the host immune response may be exacerbated in larvae due to their smaller size, higher surface-to-volume ratio, and limited lipid stores (66, 67).
Histamine-induced epidermal hyperplasia can prevent some ectoparasites from accessing host blood, as in the case of the northern fowl mite (Ornithonyssus sylviarum). Increasing inflammation and epidermal thickening separate the mite from the blood vessels of its avian host (86). It is possible that the lower feeding success we observed with larval I. scapularis on sensitized mice could be attributable to epidermal hyperplasia impeding access to the feeding lesion (6). The average length of larval I. scapularis hypostomes is 0.111 mm ± 0.037 mm (87). In our study, tick-sensitized mice exhibited sufficient epidermal thickening to potentially impact the ability of larvae to reach the feeding lesion (Tables S1 to S3). However, we observed that the epidermis immediately adjacent to the hypostome was often eroded or ulcerated with no appreciable displacement of the tick from the feeding lesion. It therefore remains unclear what effect, if any, epidermal hyperplasia has on larval attachment. The smaller size of the larval hypostome and cement cone may make dermal anchoring inherently less stable when compared to nymphs. However, given the epidermal loss and extension of granulocytes into the cement cone observed in this study, destabilization of the dermal-cement adhesion may be a more feasible explanation for failure to reach repletion.
Most studies evaluating host resistance to hematophagous arthropod feeding have focused on how inflammation impacts transmission of vector-borne pathogens from the arthropod to the host. For example, host resistance alters transmission outcomes for mosquito-borne arboviruses (88–90), sandfly-borne leishmaniasis (91–94), and tick-transmitted pathogens (46, 95–100). Tick resistance greatly reduces host susceptibility to pathogen transmission by infected ticks, including B. burgdorferi (guinea pigs and non-human primates) (46, 95, 96), Francisella tularensis (rabbits) (99), Babesia spp. (cattle) (97, 98), Anaplasma marginale (cattle) (98), and tick-borne encephalitis virus (M. musculus infested with R. appendiculatus) (100). Even in the absence of tick resistance, inflammation at nymphal I. scapularis bite sites of repeatedly infested P. leucopus reduced rates of B. burgdorferi transmission from the tick to the host by 83.3% (45). To our knowledge, our study is the first to evaluate the interplay between P. leucopus resistance to larval I. scapularis and pathogen ingestion by ticks.
We observed sex-specific variations in both tick resistance and pathogen transmission from mice to I. scapularis. In general, fewer larvae fed to repletion on females than males, which is consistent with previous reports (28, 101–104). We also found that larvae fed on previously sensitized, female P. leucopus ingested more A. phagocytophilum than those fed on tick-naïve mice. Enhancement of A. phagocytophilum acquisition from tick-sensitized female mice is consistent with the histologic changes that characterize ATR in both the host skin and replete ticks. A. phagocytophilum infects and replicates within neutrophils, eosinophils, and monocytes (105). As such, the increase in granulocytes at both the host bite site and within the replete larval midgut would be expected to enhance larval ingestion of A. phagocytophilum from tick-sensitized hosts. However, similar enhancement of A. phagocytophilum transmission was not observed in male mice, suggesting that additional factors must be at play.
Sex differences in immunity, pathogen kinetics, and host-pathogen interactions may also influence pathogen acquisition from infected hosts. General immunity differences between the sexes have been previously reported, with both female mice and humans mounting more robust innate and adaptive immune responses than males (106–109). As a result, females tend to be less susceptible to parasitic and viral infections (110–112). Differences in susceptibility also extend to tick-borne pathogens. Male C57BL/6 mice exhibit increased susceptibility to A. phagocytophilum infection with up to a 1.85-fold increase in infected neutrophils and significantly greater splenomegaly compared to infected females (113). If A. phagocytophilum burdens were sufficiently high in male P. leucopus, it is possible that any boost in acquisition provided by tick resistance could be muted.
Another possibility not explored in this study is the influence of infection on the inflammatory infiltrate at larval attachment sites. A. phagocytophilum can modify the activity of neutrophils by suppressing respiratory bursts (114), dysregulating degranulation (115), and interfering with surface selectin expression resulting in impaired transmigration from the vasculature (116, 117). Given its ability to modulate granulocyte behavior, it is possible that A. phagocytophilum could reduce leukocyte transmigration to larval attachment sites. Changes in the inflammatory infiltrate could help explain why tick-sensitized male mice did not show enhanced transmission of A. phagocytophilum to larvae, especially if higher A. phagocytophilum burdens in males lead to more profound alterations in neutrophil behavior. However, additional studies would be necessary to evaluate whether infection can influence the inflammatory response to ticks.
