Battling herbivores: the volatile story of tea
Nilesh D Gawande

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
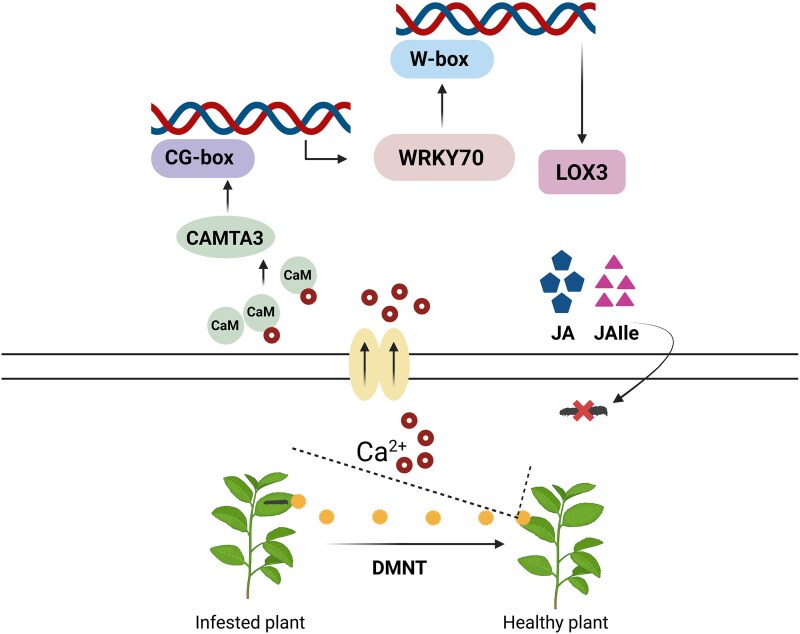 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTea Polyphenols and Effects
Tea plants (Camellia sinensis) are prone to herbivore attacks, which can cause severe yield losses. Plants possess strong systemic defense mechanisms to control such infestations. First, the plant recognizes the attack and then activates a defense response, which may include the production of toxic volatile compounds to deter herbivores or the formation of volatile compounds that attract their predators. The plant volatile compound (E)-4,8-dimethyl-1,3,7-nonatriene (DMNT) plays a critical role in defense against multiple insect pests, as well as in attracting natural enemies or predator species of the pest (Arimura et al. 2009). The activation of these responses often involves various signaling pathways, including ethylene, jasmonic acid (JA), and salicylic acid (Howe and Jander 2008; Fonseca et al. 2014).
The JA pathway possesses a strongly conserved mechanism for defense against arthropod herbivores (Howe and Jander 2008). JA and its bioactive derivatives accumulate at the damage site in response to herbivore infestation, which is perceived by the jasmonate co-receptors. The JA derivative (+)-7-iso-jasmonoyl-L-isoleucine (JA-Ile) is the main active JA derivative that controls vascular plant responses to herbivores and JA-regulated processes (Fu et al. 2022).
The tea geometrid (Ectropis obliqua) is a lepidopteran moth species and a common insect pest that feeds on the leaves and tender buds of tea gardens in China, causing deterioration in quality as well as yield losses in tea plants.
In this issue of Plant Physiology, Li et al. (2025) demonstrated how the volatile compound DMNT, induced by herbivore attack, activates the JA biosynthesis pathway in tea plants (Fig. 1). In previous studies, the authors determined that in response to E. obliqua infestation, tea plants release herbivory-induced plant volatiles, which cause an increase in JA and JA-Ile levels in surrounding non-infested plants, alerting them to the herbivory infestation. The analysis of volatile compounds released by E. obliqua using gas chromatography-mass spectrometry showed that DMNT was the prominently induced compound after the herbivory attack. DMNT-exposed tea leaves showed increased levels of JA and JA-Ile and induced JA biosynthesis genes, including CsLOX3. In contrast, JA biosynthesis gene expression was abolished in plants treated with JA inhibitors. JA inhibitor-pretreated plants showed more larval feeding than plants exposed to DMNT and did not exhibit an anti-herbivory response to DMNT, suggesting that E. obliqua infestation induces DMNT, activating the JA-dependent resistance in neighboring tea plants.
Transcriptome analysis of neighboring tree plants indicated that various transcription factors and JA biosynthesis genes were induced. A strong correlation was detected between the expression of CsLOX3, transcription factor CsWRKY70, and another gene. Subcellular localization using eYFP showed that CsWRKY70 is localized in the nucleus. The interaction between the transcription factor CsWRKY70 and the CsLOX3 promoter was confirmed through a yeast 1-hybrid assay and a dual luciferase assay. CsWRKY70 has a WRKY-DNA binding domain that recognizes the W-box motif, while CsLOX3 has a W-box cis-acting element. The electronic mobility shift assay demonstrated that CsWRKY70 directly binds to the W-box element in the CsLOX3 promoter region.
