Inactivation of the porB gene reduces the virulence of Neisseria meningitidis in transgenic mice
Cecilia Klanger, Ala-Eddine Deghmane, Lorraine Eriksson, Olof Säll, Sara Thulin Hedberg, Paula Mölling, Muhamed-Kheir Taha

TL;DR
This study shows that inactivating the porB gene in Neisseria meningitidis reduces its ability to cause severe infections in mice, especially in certain strains.
Contribution
The study demonstrates that porB gene variants influence virulence in meningococcal isolates in a transgenic mouse model.
Findings
Invasive isolates caused more severe infections than carriage isolates in transgenic mice.
Carriage isolates with porB class 3 were less virulent than invasive isolates with porB class 2 or 3.
PorB- mutants caused milder infections than their parental isolates.
Abstract
Neisseria meningitidis is a human pathogen, carried asymptomatically in the nasopharynx, that can also cause invasive meningococcal disease. Understanding the carriage/invasiveness balance is crucial, and bacterial genetic factors may impact this balance. A previous genome-wide association study reported that the gene porB class 3 was significantly associated with carriage isolates. This study aimed to examine the impact of porB variants on virulence in carriage and invasive meningococcal isolates. For this, 24 isolates were used (13 invasive and 11 carriage) belonging to different genogroups (B, C, W, Y, and cnl) and selected based on the presence of the genetic variant porB class 2 or class 3. Transgenic BALB/c mice expressing human transferrin were infected intraperitoneally with these isolates. After 3 and 24 h of infection, clinical scores (fur quality, strength, and temperature)…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
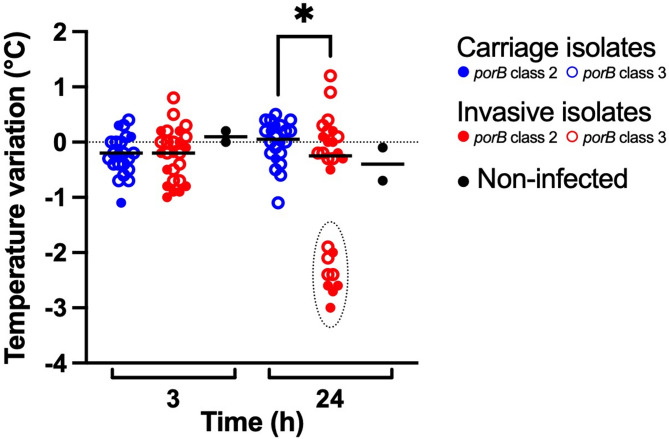 Figure 1
