The tumor–microbe connection
Gerlanda Vella, Maria Rescigno

TL;DR
Tumor-associated bacteria influence cancer progression and treatment, but their exact roles and mechanisms are still unclear.
Contribution
This paper summarizes key findings and highlights outstanding questions about tumor-associated bacteria and their interactions in the tumor microenvironment.
Findings
Tumor-associated bacteria challenge the idea of tumors as sterile environments.
TAB influence immunity, metastasis, and treatment response in cancer.
Understanding TAB-TME interactions could lead to new cancer therapies.
Abstract
The discovery of tumor‐associated bacteria (TAB) challenges the traditional view of tumors as sterile environments. These microbes are engaged in a complex dialog with the other components of the tumor microenvironment (TME), influencing immunity, metastasis, and treatment response. Yet the precise mechanisms by which TAB influence tumor biology remains incompletely understood. Deciphering the complex host–microbe interactions could unlock novel therapeutic strategies to reshape the TME and improve treatment outcomes. Here we summarize the key findings in the field, highlighting the most outstanding questions regarding bacterial sources, the roles of TAB in cancer, and their interactions with the other cellular components of the TME. Tumor‐associated bacteria (TAB) dynamically influence cancer biology by modulating tumor progression, metastatic spread, and therapeutic efficacy. Their…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
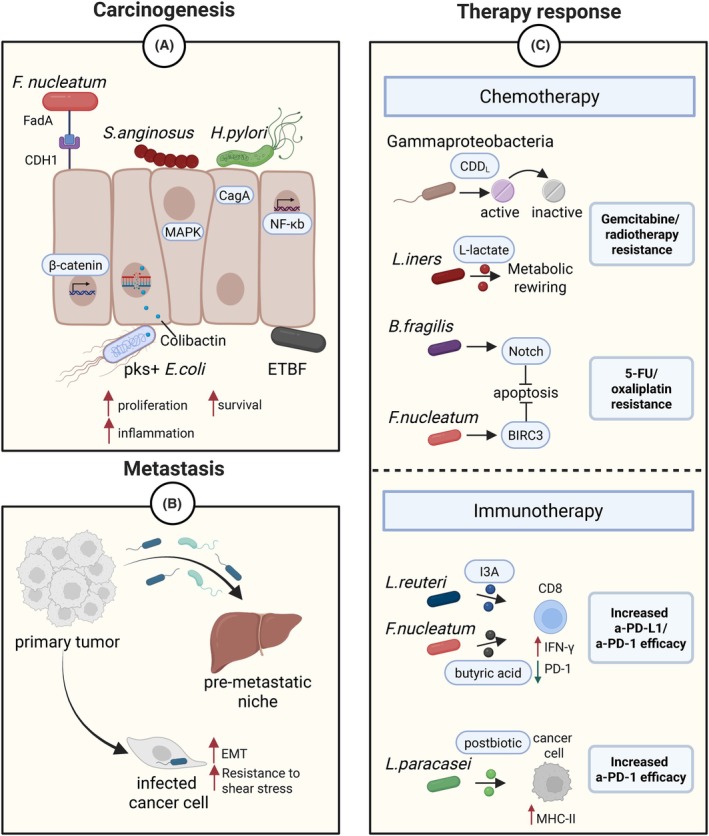 Fig. 1
Fig. 1- —European Molecular Biology Organization10.13039/100004410
- —Fondazione AIRC per la ricerca sul cancro ETS10.13039/100020581
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCancer Research and Treatments · Gut microbiota and health · Pancreatic and Hepatic Oncology Research
Introduction
1
For over a century, microorganisms have been detected within tumors, yet their presence was long met with skepticism. Only recently, thanks to advances in next‐generation sequencing (NGS), the intratumoral microbes are recognized as integral components of the tumor microenvironment (TME). The discovery of diverse microbiota—bacteria, viruses, fungi, and protozoa—coexisting within tumors has challenged the traditional view of tumors as sterile entities. We now appreciate tumors as complex, dynamic ecosystems in which microbial and human cells coexist and interact. This revelation raises important questions: What exactly are tumor‐associated bacteria doing in the TME? Are they merely passive bystanders, exploiting the tumor's resources, or are they active participants—shaping tumor progression, modulating immune responses, or even influencing therapy outcomes? Could certain microbes accelerate malignancy, while others suppress it? And if so, might targeting these microbial communities open new avenues for cancer treatment? Addressing these questions is essential to fully elucidate the role of the tumor‐associated microbiota in cancer biology.