Interestingly, the sex-specific effects on pathogen transmission were reversed in the case of B. burgdorferi. We found that larvae fed on previously sensitized male mice acquired more bacteria than those fed on naïve P. leucopus, and no difference was observed between larvae fed on naïve or tick-sensitized females. This finding was unexpected, given previous studies demonstrating that prior tick exposure reduces the susceptibility of P. leucopus to B. burgdorferi transmission by infected nymphs (45). One possible explanation is that inflammatory mediators and/or DAMPs, such as reactive oxygen species (ROS) and L-serine in serum, may act as chemoattractants for B. burgdorferi, as is seen with H. pylori (118) and other enteric pathogens, including S. enterica, E. coli, and C. koseri (119, 120). Sex differences in the production of inflammatory mediators could conceivably lead to differences in chemotactic responses. For instance, males tend to produce more histamine in response to cutaneous allergens (121), have higher basal levels of ROS, and less efficient antioxidant mechanisms (122–124). Alternatively, it is possible that the tendency toward a less robust immune response in males could allow for more effective transmission of B. burgdorferi to feeding larvae. In M. musculus infected with B. burgdorferi, males showed both a higher percentage of infected tissues and higher cutaneous and visceral spirochete burdens (125). Pathogen transmission and acquisition dynamics may be further complicated by the immunomodulatory effects exerted by B. burgdorferi, as it attempts to subvert the host immune response. B. burgdorferi suppresses host IgG responses to facilitate its dissemination and persistence (126). Consequentially, the IgG response to unrelated antigens, such as the SARS-CoV-2 spike protein, is also impaired (127). If B. burgdorferi can immunomodulate P. leucopus in a similar manner, it could alter the host response to tick attachment and lead to unexpected tick-pathogen interactions.
The observation that ATR enhances larval B. burgdorferi acquisition from male P. leucopus contrasts with previous findings in M. musculus, where prior tick sensitization of B. burgdorferi-infected C57BL/6 J mice was correlated with decreased pathogen ingestion by I. scapularis larvae (128). Our findings suggest that tick-host-pathogen interactions observed in M. musculus cannot necessarily be translated to native host species like P. leucopus. Why transmission differences exist between M. musculus and P. leucopus is unclear. Both hosts have unique genetic backgrounds (inbred C57BL/6 mice vs. outbred P. leucopus) which, in turn, influence the immune response. For instance, T cells in C57/BL6 mice favor production of Th1 cytokines such as IFN-γ (129). Differences in immunity may alter the tick-host-pathogen relationship with implications for pathogen acquisition. Alternatively, some tick salivary proteins act as chemoattractants for B. burgdorferi (130, 131), and I. scapularis alters its sialome composition depending on the host species it feeds upon (132). Differences in mouse species and tick-sensitization status likewise may trigger variation in the sialome of feeding larvae, and this, in turn, may influence the presence and/or abundance of chemoattracts. Although it is clear that ATR, host species, and sex-specific differences influence pathogen ingestion by ticks, we did not examine pathogen transmission by infected nymphs to naïve hosts. Additional studies would be necessary to determine whether initial pathogen ingestion would correlate with enhanced pathogen maintenance through the molt and/or transmission to naïve mice. Additionally, while tick sensitization enhances initial pathogen ingestion by larvae, the negative impact of ATR on larval feeding success would result in fewer B. burgdorferi-infected ticks overall. This would be expected to reduce pathogen transmission risk in natural systems.
One limitation of this study is the route in which naïve or sensitized mice were infected. We chose needle inoculation to minimize uncontrolled variables that are associated with infection by tick bite, including undefined pathogen dose and the potential for unsuccessful tick attachment. With this approach, we were able to study how ATR responses affected pathogen movement into feeding larvae in a controlled, experimental setting. We acknowledge that a naïve mouse would not exist in the wild, as the only route for infection is through tick bite. However, this type of comparison is useful for isolating and examining immunological variables impacting pathogen ingestion by ticks. Additionally, while our tick placement protocol was designed to synchronize larval attachment, it may not precisely replicate natural infestation in which mice gradually encounter ticks as they move through the environment. Nevertheless, infesting each animal with 100 larvae did not result in an unnaturally high tick burden. Field studies conducted on P. leucopus collected in New York report an average burden of 10 to 40 larvae per mouse (102), peaking at up to 65 larvae per mouse in August and early September (133). None of the mice in our study supported more than 30 larvae during a single infestation, which is within the range of reported natural tick burdens.
