Hidden transmissions of Pseudomonas aeruginosa ST111 –the importance of continuous molecular surveillance
Jasmin Kaur Jasuja, Eva-Maria Klupp, Martin Aepfelbacher, Knut Kurt William Kampe, Michael Fabian Nentwich, Stefan Kluge, Johannes Karl-Mark Knobloch

TL;DR
A study in a German hospital found hidden transmission of a dangerous Pseudomonas aeruginosa strain through molecular surveillance, revealing the importance of combining genetic and epidemiological methods.
Contribution
The study demonstrates how molecular surveillance can detect hidden hospital transmissions of Pseudomonas aeruginosa ST111 that standard methods miss.
Findings
A cluster of Pseudomonas aeruginosa ST111 blaVIM−2 isolates was identified through whole-genome sequencing, revealing sink-to-patient transmission.
Standard surveillance missed the outbreak, but molecular methods uncovered a prolonged transmission event in ICU rooms.
Environmental sampling and reconstruction efforts successfully eliminated the clone from the hospital.
Abstract
A series of transmission of Pseudomonas aeruginosa ST111 blaVIM−2, previously undetected by standard surveillance, was discovered in a tertiary care hospital in Northern Germany through molecular genetic monitoring. Hence, environmental sampling was initiated to find the source of infection. First, routine epidemiological data ruled out patient-to-patient transmission and two initial diagnoses were assessed as externally acquired. After the discovery of the highly related cluster by whole genome sequencing, a more detailed epidemiological analysis was carried out, including previous hospitalizations. An environmental investigation was initiated due to a possible connection of transmissions with an intensive care unit. Between 2018 and 2023 16 clinical isolates of Pseudomonas aeruginosa ST111 blaVIM−2 were identified of which 12 isolates belonged to ST111 carrying an In59-like…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
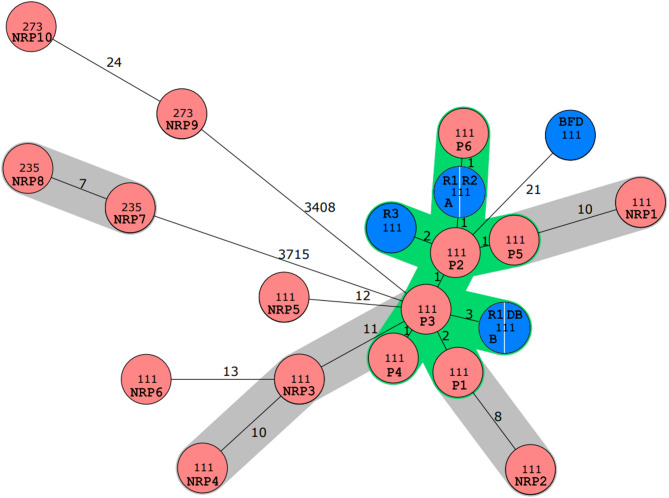 Figure 1
Figure 1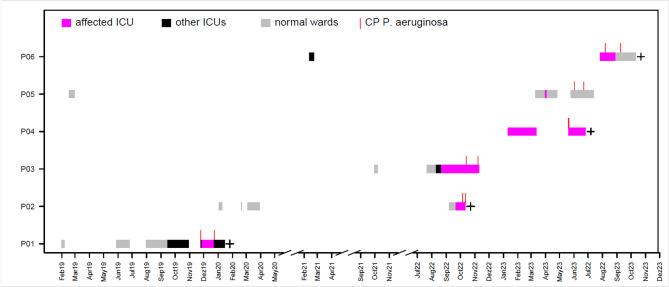 Figure 2
Figure 2- —Universitätsklinikum Hamburg-Eppendorf (UKE) (5411)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Bacterial biofilms and quorum sensing · Pharmaceutical and Antibiotic Environmental Impacts
Background
Pseudomonas aeruginosa is a leading cause of healthcare-associated infections (HAIs) and is of particular concern in critical care settings. As an important cause of healthcare-acquired infections in intensive care units (ICUs), P. aeruginosa demonstrates a remarkable ability to rapidly develop resistance to various antimicrobials, driven by chromosomal mutations and the acquisition of resistance genes encoded on mobile genetic elements such as plasmids or transposons [1–6]. The Centres for Disease Control and Prevention (CDC) and the European Centre for Disease Prevention and Control (ECDC) define multidrug resistance (MDR) as co-resistance to at least one agent in three out of eight antimicrobial categories [7].