Routes of bacterial colonization
2
The tumor mass, with its typical immunosuppressed, nutrient‐rich, and hypoxic conditions represents a favorable environment for bacteria colonization and persistence, particularly for anaerobic species. Moreover, microbial adaptations, including phagocytosis resistance and biofilm formation, further support their survival within tumors. Rather than being evenly distributed, bacteria tend to localize within poorly vascularized microniches enriched with protumoral macrophages, highlighting the importance of the local microenvironment within the tumor mass [1]. But where do these bacteria come from? Well, a univocal answer to this question doesn't exist, as three putative routes of bacteria dissemination have been proposed.
The bloodstream likely serves as the primary highway for bacterial entry to tumors. Bacteria may exploit tumor‐specific features for tumor colonization. The characteristic vascular permeability of tumors may facilitate passive bacterial entry, while specific molecular interactions, such as the binding of bacterial Fap2 to host GalNAc, enable active targeting of malignant tissues [2].
Alternative routes comprise mucosal barriers, particularly relevant for tumors at epithelial surfaces like colorectal cancer (CRC) or lung cancers, and the adjacent healthy tissues. Regarding this last route, there are two possibilities: either microbes migrate from healthy tissue into developing tumors, or local microbial communities residing in healthy tissue expand as the tumor forms. The directionality of this relationship remains unclear, as does the fundamental question of how bacteria normally colonize healthy internal organs. Notably, some species employ multiple colonization strategies. For instance, Fusobacterium nucleatum, an oral commensal, abundantly found in human CRC, can reach tumors via both the direct migration through the gastrointestinal tract and the hematogenous spread during transient bacteremia occurring during daily oral hygiene [3, 4].
Tumor establishment
3
The observation that certain bacterial species are more abundant in malignant tissues compared to adjacent healthy tissues—especially in advanced stage tumors—suggests that these microbes may play an active role in carcinogenesis and tumor progression. Probably one of the most prominent examples is F. nucleatum. The ability of this bacterium to influence cancer progression is largely mediated by its adhesin protein, FadA, which binds to CDH1 on the surface of host epithelial cells and activates the protumorigenic Wnt/β‐catenin signaling pathway [5]. Other examples include Helicobacter pylori and Streptococcus anginosus, both implicated in gastric cancer (GC) carcinogenesis. H. pylori translocates the CagA oncoprotein into gastric epithelial cells, triggering proinflammatory signaling pathways, while S. anginosus directly interacts with gastric epithelial cells and activates the MAPK pathway, promoting tumorigenesis [6, 7]. Similarly, enterotoxigenic Bacteroides fragilis (ETBF), particularly enriched in precancerous lesions in familiar adenomatous polyposis (FAP) patients, drives colonic tumorigenesis by activating IL‐17‐dependent NF‐κB signaling and inducing chronic inflammation [8].
Some bacteria can directly cause DNA damage. For instance, certain Escherichia coli strains carry the polyketide synthase (pks) pathogenicity island, encoding enzymes that synthesize colibactin. This toxin induces DNA adducts and double‐strand breaks, contributing to genomic instability in CRC [9] (Fig. 1A).
Bacterial modulation of cancer biology. (A) Certain bacteria can promote carcinogenesis by favoring cell proliferation/survival and promoting inflammation: F. nucleatum activates the Wnt/β‐catenin signaling pathway upon binding to CDH1 on the surface of host epithelial cells via the adhesin protein FadA; H. pylori triggers proinflammatory signaling pathways after translocating its CagA oncoprotein into gastric epithelial cells; S. anginosus activates the MAPK pathway; ETBF activates IL‐17‐dependent NF‐κB signaling; pks + E. coli induces DNA damage via its genotoxic colibactin metabolite. (B) Bacteria can promote metastasis by favoring the establishment of a premetastatic niche, inducing the EMT in cancer cells, and enhancing the shear stress resistance via host cytoskeletal remodeling. (C) Tumor‐associated bacteria can influence the response to chemotherapy and immunotherapy: Gammaproteobacteria inactivate gemcitabine via the cytidine deaminase (CDDL); B. fragilis promotes 5‐FU/oxaliplatin resistance through Notch activation and apoptosis inhibition; F. nucleatum induces 5‐FU resistance via the antiapoptotic factor BIRC3 upregulation; L. iners confers radiation/gemcitabine resistance via L‐lactate‐mediated metabolic reprogramming; L. reuteri enhances anti‐PD‐L1 efficacy via AhR‐mediated IFNγ production in CD8+ T cells; F. nucleatum‐derived butyrate and L. paracasei‐derived postbiotic improve anti‐PD‐1 response via PD‐1 downregulation in CD8+ T cells and MHC‐II upregulation in cancer cells, respectively.