Our findings showcase the complexity of factors influencing ATR and highlight that there is still much to be discovered. Sex, host species, degree of prior tick sensitization, and tick life stage all influence the presence and severity of ATR with implications for the vector-pathogen life cycles. Additionally, our study highlights the importance of including both sexes in ATR and tick-borne pathogen studies. Future studies accounting for the various factors influencing tick resistance and pathogen movement in natural systems will provide greater insight into the mechanistic underpinnings of ATR.
MATERIALS AND METHODS
Animal models
All mouse experiments were conducted in accordance with the guidelines and protocols approved by the American Association for Accreditation of Laboratory Animal Care (AAALAC) and by the Office of Campus Veterinarian at Washington State University (Animal Welfare Assurance A3485-01, IACUC-approved protocol #6097). White-footed mice (P. leucopus) were originally obtained from the Peromyscus Genetic Stock Center (University of South Carolina) to start a laboratory colony maintained at Washington State University. Mice were maintained in an AAALAC-accredited facility at Washington State University in Pullman, WA. All procedures were approved by the Washington State University Biosafety and Animal Care and Use Committees.
Pathogen-free I. scapularis larvae were obtained from Oklahoma State University (Stillwater, OK, USA). Ticks were kept in glass vials in an incubator maintained at 23°C and 95–100% relative humidity and a 16:8 h light:dark photoperiod.
Infestation procedure
Eight tick-naïve mice of each sex, between 6 and 12 weeks of age, were infested with I. scapularis larvae four successive times, separated by two-week tick-free intervals after the primary and tertiary infestation and one-week tick-free intervals after the secondary infestation. For each infestation, mice were anesthetized (1–2% isoflurane, 1.0 L per minute oxygen flow rate), and 100 larval ticks were placed around and between the ears and over the shoulders. Mice were maintained under anesthesia for 20 minutes following tick placement to allow time for attachment. Following recovery from anesthesia, mice were housed individually in specialized multi-cage systems to prevent tick escape. Systems included an innermost cage containing a raised metal rack suspended above 1 to 2 cm of water and a larger outer cage containing approximately 2 cm of water. Replete ticks were collected daily following natural detachment over the course of seven days. Ticks were either weighed and maintained to monitor post-feeding survival and molting or were saved in 10% neutral buffered formalin for histologic examination. A subset of male and female mice was euthanized on day three post-tick placement during the primary and quaternary infestations to examine leukocyte infiltrates at tick bite sites. For leukocyte quantification by qRT-PCR, four to six tick attachment sites per group were collected from the pinnae using a 4 mm sterile dermal biopsy punch (McKesson, 16-9840), placed in RNAlater Stabilization Solution (Thermo Fisher Scientific, AM7020), and frozen at −80°C. For comparison, biopsies from equivalent sites on the pinna were collected from control mice which were never infested with ticks. Bite sites collected from the head and ears for histologic examination were placed in 10% neutral buffered formalin for 24 hours prior to processing.
Bacterial maintenance and inoculation
A. phagocytophilum strain HZ was cultured in HL60 cells (ATCC, CCL-240) maintained in Roswell Park Memorial Institute (RPMI) 1640 medium supplemented with 1× Glutamax (Gibco, 35050061) and 10% heat-inactivated fetal bovine serum (FBS; Atlanta Biologicals, S11550). Cultures were maintained at a concentration between 1 × 10^5^ and 1 × 10^6^ cells per mL at 37°C and 5% CO_2_. On the day of infection, A. phagocytophilum was quantified using the previously described method (134, 135) and was liberated from HL60 host cells via syringe lysis with a 27-gage needle. Age-matched male and female P. leucopus mice were infected intraperitoneally with 1 × 10^7^ A. phagocytophilum HZ in 100 µL of PBS (Intermountain Life Sciences, BSS-PBS). Mice were rested for three days to allow them to reach peak bacteremia (136). On day 3 post-infection, between 25 µL and 50 µL of infected blood was collected from the lateral saphenous vein of each mouse, and A. phagocytophilum burdens were assessed via quantitative PCR (16S relative to mouse β-actin) to confirm infection and for the purpose of burden-matching (134, 137, 138). The following day, infected mice were infested with 100 larval ticks as described above. For analysis, ticks fed during quaternary infestations were normalized to ticks fed on burden-matched hosts during primary infestations.