MDR P. aeruginosa, including P. aeruginosa clones harbouring plasmid-encoded carbapenemases, are prevalent globally and typically observed in hospital environments. These clones are selected within hospitals due to antibiotic selection pressure and favourable growth conditions [8]. Thereby, in P. aeruginosa isolates spontaneously developing phenotypic resistance by mutations during antibiotic therapy must be distinguished from isolates carrying specific resistance genes like carbapenemases [9]. The emergence of widely disseminated carbapenemase producing P. aeruginosa (CP-PA) strains, designated as high-risk clones, has made whole-genome sequencing (WGS) an essential tool for understanding their epidemiology. Several high-risk clones are identified by their sequence types (STs) and have emerged on an international scale. Most hospital outbreaks in Europe are associated with multi locus sequence types (MLST) ST111, ST175, ST233, ST235, ST277, ST357, ST654 and ST773 [2, 8, 10]. CP-PA strains are frequently detected in wastewater and rinse water, where they form multi-bacterial biofilms in plumbing systems [11]. Within these protective and impenetrable biofilms, Enterobacterales and P. aeruginosa strains exchange carbapenemase-encoding plasmids [11]. During outbreaks, clones associated with the aforementioned STs are often found causing infections in vulnerable patient populations within ICUs. In a systematic review by Büchler et al. 100 out of 126 included studies screened the environment in outbreak situations and in all but three contaminated environment was identified as the primary source [12]. Hence, environmental screening was identified as an important outbreak control measure [12].
Here, we report a hidden outbreak with metallo-β-lactamase blaVIM−2-producing and qacE∆1-harbouring P. aeruginosa ST111 in a German tertiary care hospital, uncovered by perennial routine core genome multilocus sequencing typing (cgMLST), involving sink-to-patient transmission. To identify the outbreak source, an epidemiological investigation was initiated. Environmental samples were collected from the relevant ICUs, and the identified P. aeruginosa strains were analysed using cgMLST.
Results
Outbreak description
Between July 2018 and August 2023, a total of 131 non-repetitive phenotypically carbapenem-resistant P. aeruginosa (CR-PA) isolates cultured from clinical specimens of an adult hospital were analysed by cgMLST. Among these, carriage of carbapenemase genes were identified in 29 isolates (22.1%), thereof 16 isolates harbouring the blaVIM−2 gene (55.2%), four isolates harbouring blaVIM−1 (13.8%), and two isolates harbouring blaVIM−5 (6.9%). The predominant ST was ST111 (n = 14), followed by ST273 (n = 5). cgMLST identified a cluster (≤ 11 different alleles) of eleven ST111 CP-PA blaVIM−2 isolates, all also harbouring qacE∆1, a gene encoding an efflux pump for quaternary ammonium compounds (QAC). Six out of eleven blaVIM−2P. aeruginosa ST111 isolates were identified as highly related with a maximum allelic distance of three alleles (Fig. 1). In initial routine epidemiological categorization of these closely related isolates four isolates were categorized as possible nosocomial acquisition without direct epidemiological link to other patients, whereas two isolates were categorized as ‘externally acquired’ as patients were re-admitted or hospitalized after external stays. Due to the confirmed highly close relationship of the individual isolates but first recovery months and even years apart a more detailed epidemiological re-investigation was initiated, focusing on these isolates.