Figure 1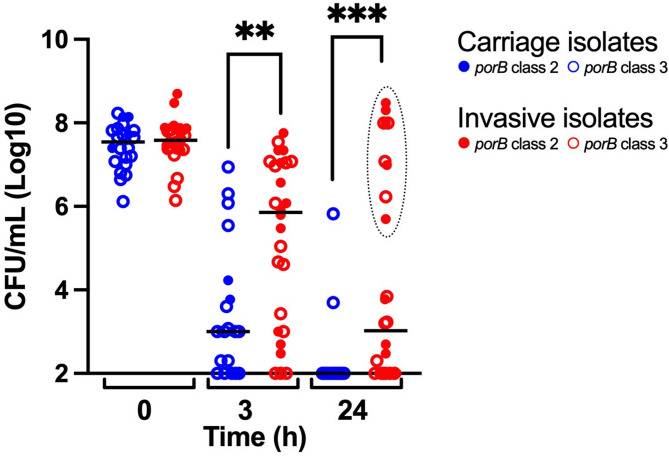 Figure 2
Figure 2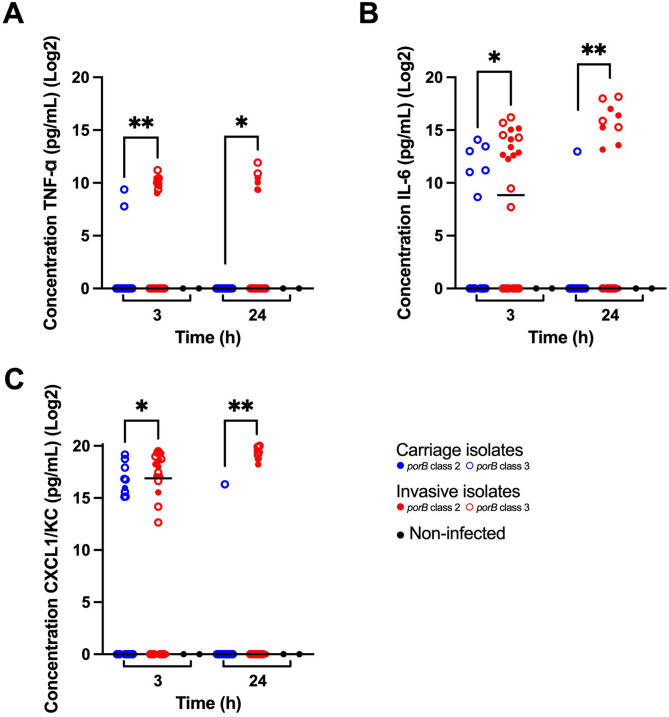 Figure 3
Figure 3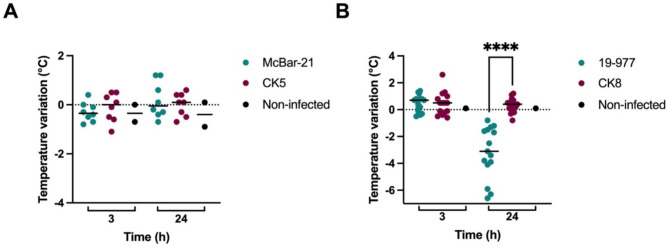 Figure 4
Figure 4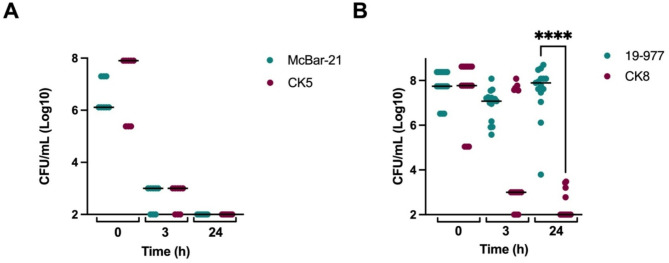 Figure 5
Figure 5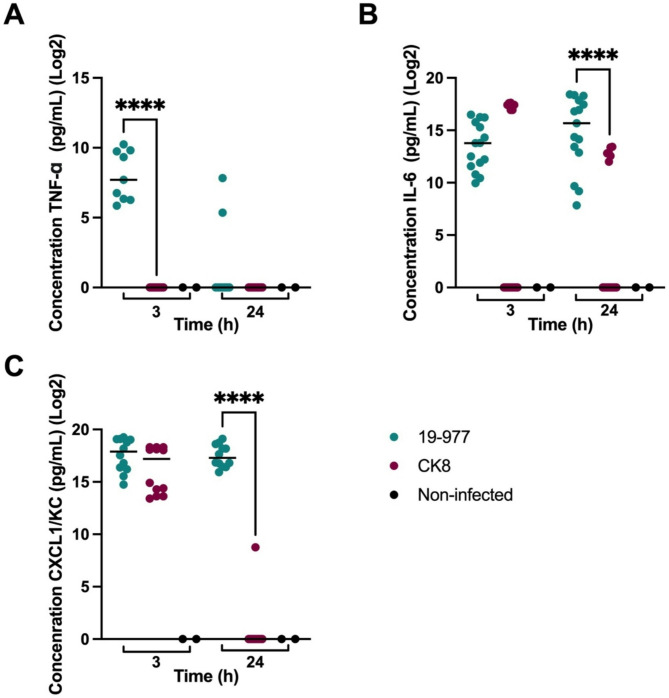 Figure 6
Figure 6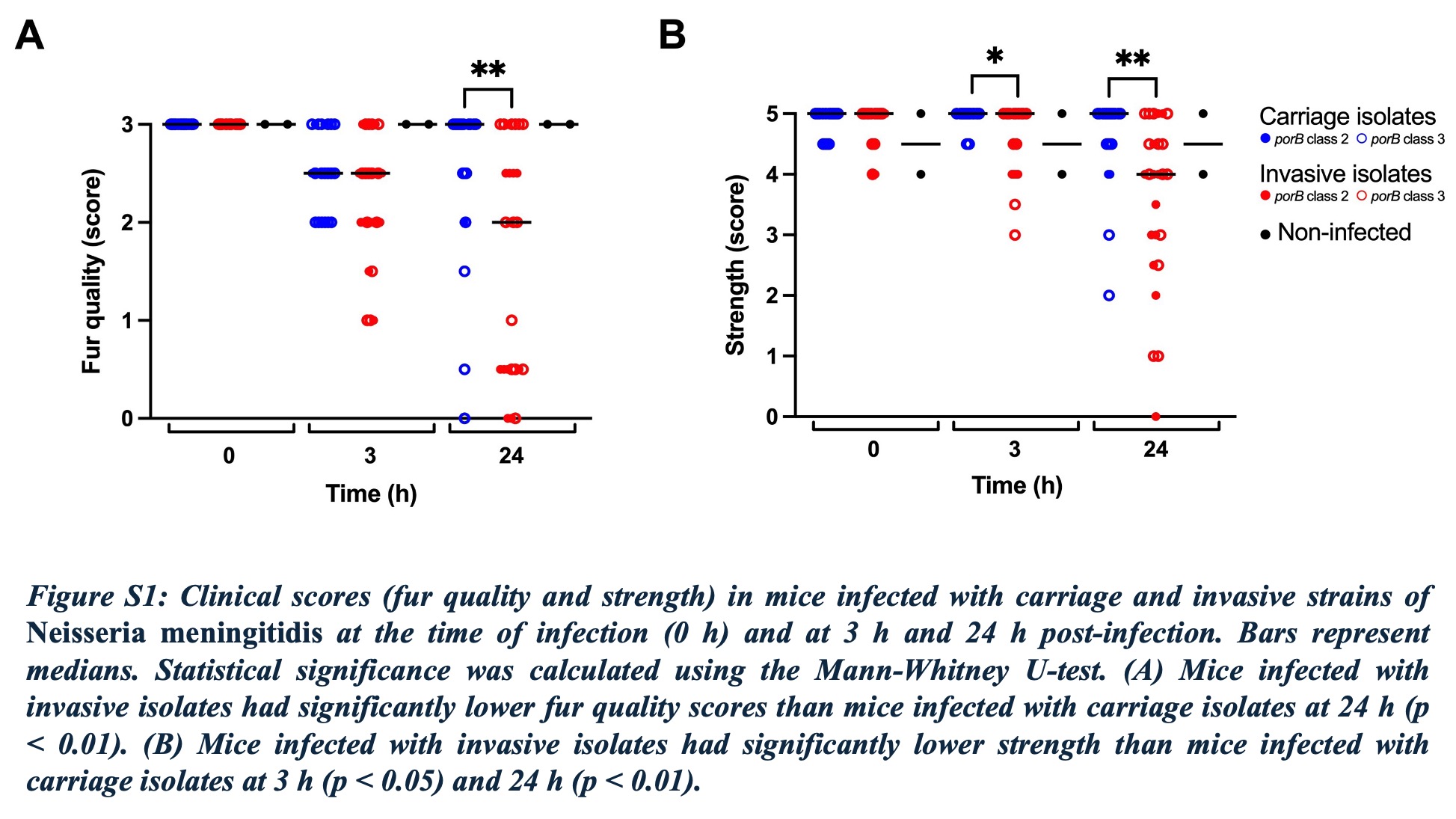 Figure 7
Figure 7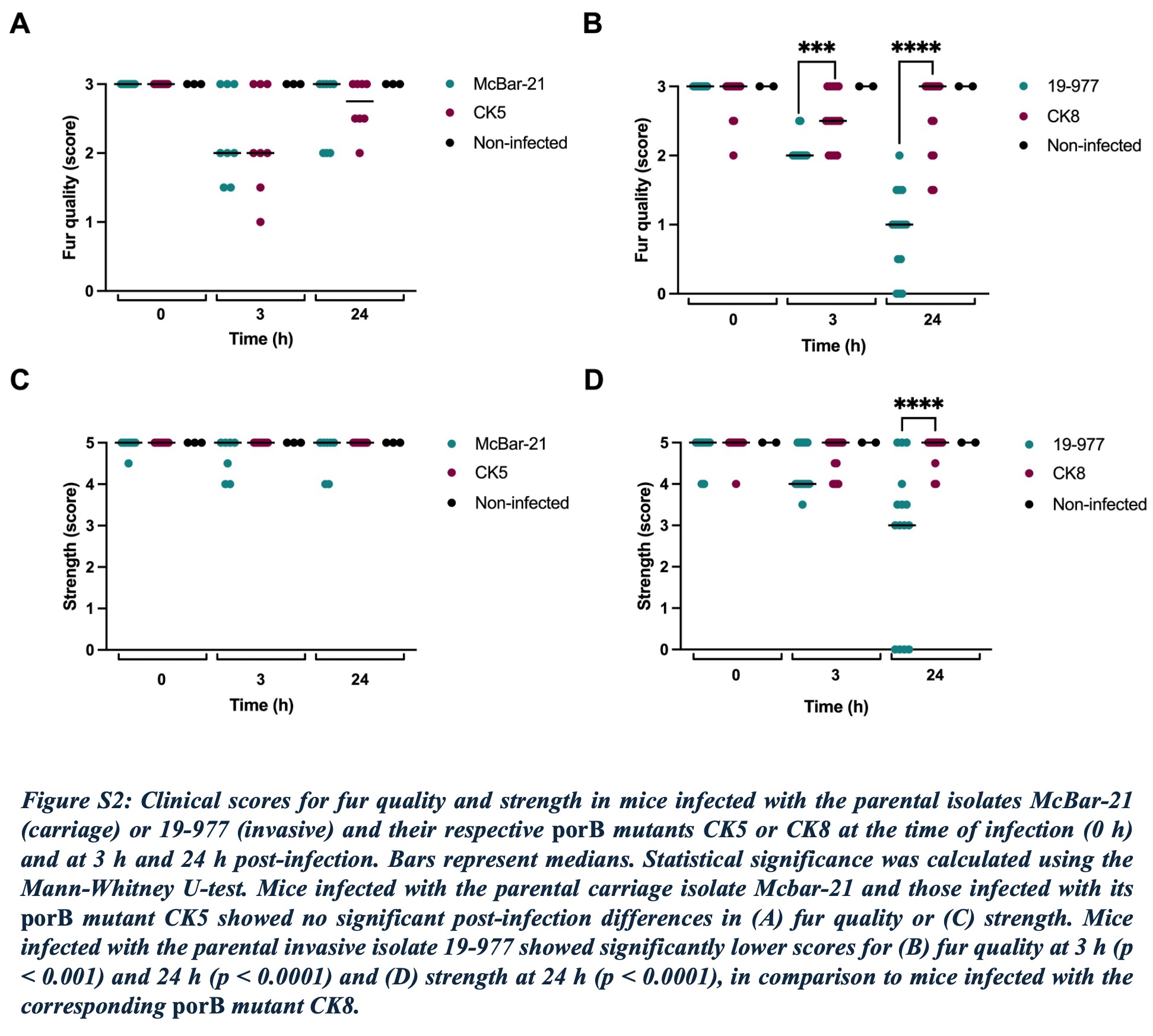 Figure 8
Figure 8- —Örebro University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Infections and Vaccines · Pneumonia and Respiratory Infections · Amoebic Infections and Treatments
Background
Neisseria meningitidis, also known as meningococcus, is a human pathogen that colonises the nasopharynx and can cause invasive meningococcal disease (IMD) including septicaemia and meningitis [1, 2]. IMD is rare, but has a high burden of disease. It is still unclear how bacteria and host interaction can cause a switch from asymptomatic carriage to IMD. The incidence of IMD varies widely across the world and by age. Fatality rates range from 2 to 20% with correct treatment, but can reach 80% in untreated cases. Up to 25% of survivors are left with lifelong disabling sequelae [3, 4]. Meningococcal carriage rates vary by region and age, with the highest rates (10–35%) seen among young adults [5, 6]. Among twelve defined serogroups, six major serogroups (A, B, C, Y, W, and X) are the leading cause of meningococcal disease globally. Non-capsulated isolates, known as capsule null locus (cnl), are generally less invasive among immunocompetent individuals but are common in carriers [2]. Meningococci can also be classified by multilocus sequence typing into sequence types (ST) and clonal complexes (cc) [7], with cc11, cc32, cc269, and cc41/44 being hyperinvasive and over-represented in invasive cases [5, 8, 9].