B. burgdorferi B31 (strain MSK5) was cultured in modified Barbour-Stoenner-Kelly II (BSK-II) medium supplemented with 6% normal rabbit serum (NRS; Pel-Freez; 31126-5) and maintained at 37°C and 5% CO_2_. Spirochete growth was monitored daily using dark-field microscopy. Prior to infection, PCR was used to screen the plasmid profiles of B. burgdorferi cultures (139). Male and female age-matched P. leucopus mice were infected intradermally with 1 × 10^5^ low-passage B. burgdorferi B31 MSK5 in 100 µL 50:50 PBS/NRS. Mice were rested for four weeks to allow spirochetes time to disseminate (140), then infested with 100 larval ticks as described above.
Histology and immunohistochemistry
Skin samples and whole replete larval ticks were fixed in 10% neutral buffered formalin for 24 hours and embedded in paraffin. Five-micrometer sections were collected and either stained with H&E for routine histopathology, toluidine blue for detection of mast cells, Luna stain for detection of eosinophils, or were deparaffinized using clearing agent Clear-Rite 3 (Medix Corp, 6901) and rehydrated for immunohistochemistry. Heat-induced epitope retrieval (HIER) was performed by heating slides in a microwave at 700 watts for 20 minutes in either citrate buffer, pH 6.0 (Abcam, ab64214), or EDTA disodium salt dihydrate solution adjusted to pH 8.0 (Invitrogen, 15576028). Following a 30-minute room-temperature incubation with high protein blocking buffer (ThermoFisher, 00-4952-54), samples were incubated overnight at 4°C with one of the following antibodies diluted to 1:100 in high protein blocking buffer: α-myeloperoxidase polyclonal antibody (ThermoFisher, PA5-16672) for neutrophils, IBA1 polyclonal antibody (Fisher, PIPA527436) for macrophages, or MCP-8 monoclonal antibody (BioLegend, 647401-BL) for basophils. Following a room-temperature 10-minute 3% hydrogen peroxide block, samples were incubated with one of two secondary antibodies at room temperature for 1 hour: goat pAB to rabbit IgG conjugated to HRP (Abcam, ab97051) at 1:200 dilution or goat pAB to rat IgG conjugated to HRP (ThermoFisher, 31470) at 1:250 dilution. Slides were treated with the AEC Substrate Kit (Abcam, ab64252) with extended incubation for 25 minutes. Slides were stained with Mayer’s Hematoxylin Solution (H&E, Sigma, MHS32-1L) and cover-slipped using aqueous mounting medium for IHC (Abcam, ab64230).
For the detection of macrophages and basophils, the above protocol was slightly modified. Peroxide block was performed prior to antigen retrieval using 0.5% hydrogen peroxide in 85% ethanol incubated at room temperature for 30 minutes. For heat-induced epitope retrieval, the microwave was set to 800 watts for 25 minutes.
For the detection of T lymphocytes, deparaffinization and immunohistochemistry were performed by the Washington Animal Disease Diagnostic Laboratory Histology Core using the Ventana Discovery Ultra automated stainer (Roche). Heat-induced epitope retrieval (HIER) was performed by heating slides to 95°C in ULTRA CC1 antigen retrieval solution (Roche, 950124) for 56 minutes. Slides were incubated for 24 minutes at room temperature with CD3 polyclonal antibody (Dako, A0452) diluted to 1:400 in Antibody Dilution Buffer (Roche, ADB250). To visualize immunoreactivity, slides were treated using the UltraView Universal Alkaline Phosphatase Red Detection Kit (Roche, 760-501) according to the manufacturer’s instructions.
Histologic evaluation of larval attachment sites
Severity scores were assessed by a board-certified veterinary anatomic pathologist blinded to sample identity and experimental condition.
Severity was defined as follows:
Mild – Small numbers of inflammatory cells are present, and inflammation is confined to the dermis.Moderate – Small to moderate numbers of inflammatory cells are present and extend into the superficial subcutis and/or skeletal muscle.Severe – Numerous inflammatory cells are present with extension deep into the subcutis and skeletal muscle and/or crossing of the auricular cartilage.
To assess epidermal hyperplasia, measurements of epidermal thickness were collected adjacent to sites of larval attachment using an ocular micrometer at 400 × magnification. Measurements were taken in duplicate from each site and averaged. As a control, epidermal measurements were collected from the same animal at the same anatomical location, away from sites of tick attachment and from uninfested control animals at comparable locations. Epidermal measurements were taken perpendicular to and beginning at the basement membrane.
Analysis of larval midgut contents
To quantify nucleated cells within replete larval midguts, 10 replete larvae from each infestation on both male and female hosts were evaluated. Three 150 × 150 µm images were collected per larva centered within a separate midgut lobule. Nuclei were quantified in each image using ImageJ (parameters: pixel count, 50–infinity; circularity, 0.20–100).