Fig. 1. Minimum spanning tree of blaVIM-2-positive Pseudomonas aeruginosa of a tertiary care hospital in northern Germany. A total of 16 Pseudomonas aeruginosa blaVIM-2 clinical isolates (red isolates, P1-6: patient 1 to 6; NRP1-10: non-related patient 1 to 10), were identified by WGS in clinical isolates from July 2018 to August 2023 and were analysed by cgMLST. A total of 4478 core genes were used. The grey shading marks clusters with the published threshold for clonal relatedness (≤ 11 distinct alleles). Within, a highly related cluster of ≤ 3 distinct alleles (green shading) six patient isolates (P1-P6) clustered with five environmental samples (blue isolates R1 = room 1; R2 = room 2; R3 = room 3; DB = dialysis bag; BFD = bedpan flushing device)
Epidemiology
The dates of first detection of the six highly related isolates were ranged between December 2019 and August 2023. Initial microbiological detections were made from relevant clinical specimens including the respiratory tract, skin, wounds and blood cultures (Fig. 2). An initial screening at admission was only conducted in two out of six patients, proving colonization with CP-PA. Hence, most of these cases were classified as nosocomial according to the German hospital infections surveillance system (KISS). The first externally acquired case was first hospitalized early 2019 and had an infection 298 days after the initial admission with intermittent hospitalization. The second externally acquired case was admitted early 2023 and infection with the outbreak strain was detected 130 days later with intermittent hospital stays. The initial detection in nosocomial-acquired cases occurred 8, 11, 29 and 85 days post admission (median: 20 days), respectively, and were all found in clinical specimen. Retrospective analysis showed that all six patients had infections caused by P. aeruginosa ST111 blaVIM−2, sourced from various clinical conditions, such as surgical site infections, acute respiratory distress syndrome (ARDS), pneumogenic sepsis, pneumonia and anastomosis- and respiratory insufficiency. Of these, four patients died in the further course of their hospital stay. Whether the ST111 infection was the cause of death cannot be ruled out with certainty.
Fig. 2. Line list of patients with confirmed transmission events of ST111 Pseudomonas aeruginosa bla_VIM-2_. A total of six patients were affected (P1-P6). The broad horizontal lines mark the respective stays in a normal ward (grey) of the affected intensive care unit (magenta) or another intensive care unit in the hospital. The vertical red lines indicate the respective first and last detection of a CP-PA in the affected patients. The black cross marks patients, who died in the course of their hospitalization
We subsequently investigated the wards where the initial detections occurred, and it was found that four of the six detections were from ward A in the years 2022 and 2023, with the remaining two from wards B in 2019 and D in 2023, respectively.
Since these findings did not immediately point to a clear source for possible transmissions, previous hospital stays were reviewed, particularly as the affected patients had been hospitalised for extended periods. The review indicated that the isolates detected on wards B and D were also associated with prior stays on ward A (Fig. 2). Interestingly, all affected patients could be associated with a stay in two adjacent patient rooms on ward A. As a result, environmental sampling was initiated on ward A.