The silencing of the CsWRKY70 gene in tea leaves using an antisense oligonucleotide significantly reduced the expression of the CsLOX3 as well as the levels of JA and JA-Ile. In contrast, plants expressing CsWRKY70 showed the opposite effect, with higher levels of these compounds. Feeding E. obliqua larvae on the tea leaves with silenced CsWRKY70 resulted in increased leaf damage and a significant elevation in larval weights compared with the control. The exogenous application of DMNT in CsWRKY70-silenced plants resulted in reduced expression levels of CsWRKY70 and CsLOX3 along with the downregulation of the biosynthesis of JA and JA-Ile genes. This indicates that CsWRKY70 enhances resistance to E. obliqua through CsLOX3 expression and DMNT-mediated JA induction.
Furthermore, tea seedlings exposed to DMNT and other herbivore-induced plant volatiles showed that short exposure to DMNT induces CsWRKY70 and CsLOX3. DMNT enhances the luciferase activity of CsWRKY70, while other compounds reduce CsWRKY70 activity, indicating that DMNT specifically induces CsWRKY70.
During biotic stress conditions, such as plant-insect interactions, calcium (Ca²⁺) acts as a secondary messenger, and herbivory attacks induce changes in the cytosolic Ca²⁺ through various channels and signaling pathways (Malabarba et al. 2021; Kloth and Dicke 2022). Therefore, the authors measured transmembrane Ca²⁺ flux in mesophyll cells of tea leaves after exposure to DMNT. Increasing DMNT concentrations rapidly elevated Ca²⁺ influx in mesophyll cells. Pretreatment with LaCl₃, a competitive inhibitor of Ca²⁺ channels, reduced Ca²⁺ influx, indicating the involvement of Ca²⁺ channels in DMNT-induced Ca²⁺ influx. CsCML42 and CsCDPK1 were induced following short DMNT exposure. These results suggest that DMNT mediates signaling that triggers Ca²⁺ influx in mesophyll cells by regulating the expression of calmodulin and its associated protein kinases.
The CsWRKY70 promoter contains a CG-box cis-acting element, recognized as a specific binding site for calmodulin-binding transcription activator (CAMTA) transcription factors. Yeast 1-hybrid assays, electrophoretic mobility shift assay, and dual luciferase assays revealed that only CAMTA3 from tea plants binds to the CG-box of the CsWRKY70 promoter, indicating that CAMTA3 regulates CsWRKY70 expression. Expression analysis using qRT-PCR showed that DMNT exposure significantly induced CAMTA3, while silencing CAMTA3 with antisense oligonucleotide reduced the expression of CsWRKY70, and CsLOX3, as well as decreased the accumulation of JA and JA-Ile upon DMNT exposure. These findings suggest that CAMTA3 plays a critical role in regulating CsWRKY70 expression in response to DMNT and enhances JA and JA-Ile biosynthesis.
Overall, this study demonstrates that DMNT activates the CAMTA3-WRKY70-LOX3 module through the Ca^2+^ signaling pathway, resulting in JA and JA-Ile biosynthesis and activating herbivore resistance in tea plants. It will be interesting to further study the molecular mechanism of how herbivore-induced plant volatiles other than DMNT impart herbivore resistance in tea plants.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arimura G, Matsui K, Takabayashi J. Chemical and molecular ecology of herbivore-induced plant volatiles: proximate factors and their ultimate functions. Plant Cell Physiol. 2009:50(5):911–923. 10.1093/pcp/pcp 03019246460 · doi ↗ · pubmed ↗
- 2Fonseca S, Rosado A, Vaughan-Hirsch J, Bishopp A, Chini A. Molecular locks and keys: the role of small molecules in phytohormone research. Front Plant Sci. 2014:5:709. 10.3389/fpls.2014.0070925566283 PMC 4269113 · doi ↗ · pubmed ↗
- 3Fu W, Jin G, Jiménez-Alemán GH, Wang X, Song J, Li S, Lou Y, Li R. The jasmonic acid-amino acid conjugates JA-Val and JA-Leu are involved in rice resistance to herbivores. Plant Cell Environ. 2022:45(1):262–272. 10.1111/pce.1420234661303 · doi ↗ · pubmed ↗
- 4Howe GA, Jander G. Plant immunity to insect herbivores. Annu Rev Plant Biol. 2008:59(1):41–66. 10.1146/annurev.arplant.59.032607.09282518031220 · doi ↗ · pubmed ↗
- 5Kloth KJ, Dicke M. Rapid systemic responses to herbivory. Curr Opin Plant Biol. 2022:68:102242. 10.1016/j.pbi.2022.10224235696775 · doi ↗ · pubmed ↗
- 6Li B, Deng Y, Chai M, Ni H, Zhang C, Wang K, Luo L, Yu K, Du W, Guo D, et al A calcium-decoding module translates volatile DMNT into jasmonic acid-mediated herbivore resistance in Camellia sinensis. Plant Physiol. 2025:199(1):kiaf 386. 10.1093/plphys/kiaf 38640880200 · doi ↗ · pubmed ↗
- 7Malabarba J, Meents AK, Reichelt M, Scholz SS, Peiter E, Rachowka J, Konopka-Postupolska D, Wilkins KA, Davies JM, Oelmüller R, et al ANNEXIN 1 mediates calcium-dependent systemic defense in Arabidopsis plants upon herbivory and wounding. New Phytol. 2021:231(1):243–254. 10.1111/nph.17277.33586181 · doi ↗ · pubmed ↗