The pathogenesis of IMD results from an interaction of host factors and susceptibility, environmental factors such as exposure to the wind-driven dust of the Harmattan, and microbial factors [2, 10]. Host factors include complement component deficiencies or treatment with complement inhibitors, asplenia, HIV infection, other types of immunosuppression, and smoking [11, 12]. Microbial virulence factors include the polysaccharide capsule, the outer membrane proteins (OMPs, e.g. PorA and PorB), adhesion molecules (pili and opacity proteins; Opa and Opc), the iron acquisition mechanisms, and the endotoxin lipooligosaccharide (LOS) [2, 13]. Although several virulence factors of N. meningitidis are known, with the most important being the capsule expression [14], it is largely unknown which genetic elements are associated with invasive disease [5], and thus why some isolates lead to carriage while others cause severe infection.
In our previous genome-wide association study (GWAS), we analysed carriage and invasive N. meningitidis isolates [15]. The study identified single nucleotide polymorphisms (SNPs) and genes associated with isolates of either carriage or invasive type. Among these genes was porB class 3, associated with carriage isolates. The porB gene encodes for porin B (PorB), a major OMP. This voltage-gated porin is a known virulence factor for Neisseria species, and is involved in host-cell interaction [2, 16]. In any given strain of N. meningitidis, PorB is encoded by one of two homolog families, to which the allele of the porB locus is assigned: porB class 2 and porB class 3 [17].
In the present study, we built on our previous findings by using an animal model with transgenic BALB/c mice expressing human transferrin [18, 19]. The aim was to examine the virulence of carriage and invasive isolates of N. meningitidis, focusing on genetic variants of porB and isolates with inactivated porB, in humanized mice.
Methods
Bacterial isolates
Bacterial isolates used in this study were chosen from the earlier GWAS study (Table 1) [15]. The carriage isolates (n = 11) were collected during a carriage study among Swedish university students [20], and the invasive isolates (n = 13) were collected in Sweden during the same years, 2018 and 2019. These 24 isolates belonged to different genogroups (B, C, W, Y, and cnl) and were selected to ensure overall genetic similarity between the isolates, with the primary difference being the presence of either porB class 2 or class 3. Capsule expression of the isolates was determined using co-agglutination (Difco antisera, BD Diagnostics), and was categorised as positive (+), negative (-), or, if the result was uncertain, as positive/negative (+/-). The strength of the agglutination reaction was assessed qualitatively, and isolates marked as +/– showed weaker reactions compared to strongly positive (+) isolates, but not absent reactions as in the negative isolates. Isolates were grown on gonococcal agar plates (3.6% Difco GC Medium Base agar, BD Diagnostics) before co-agglutination, and on agar plates containing GCB medium (Difco, Thermo Fisher Scientific) supplemented with Kellogg’s supplements I and II [21] for the other experiments. The isolates were incubated on the agar plates overnight, for 18–20 h, in a humid atmosphere with 5% CO_2_ at 37° C.Table 1 Basic characteristics of* Neisseria meningitidis isolates included in this study (n = 26). ST = sequence type; cnl = capsule null locusIsolateGenogroupClinicalcharacteristicClonal complexporB* classCapsuleexpression ^a^Origin19 − 3YInvasiveST-232+[15]19–192YInvasiveST-232+[15]McBar-1234YCarriageST-233+[20]McBar-1425YCarriageST-233-[20]19–926YInvasiveST-233+[15]19–978YInvasiveST-233+[15]McBar-21YCarriageST-232+[20]18–122WInvasiveST-112+[15]18–391WInvasiveST-112+[15]18–62WInvasiveST-112+[15]McBar-1399WCarriageST-112+/-[20]19–419CInvasiveST-323+[15]19–977CInvasiveST-323+[15]McBar-1859CCarriageST-323-[20]McBar-2341CCarriageST-323+[20]McBar-621BCarriageST-323-[20]19–853BInvasiveST-322+[15]19–30BInvasiveST-41/443+/-[15]McBar-1351BCarriageST-41/443-[20]McBar-1820BCarriageST-41/443-[20]McBar-1226cnlCarriageST-41/443Not