Environmental sampling and molecular epidemiology
The extensive environmental examinations (see methods) were conducted twice in November 2023 and comprised 36 environmental samples (15 specimen from sinks of adjacent bathroom across seven rooms, six specimen from wardrobes in the bathrooms, one medical product kept in one of the patient room, two from a toilet in a patient room and twelve specimen from a sluice room) (Supplementary Fig. 1). The wastewater piping of rooms R1 and R2 are connected horizontally to the same wastewater line. Room R3 is separately connected to a wastewater pipeline, which merges only with the line serving R1 and R2 in the basement. During environmental sampling no positive screened patient was occupying the affected rooms and ward, respectively. Eleven environmental CR-PA isolates were identified. Thereof, two isolates were non-carbapenemase producing P. aeruginosa. In total, nine CP-PA isolates were identified with blaVIM−2 (n = 5) being the predominant carbapenemase. MLST by whole genome sequencing uncovered that all environmental blaVIM−2P. aeruginosa belonged to ST111. P. aeruginosa ST111 blaVIM−2 was found in three out of six sinks in patient rooms, on the surface of a dialysis therapy bag kept near the washbasin in patient’s bathroom, and in the sink of a bedpan flushing device in the sluice room. Applying a threshold for high relation of ≤ 3 different alleles, we identified five environmental blaVIM−2 and qacE∆1 positive P. aeruginosa ST111 isolates clustering with clinical isolates of patients hospitalised between 2019 and 2023. The four environmental samples were obtained from three washbasin sinks and from the exterior of a dialysis bag stored above a washbasin. All four environmental P. aeruginosa ST111 blaVIM−2 isolates were found in three patient rooms (R1-R3). Thereof, all patients had an associated stay for rooms R1 and R2, but none for R3. One specimen from the sluice room was positive for ST111 but did not harbour any carbapenemase and cluster distance threshold was > 12.
In order to assess whether plasmid transfer was involved in the transmission events, a detailed analysis of the resistance gene profile was carried out and attempts were made to determine the location of blaVIM−2. The resistance gene profile was almost identical between the ST111 isolates, including identical resistance-associated mutations and a profile of genes associated with reduced sensitivity to biocides and heavy metals (Supplementary Table, orange section). Analysis of the immediate genomic environment of blaVIM−2 revealed for ten of the ST111 isolates contigs so small (930 to 1539 bases) that it was not possible to evaluate their location. For eight isolates, assembly resulted in larger contigs (15 to 50 kb). The highly related isolates showed homology to chromosomally integrated In59-like class 1 integrons (Supplementary Fig. 2; Supplementary Table green section), which carry the resistance genes aac(6’)-29, qacE∆1and sul1 in addition to blaVIM−2 [13]. However, de novo assembly revealed an In59-like integron without blaVIM−2 in some strains, as well as some possible insertions and deletions in this region (Supplementary Fig. 2; Supplementary Table green section). This observation may also be due to the limitations of short read sequencing. As screening for plasmid replicon sequences using PlasmidFinder resulted in no hits for known plasmids in the database chromosomal integration of blaVIM−2 is suggested.
Due to the positive environmental findings, re-sampling of the environment was initially planned. However, from April to May 2024 the ward was closed for renovation and conversion into a regular ward. During this time, no patients were admitted to these rooms and consequently washbasins were not in use. Microbiological controls after reopening the ward did not yield additional P. aeruginosa ST111 blaVIM−2 isolates in six investigations over a period of twelve months.
Discussion
In this investigation, we uncovered a hidden prolonged outbreak involving the high-risk clone P. aeruginosa ST111 blaVIM−2 in a hospital in northern Germany, which had not been recognised as an outbreak by routine epidemiological surveillance. A purely epidemiological assessment was hindered by its prolonged nature over five years going along with patient transfers, external stays, and re-admissions with colonization or infection possibly being misclassified as externally acquired. Hence, epidemiological assessment alone carries the risk to detect only a fraction of the actual transmissions. The frequent occurrence of P. aeruginosa ST111 blaVIM−2 isolates, defined as highly related cluster in molecular surveillance, prompted a closer examination of its epidemiology. From an epidemiological perspective, patient-to-patient transmission was ruled out as there was no direct contact between these patients, all of whom had been hospitalised in ward A but during different periods of time. The investigation of further common procedures like dialyses and bronchoscopy were also excluded, as not every aforementioned patient was dialysed or got a bronchoscopy, especially prior to infection with P. aeruginosa ST111 blaVIM−2. However, all patients were associated with a stay in two adjacent patient rooms in ward A. As a result of this retrospective epidemiological analysis, repetitive environmental sampling was performed on ward A.