Collectively, these findings suggest that bacteria may play a broader and more dynamic role in cancer development than previously appreciated. Unraveling the intricate crosstalk between tumor‐associated bacteria (TAB) and the host could reveal novel mechanisms of tumorigenesis and potentially uncover innovative strategies for cancer prevention and therapy.
Metastases
4
Either freely circulating in the bloodstream or residing within circulating cancer cells, bacteria can alter the metastatic process through three main mechanisms: (1) counteracting fluid shear stress via host cytoskeleton remodeling [10]; (2) creating a premetastatic niche [11]; (3) modulating the host epithelial‐to‐mesenchymal transition (EMT) [12] (Fig. 1B).
The idea that the tumor cell spread is not random dates back to the late 19th century with Paget's “seed and soil” hypothesis, where cancer cells (“seeds”) grow only in favorable organ environments (“soil”). Recent research expands this view, showing that bacterial communities in metastases are influenced more by the metastatic site than the primary tumor type [13]. This suggests the intriguing possibility of organ‐specific microbial tropisms, where certain bacteria are drawn by tissue factors like nutrients or signals, shaping a microenvironment that supports metastasis. Given its potential significance, this idea warrants further investigation to clarify the role of microbes in metastasis and how they might be targeted in cancer treatment.
Therapy response
5
When we think of therapy resistance in cancer, one of the first concepts that comes to mind is the innate or acquired resistance driven by genetic mutations within cancer cells. However, the mechanisms of drug resistance extend beyond tumor‐intrinsic genetic alterations and can also be influenced by TAB (Fig. 1C). This can occur by direct modification of the drugs. This is the case of Gammaproteobacteria that can convert the gemcitabine into its inactive form via the bacterial cytidine deaminase, limiting gemcitabine's therapeutic efficacy in a murine CRC model [14]. Alternatively, bacteria‐driven chemoresistance can rely on the host metabolic and cellular pathway alterations. Bacteroides fragilis, which is more abundant in chemoresistant than in responder CRC, promotes resistance to 5‐fluorouracil (5‐FU) and oxaliplatin, through the surface protein SusD/RagB that interacts with and activates the Notch1 prosurvival signaling pathway in host cells. Consistently, elimination of B. fragilis using targeted phage therapy successfully restored chemosensitivity in CRC mouse models [15]. In cervical tumors, Lactobacillus iners appears to contributes to resistance against both gemcitabine and radiation. It has been proposed that this effect could be mediated by bacterial L‐lactate, which reprograms cancer cells to rely on lactate metabolism, thereby potentiating lactate utilization over glucose under stress conditions, including irradiation [16].
In addition to regulating the efficacy of antitumor drugs, TAB can profoundly impact immunotherapy outcomes by modulating the host's immune system. For instance, Lactobacillus reuteri migrates to, colonizes, and persists within murine melanoma tissue, where it promotes antitumor immunity and enhances the efficacy of anti‐PD‐L1 therapy. This effect occurs through activation of the AhR in CD8^+^ T cells by the bacterial metabolite I3A, which leads to IFNγ production in a CREB‐dependent manner [17]. Moreover, a Lactobacillus paracasei‐derived postbiotic upregulates MHC‐II expression on murine breast cancer cells, boosting anti‐PD‐1 immunotherapy efficacy [18]. Notably, the same bacterium can exert opposing effects. For instance, F. nucleatum contributes to CRC carcinogenesis and impairs 5‐FU efficacy by upregulating the host's antiapoptotic factor BIRC3 [19], yet it may also sensitize microsatellite‐stable (MSS) CRC tumors to anti‐PD‐1 immunotherapy. This is mediated by the bacterial butyric acid, which enhances CD8^+^ T cell activity through TBX21‐mediated PD‐1 downregulation. Consistently, high intratumoral F. nucleatum levels correlate with favorable responses, suggesting its potential as a biomarker for immunotherapy in MSS CRC [20]. In light of these observations, the TAB should be considered a potential stratification factor when selecting anticancer therapies. Screening for specific microbial signatures may enhance the ability to predict treatment outcomes and guide personalized therapeutic strategies. An additional layer of complexity arises from the variability in bacterial load across different tumor types. While the presence of certain microbes is clearly linked to therapy resistance or efficacy in tumors with high bacterial load, such as melanoma and CRC, the extent to which this effect is relevant in tumors harboring low bacterial loads remains uncertain. It is plausible that a threshold level of microbial burden is required to significantly influence therapy response.