applicable[20]18–641cnlInvasiveST-41/443Not applicable[15]McBar-1322BCarriageST-353+[20]19-708-1BInvasiveST-353+[15]CK5 ^b^YCarriageST-23Notapplicable+This studyCK8^b^CInvasiveST-32Notapplicable+This study^a^ Level of capsule as determined by co-agglutination, expressed as positive (+), negative (-), or uncertain (+/-)b porB mutant harbouring an inactivated porB gene. CK 5 was derived from McBar-21,and CK8 was derived from 19 to 977
Construction of porB mutants
Isolates were selected for creation of porB mutants based on the carriage and invasive potential observed in mice and the demonstrated capsule expression: the carriage isolate McBar-21 (mutant CK5) and the invasive isolate 19–977 (mutant CK8) (Table 1). The porB mutants were created by transformation of the previously described plasmid pGEM-porB::erm [22]. Transformants were selected on GCB agar plates supplemented with erythromycin at 2 µg/mL. Inactivation of porB was verified by PCR, using primers porB1F and porB100R and ERAM-1 and ERAM-3 (Supplementary Material Table S1) [22], and analysed by gel electrophoresis.
Experimental infection in mice
The congenic BALB/c transgenic mice expressing human transferrin used in this study were established in the laboratory at the Institut Pasteur (using BALB/c mice from Janvier Labs, France) as previously described [19]. The mice were bred in-house and kept in a biosafety containment facility in filter-topped cages with sterile litter, water, and food, according to institutional guidelines. Female mice aged 8–12 weeks were used. The mice were injected intraperitoneally with 0.5 mL bacterial suspension of 5 × 10^7^ colony-forming units (CFU)/mL. Since the injection was considered a low-pain procedure, no anaesthesia was used. The carriage (n = 11) and invasive (n = 13) N. meningitidis isolates (Table 1) were tested in mice to assess their effects on clinical and biological markers of infection. Each mouse was injected with bacterial suspension of one isolate. All isolates were run in duplicate, using two separate experiments. NaCl (0.9%) was injected as negative control. Additional infection experiments were also performed for two of the isolates, one carrier and one invasive isolate, and their respective porB mutants (Table 1). The carriage isolate and its mutant were tested in two separate experiments with 3 and 5 mice per isolate, respectively (8 mice in total per isolate). The invasive isolate and its mutant were tested in three separate experiments with 3, 6, and 6 mice per isolate, respectively (15 mice in total per isolate). For all experiments, a total of 100 mice were used.
To determine the severity of infection with N. meningitidis in mice, the clinical outcomes of infection were assessed before infection (0 h) and at 3 and at 24 h post-infection, by scoring the quality of the fur, the strength, and the temperature of the mouse. Clinical observations were performed by the same assessor using standardised criteria and were not blinded to isolate type. Efforts were made to minimise potential bias through consistent evaluation. Hypothermia has been noted as a symptom of severe meningococcal infection in mice [23]. Fur quality was scored from 0 to 3, with 0 indicating erected and ruffled fur and 3 indicating glossy and smooth fur. Strength was scored from 0 to 5 by measuring the number of metallic chains a mouse could lift. Transcutaneous temperature was measured using an infrared thermometer for rodents (Bioseb, Vitrolles, France). The initial bacterial inoculum was determined, as well as bacterial loads (CFU/mL) in blood. Blood was drawn retro-orbitally at 3 h and 24 h post-infection. Serial dilutions were plated onto GCB plates that were incubated overnight at 37 °C in 5% CO_2_, and the bacterial count was then determined.