The observation of allelic differences far less than the distance threshold for genomic relatedness of ≤ 11 different in five environmental and clinical isolates of P. aeruginosa ST111 blaVIM−2 proved a nearly identical clone and confirmed sink-to-patient transmission for these cases [14, 15]. Indeed, sink colonisation by MDR Gram-negative bacteria is well documented and is considered as a potential transmission route, frequently associated with HAIs and posing a risk even in non-outbreak situations [16, 17]. Efforts to replace or eradicate contaminated sinks have been employed, but often fail [18], and it is even recommended that ICU rooms should not be equipped with sinks [19]. Our findings align with this, as the environmental P. aeruginosa ST111 blaVIM−2 strain remained detectable in 2023, despite its initial molecular detection occurring four years earlier. However, only six P. aeruginosa ST111 blaVIM−2 infections were traced back to specific sinks in the ICU, though it is assumed that ST111 persists throughout the year in patients [20]. If the infections with ST111 were indeed hospital-acquired, the initial classification of some cases as ‘externally acquired’ under the KISS definition would no longer apply. However, the environmental samplings showed ten different P. aeruginosa ST111 blaVIM−2 isolates indicating the overall frequency of sink-to-patient transmission was low but increased with the length of hospital stay as R1 and R2 was often occupied by patients with long-term stays, while patients in R3 did not. However, Rath et al. analysed the toilet-to-patient transmission rate of ST235 blaFIM−1 and ST309 non-carbapenemase producing P. aeruginosa in a bone marrow transplant unit with cgMLST, showing only a low-genetic diversity but only three toilet-to-patient-transmission over six years [21]. Indeed, our reported ward was originally built as an intermediate care unit and was converted into an intensive care unit later on. As a result, the washbasins were not located directly in the patient’s room but in a separate bathroom, which might have reduced the frequency of transmission.
In addition, alternative transmission routes must be considered. The positive screening of a dialysis therapy bag, placed above the washbasin suggests a possible transmission route, though not all patients were receiving dialysis. Rather, the dialysis therapy bag may represent a potential sink-to- environment-transmission, which might have been favoured by different pressure conditions leading to sink-to-environment-transmission. Bronchoscopy was excluded as a common transmission route as well as other environmental sources on epidemiological basis. However, it should be noted that environmental investigation was initiated four years after the first detection of P. aeruginosa ST111 blaVIM−2 and hence, potential transmission routes might have been missed meanwhile.
Interestingly, the first P. aeruginosa ST111 blaVIM−2 isolate was already detected in 2019 in a patient in screening and blood culture, which was conducted on admission to ward A and was declared as externally acquired since the patient was transferred from an external hospital where the patient was treated for one month. Before that, the patient had hospital stays in our hospital, but remained on normal wards. A P. aeruginosa was never detected. All other cases in the cluster occurred after the admission of the index case bringing up the discussion that first the outbreak stem was transmitted from patient to sink following hidden sink-to-patient transmission and leading to the hidden outbreak. We found a median duration of 20 days from admission to infection with the outbreak strain, which is lower than reported by Volling et al. [22]. However, Volling et al. reported a non-ST111 P. aeruginosa outbreak strain. In the two externally acquired cases colonisation time a maximum of 298 days and 130 days, respectively, is suggested. Notably, during this period both patients were not continuously hospitalized, making it hard to estimate the colonization-to-infection-period.
In our study, the qacEΔ1 resistance gene was detected in ST111 isolates as part of the In59-type integron, similar to the findings of Rath et al. However, it remains unclear whether this gene contributes to resistance against QAC-based disinfectants or plays a role in outbreak promotion even if a QAC-based disinfectant was used in routine hospital disinfection.
ST111 as well as other ST’s multi-resistance is well-documented, but association to resistance gene qacE∆1 has not been reported so far. Hence, further research is needed to elucidate any direct connections between this clone and qacEΔ1.