Concluding remarks
6
Growing appreciation is being given to the TAB within the TME, as they can play a dual role—either promoting tumor growth and therapy resistance or supporting antitumor immunity. Targeted strategies to eliminate harmful TAB, such as using antibiotics or bacteriophages, while preserving beneficial bacteria, are therefore essential. Despite advances, several key questions remain unresolved: What evolutionary pressures enable certain bacteria to adapt to the tumor niche? How do TAB interact causally and spatially with other TME components, like immune and stromal cells? Do specific bacteria show preference for certain metastatic sites, and what governs this tropism? Moreover, can microbial signatures within tumors serve as reliable biomarkers for prognosis or treatment response? Alongside these, the dynamic interactions within bacterial communities themselves—whether competitive or cooperative—must also be considered. Ultimately, understanding the complex roles and interactions of TAB in the TME may reveal novel molecular targets for innovative anticancer therapies.
Conflict of interest
MR is the founder and CSO of Postbiotica srl.
Author contributions
GV conducted the literature review and wrote the article. MR provided expert feedback, revised the article, and approved the final version.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Galeano Niño JL , Wu H , La Course KD , Kempchinsky AG , Baryiames A , Barber B , et al. Effect of the intratumoral microbiota on spatial and cellular heterogeneity in cancer. Nature. 2022;611(7937):810–817.36385528 10.1038/s 41586-022-05435-0PMC 9684076 · doi ↗ · pubmed ↗
- 2Abed J , Emgård JEM , Zamir G , Faroja M , Almogy G , Grenov A , et al. Fap 2 mediates Fusobacterium nucleatum colorectal adenocarcinoma enrichment by binding to tumor‐expressed gal‐Gal N Ac. Cell Host Microbe. 2016;20(2):215–225.27512904 10.1016/j.chom.2016.07.006PMC 5465824 · doi ↗ · pubmed ↗
- 3Abed J , Maalouf N , Manson AL , Earl AM , Parhi L , Emgård JEM , et al. Colon cancer‐associated Fusobacterium nucleatum may originate from the oral cavity and reach colon tumors via the circulatory system. Front Cell Infect Microbiol. 2020;10:400.32850497 10.3389/fcimb.2020.00400 PMC 7426652 · doi ↗ · pubmed ↗
- 4Rubinstein MR , Baik JE , Lagana SM , Han RP , Raab WJ , Sahoo D , et al. Fusobacterium nucleatum promotes colorectal cancer by inducing Wnt/β‐catenin modulator annexin A 1. EMBO Rep. 2019;20(4):e 47638.30833345 10.15252/embr.201847638 PMC 6446206 · doi ↗ · pubmed ↗
- 5Rubinstein MR , Wang X , Liu W , Hao Y , Cai G , Han YW . Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E‐cadherin/β‐catenin signaling via its Fad A adhesin. Cell Host Microbe. 2013;14(2):195–206.23954158 10.1016/j.chom.2013.07.012PMC 3770529 · doi ↗ · pubmed ↗
- 6Javaheri A , Kruse T , Moonens K , Mejías‐Luque R , Debraekeleer A , Asche CI , et al. Helicobacter pylori adhesin Hop Q engages in a virulence‐enhancing interaction with human CEACA Ms. Nat Microbiol. 2016;2:16189.27748768 10.1038/nmicrobiol.2016.189 · doi ↗ · pubmed ↗
- 7Fu K , Cheung AHK , Wong CC , Liu W , Zhou Y , Wang F , et al. Streptococcus anginosus promotes gastric inflammation, atrophy, and tumorigenesis in mice. Cell. 2024;187(4):882–896.e 17.38295787 10.1016/j.cell.2024.01.004 · doi ↗ · pubmed ↗
- 8Chung L , Thiele Orberg E , Geis AL , Chan JL , Fu K , De Stefano Shields CE , et al. Bacteroides fragilis toxin coordinates a pro‐carcinogenic inflammatory Cascade via targeting of colonic epithelial cells. Cell Host Microbe. 2018;23(2):203–214.e 5.29398651 10.1016/j.chom.2018.01.007PMC 5954996 · doi ↗ · pubmed ↗