The cytokines interleukin-6 (IL-6), tumour necrosis factor α (TNF-α), and CXC chemokine ligand 1/keratinocyte-derived cytokine (CXCL1/KC) in sera from the 3 h and 24 h blood draws were measured using enzyme-linked immunosorbent assay (ELISA). Samples of sera were diluted 10 to 1 000 times, depending on the immunoassays for different cytokines. Kits for IL-6, TNF-α, and CXCL1/KC (R&D Systems) were used according to their respective protocols.
Animals were euthanised by cervical dislocation, in accordance with institutional and national ethical guidelines, and in compliance with the AVMA Guidelines for the Euthanasia of Animals (2020) [24]. The procedure was performed by personnel with demonstrated high technical proficiency and was selected for its ability to ensure rapid, humane death in small rodents. When necessary, topical ophthalmic anaesthetic (proparacaine 0.5% with heparin 5 U/mL) was applied. All efforts were made to ensure the animals were treated with care and euthanised as humanely as possible.
Statistical measures
Clinical infection outcomes were expressed as medians. The Mann-Whitney U-test was used to compare the outcome of infection between mice infected with the carriage and invasive isolates, between isolates of porB class 2 and 3, and between the parental isolates with their respective mutants. The data for IL-6 and CXCL1 were log2 transformed after adding an arbitrary value of 1 to all samples. A p-value of < 0.05 was considered significant. All statistics were calculated using version 10.4.0 of GraphPad Prism (GraphPad Software).
Results
Evaluation of infection outcomes in mice
Data on the co-agglutination tests for the capsule expression of the 24 bacterial isolates are presented in Table 1. Of the 24 isolates, 15 were positive by co-agglutination, five were negative (all carriage isolates), and two were uncertain. Two isolates were cnl.
All 24 isolates were tested in transgenic mice to assess their effects on clinical and biological markers of infection. After infection with the original isolates, the mice infected with carriage versus invasive isolates showed significant differences in clinical outcomes, bacterial loads, and cytokine levels. Mice infected with invasive isolates were more ill at 24 h post-infection, as indicated by significantly lower temperature (Fig. 1) and scores for fur and strength (Supplementary Material Figure S1). This observation was corroborated by significantly higher bacterial loads in blood from mice infected with invasive isolates compared to carriage isolates (Fig. 2) at 3 h and 24 h post-infection. Post-infection cytokine levels were also significantly higher in mice infected with invasive isolates. For TNF-α, the difference was more significant at 3 h post-infection (Fig. 3A). For IL-6 (Fig. 3B) and CXCL1/KC (Fig. 3C), the difference was more significant at 24 h post-infection. The variation of all scores within the groups was greater in the mice infected with invasive isolates, compared to mice infected with the carriage isolates.
Further analyses showed no significant differences in the outcomes of infection between porB class 2 or 3, or between invasive isolates of different porB class (Figs. 1, 2 and 3). Carriage isolates of porB class 3 were significantly less virulent than both invasive isolates of porB class 3 at 24 h (p 0.0043) and invasive isolates of porB class 2 at 3 h (p 0.0058) and 24 h (p 0.0010). However, carriage isolates of porB class 2 did not show lower virulence than invasive isolates of either porB class 2 or porB class 3.
Two groups were identified among the mice infected with invasive isolates: one infected with isolates that showed high virulence, and one infected with isolates that showed lower virulence. This was most evident at 24 h post-infection. The isolates that showed high virulence all belonged to serogroups C and W (Figs. 1, 2 and 3) but were of different porB class (Table 1).