The observation of P. aeruginosa ST111 blaVIM−2 isolates with confirmed sink-to-patient transmission in an ICU-setting highlights the vulnerability of ICU patients and the conducive environment for antibiotic selection pressure. Previous studies have similarly documented the predominance of ST111 in ICUs and other high-risk wards, including in Germany, where ST111 blaVIM−2 was identified in a cluster of 15 isolates as well as in a Greek hospital [14, 21, 23]. ST111 strains are frequently associated with carbapenemase production, particularly blaVIM−2, leading to multidrug resistance [24, 25]. By chance, the transmission series might be interrupted by eliminating of the clone from the ward during a reconstruction project.
Limitations
This study has several limitations. First, environmental sampling was conducted four years after the first detection of the high-risk clone in clinical isolates. As a result, earlier environmental isolates and sources may have been missed, limiting our ability to accurately determine the duration of the high-risk clone’s presence in the hospital environment in- and outside the sink. To address this, we propose regular environmental sampling to allow for the timely identification of high-risk clones.
Second, while all patients included in the study were infected with the high-risk clone, only two out of six patients were initially screened for colonisation. Therefore, it is unclear whether transmission of the high-risk clone frequently leads to direct infection or if infection regularly resulted from previously undetected colonisation. Hence, we suggested that long-term patients should be screened during their ICU stay. However, further investigation is needed to determine the required time from colonisation to infection, as well as the trigger factors transforming a colonisation into an infection.
Third, during de novo assembly, blaVIM−2 was located in a chromosomally integrated In59-like integron in some strains. In strains without detectable integration into In59, it is unclear whether this is an artefact of the assembly or whether genetic events have actually taken place. However, the three resistance genes blaVIM−2, qacEΔ1 and sul1 were detected in all ST111 strains. Current literature as well as our detailed analysis of the resistance gene profile does not provide sufficient data directly linking qacEΔ1 to ST111 P. aeruginosa and the role in hospital transmission and infection. Further research is required to elucidate these questions.
Fourthly, the outbreak took place during the pandemic, which led to a change in the usual patient clientele.
Conclusion
Our report highlights the importance of molecular surveillance, which is more sensitive in detecting high-risk clones compared to conventional epidemiological assessment, which is often hampered by clinical processes such as internal and external transfers and prolonged hospital stays. Epidemiological surveillance should account for several years of data, particularly in cases of high genomic relatedness, to detect silent transmissions at an early stage. Colonisations and infections initially classified as externally acquired may, upon retrospective analysis, prove to be hospital-acquired, leading to the misclassification of silent transmissions over several years, as observed in our case. In this context, it is particularly important that medical institutions that conduct such detailed epidemiological analyses are not blamed for the discovery of transmission events that would have gone unnoticed in institutions without targeted molecular and epidemiological analyses.
Methods
The investigation was conducted from July 2018 to August 2023 in a tertiary care hospital in northern Germany, encompassing all medical specialities excluding paediatrics. During this period, all P. aeruginosa clinical isolates of adult wards recovered from routine diagnostic except screening material were collected. Isolates exhibiting phenotypic resistance to piperacillin, ceftazidime, ciprofloxacin, meropenem and imipenem based on the official multidrug-resistant (MDR) definition by the German healthcare authorities underwent further molecular sequencing.