Quantitative reverse transcriptase polymerase chain reaction (qRT-PCR)
RNA was extracted from skin biopsy samples and from replete ticks using the Direct-zol RNA MicroPrep Kit (Zymo Research, R2062). cDNA was synthesized from 300 to 500 ng. Total RNA was performed using the Verso cDNA Synthesis Kit (ThermoFisher, AB-1453). Bacterial burden and leukocyte transcripts were assessed by qRT-PCR with iTaq universal SYBR green Supermix (Bio-Rad; 1725125), with the primers listed in Table S4 and cycle conditions as recommended by the manufacturer. For leukocyte marker quantification, values were converted to logarithm base 10.
Statistical analysis
Statistical tests were done using the program R (version 4.2.2). Data were evaluated independently for each host sex. The number of larvae recovered from naïve and sensitized hosts and larval midgut leukocyte counts was analyzed using a negative binomial generalized linear mixed-effects model with the function glmer.nb (package MASS). Models treated the host’s prior tick exposure as a fixed effect and host ID, infestation time point, and tick cohort as random effects. The proportion of larvae molting successfully was analyzed using a generalized linear mixed-effects model with the function glmer (package MASS). The time taken to molt was analyzed using a negative binomial generalized linear mixed-effects model with the function glmer.nb (package MASS). Models treated the host’s prior tick exposure as fixed effects and host ID, host sex, and tick cohort as random effects. Replete larval weights were analyzed using a linear mixed-effects regression model with the function lmer (package lmerTest), treating the host’s prior tick exposure as a fixed effect, and host ID, infestation time point, and tick cohort as random effects. Epidermal thickness at larval attachment sites was analyzed using a linear mixed-effects regression model with the function lmer (package lmerTest), treating the host’s prior tick exposure as a fixed effect and biopsy sample ID as a random effect. Leukocyte marker quantification at larval bite sites by qRT-PCR was analyzed using a linear regression model with the function lm (included in base R). Post-hoc comparisons were performed using the function emmeans (package emmeans) for larval replete weights, molting success, molting time, midgut leukocyte counts, and qRT-PCR quantification of leukocyte markers. A. phagocytophilum and B. burgdorferi burdens were analyzed using a parametric Welch’s t-test. For all analyses, the cutoff for statistical significance was set at a P value of < 0.05. Descriptive statistics for each experiment are summarized in Table S1. Statistical models and results are summarized in Table S2.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1CDC. 2024. Tickborne disease surveillance data summary. Ticks. Available from: https://www.cdc.gov/ticks/data-research/facts-stats/tickborne-disease-surveillance-data-summary.html. Retrieved 7 Aug 2024.
- 2Kugeler KJ, Schwartz AM, Delorey MJ, Mead PS, Hinckley AF. 2021. Estimating the frequency of lyme disease diagnoses, United States, 2010-2018. Emerg Infect Dis 27:616–619. doi:10.3201/eid 2702.20273133496229 PMC 7853543 · doi ↗ · pubmed ↗
- 3Eisen RJ, Kugeler KJ, Eisen L, Beard CB, Paddock CD. 2017. Tick-borne zoonoses in the united states: persistent and emerging threats to human health. ILAR J 58:319–335. doi:10.1093/ilar/ilx 00528369515 PMC 5610605 · doi ↗ · pubmed ↗
- 4Trager W. 1939. Acquired immunity to ticks. J Parasitol 25:57. doi:10.2307/3272160 · doi ↗
- 5Ferreira BR, Silva JS. 1999. Successive tick infestations selectively promote a T-helper 2 cytokine profile in mice. Immunology 96:434–439. doi:10.1046/j.1365-2567.1999.00683.x 10233725 PMC 2326753 · doi ↗ · pubmed ↗
- 6Karasuyama H, Miyake K, Yoshikawa S. 2020. Immunobiology of acquired resistance to ticks. Front Immunol 11:601504. doi:10.3389/fimmu.2020.60150433154758 PMC 7591762 · doi ↗ · pubmed ↗
- 7Yoshikawa S, Miyake K, Kamiya A, Karasuyama H. 2021. The role of basophils in acquired protective immunity to tick infestation. Parasite Immunol 43:e 12804. doi:10.1111/pim.1280433124059 PMC 8244031 · doi ↗ · pubmed ↗
- 8Askenase PW, Bagnall BG, Worms MJ. 1982. Cutaneous basophil-associated resistance to ectoparasites (ticks). I. Transfer with immune serum or immune cells. Immunology 45:501–511.7061108 PMC 1555256 · pubmed ↗