For DNA extraction, the QIASymphony SP Instrument and QIAsymphony DSP Virus/Pathogen Mini Kit (Qiagen, Venlo, The Netherlands) were used as recently published [26]. Sequencing was performed using the NextSeq500 platform (Illumina, San Diego, USA), in combination with the NEBNext Ultra DNA Library Prep Kit and NEBNext Multiplex Oligos for Illumina (NEB, Ipswich, USA). Assembly of genomes and cgMLST was conducted with the Seqsphere software package (Ridom, Münster, Germany Version 11.0.0; integrated Velvet assembler Version 1.1.04), applying a clustering threshold of ≤ 11 allelic differences, based on literature review [14, 21]. A total of 4478 core genes were used for cgMLST. Detailed information about the assembled genomes is given in the Supplementary Table (blue section). The assembled genomes of the six patients, the non-related patients and environmental specimen have been deposited to the public database (NCBI GeneBank database) under the BioProject no. PRNJA1288733. Resistance gene analysis was conducted using the AMRFinder (Version 1.3.1) integrated in the Seqsphere package (all identified genes or mutations displayed in the Supplementary Table, orange section). Additionally, screening for plasmid replicon sequences was performed using PlasmidFinder 2.1 [27]. Large contigs containing blaVIM−2 were analysed for homologies to known sequences using nucleotide BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi). After identifying an In59 integron (accession number AF263519.1) as closely related, an alignment of all contigs with a length of more than 10,000 base pairs containing blaVIM−2 and/or qacE∆1 in comparison to the In59 integron and the blaVIM−2 gene alone (Supplementary Fig. 2) was generated using MAUVE (version 20150226) and the progressiveMauve alginment [28].
Following epidemiological indications as aforementioned, we conducted an environmental sampling in the ICU ward with association to the ST111 cluster. The environmental sampling was initiated in November 2023 with a focus on sinks and rinse water. Sinks in six bathrooms and surrounding of the washbasin across seven rooms and the unclean work space were sampled. A total of 36 specimen were collected in two rounds of environmental sampling.
For sampling, swabs and moistened sponge were utilised for siphons and environmental surroundings, respectively, transported to the microbiology laboratory, and processed within 12 h. Swabs were vortexed and subcultured onto blood agar, MacConkey agar, and ESBL agar (bioMérieux). Sponges were packed in sterilised boxes and were incubated in Tryptic Soy Broth (TSB) at 37 °C for 24 h. Afterwards 10 µl were subcultured on ESBL agar and incubated at 36 °C under aerobic conditions for 24–48 h. Colonies consistent with P. aeruginosa subcultured from swabs and sponges were identified via MALDI-TOF MS, and antimicrobial susceptibility testing was performed with VITEK2. If phenotypic MDR P. aeruginosa was detected, aforementioned sequencing protocol was applied to the environmental isolates.
Sink-to-patient transmission was defined as hospital-acquired colonisation (detected in rectal swabs, throat swabs, clinical specimen without initiating antibiotic treatment) or infection (clinical specimen with initiation of antibiotic treatment) with P. aeruginosa ST111 blaVIM−2 with a distance threshold of ≤ 11 alleles to ST111 blaVIM−2 isolates previously recovered from the environment in a room occupied by the respective patient for at least 72 h.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
Supplementary Material 2
Supplementary Material 3
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gleyce Hellen de Almeida de Souza, Rossato L, Brito GT. Graciela mendonça Dos Santos bet, Simone simionatto. Carbapenem-resistant Pseudomonas aeruginosa strains: a worrying health problem in intensive care units. Rev Inst Med Trop Sao Paulo. Sep 2021;27:63–71.10.1590/S 1678-9946202163071 PMC 849449234586305 · doi ↗ · pubmed ↗
- 2Oliver et al. The increasing threat of Pseudomonas aeruginosa high-risk clones. Drug Resist Updates July-August 2015, pp. 41–59.10.1016/j.drup.2015.08.00226304792 · doi ↗ · pubmed ↗
- 3Pirzadian J, ‘t Voor. Holt AF, Hossain M, Klaassen CHW, de Goeij I, Koene HHHT, Bode LGM, Vos MC, Severin JA. Limiting spread of VIM-positive Pseudomonas aeruginosa from colonized sink drains in a tertiary care hospital: a before-and-after study. P Lo S One. 2023;18(3):e 0283346.10.1371/journal.pone.0282090 PMC 1003824236961784 · doi ↗ · pubmed ↗