Fig. 1. Temperature variation in mice infected with carriage and invasive isolates of Neisseria meningitidis, at 3 h and 24 h post-infection from each experiment. Non-infected mice are also included. The bars represent medians. Statistical significance was calculated using the Mann-Whitney U-test. At 24 h,the mice infected with invasive isolates showed significantly lower temperature than the mice infected with carriage isolates (p < 0.05). The dotted oval at 24 h highlights isolates of serogroups C and W. Data on capsule expression are shown in Table 1
Fig. 2. Bacterial load in mice infected with carriage and invasive isolates of Neisseria meningitidis at the time of infection (0 h; representing the inoculum) and at 3 h and 24 h post-infection from each experiment. The bacterial load at 0 h reflects the initial concentration of the bacterial suspension used for intraperitoneal injection and not the bacterial load in blood. Bars represent medians. Statistical significance was calculated using the Mann-Whitney U-test. The mice infected with invasive isolates showed a significantly higher number of bacteria than the mice infected with carriage isolates at both 3 h (p < 0.01) and 24 h (p < 0.001). The dotted oval at 24 h highlights isolates of serogroups C and W. Data on capsule expression are shown in Table 1
Fig. 3. Concentrations of cytokines 3 h and 24 h post-infection in mice infected with carriage and invasive isolates of Neisseria meningitidis, and in non-infected mice from each experiment. Results are expressed as medians. Statistical significance was calculated using the Mann-Whitney U-test. (A) TNF-α levels were significantly higher in the mice infected with invasive isolates at 3 h (p < 0.01) and 24 h (*p *< 0.05). (B) IL-6 levels were significantly higher in the mice infected with the invasive isolates at 3 h (p < 0.05) and 24 h (p < 0.01). (C) CXCL1/KC levels were significantly higher in the mice infected with the invasive isolates at 3 h (p < 0.05) and 24 h (p < 0.01)
Infection in mice with porB mutants
Mice infected with the carriage isolate McBar-21 and its mutant CK5 (harbouring an inactivated porB gene) showed no significant differences in post-infection clinical scores (Fig. 4A and Supplementary Material Figure S2). Mice infected with the invasive isolate 19–977 had a significantly lower temperature 24 h post-infection in comparison to those infected with its mutant CK8 (Fig. 4B). Other clinical markers such as fur quality and strength reflected the same pattern (Supplementary Material Figure S2). Moreover, the mice infected with 19–977 showed a significantly higher bacterial load than their counterparts infected with CK8 (Fig. 5B). The mice infected with this invasive mutant, harbouring an inactivated porB gene, seem to have cleared the bacteria to a similar extent as the mice infected with the carriage isolates (Fig. 5A). The cytokine levels followed the same pattern, being significantly higher in the mice infected with 19–977 than in those infected with CK8 (Fig. 6). For TNF-α, the difference was more significant at 3 h post-infection (Fig. 6B), while for IL-6 (Fig. 6B) and CXC1/KC (Fig. 6C), the difference was more significant at 24 h post-infection.
Fig. 4. Temperature variation in mice infected with the parental isolates McBar-21 (carriage) or 19–977 (invasive) and their respective porB mutants CK5 and CK8,at 3 h and 24 h post-infection. Bars represent medians. Statistical significance was calculated using the Mann-Whitney U-test. Non-infected mice are also included. (A) Mice infected with the parental carriage isolate McBar-21 and those infected with its porB mutant CK5 showed no significant differences. (B) Mice infected with the parental invasive isolate 19–977 showed a significantly lower temperature than those infected with its porBmutant CK8 at 24 h (p < 0.0001)
Fig. 5. Bacterial load in mice infected with the parental isolates McBar-21 (carriage) or 19–977 (invasive) and their respective porBmutants CK5 and CK8, at the time of infection (0 h; representing the inoculum) and at 3 h and 24 h post-infection. The bacterial load at 0 h reflects the initial concentration of the bacterial suspension used for intraperitoneal injection and not the bacterial load in blood. Bars represent medians. Statistical significance was calculated using the Mann-Whitney U-test. (A) Mice infected with the parental carriage isolate Mcbar-21 and those infected with its porB mutant CK5 showed no significant differences. (B) Mice infected with the parental invasive isolate 19–977 showed a significantly higher number of bacteria than those infected with its porB mutant CK8 at 24 h (p < 0.0001).
Fig. 6. Concentrations of cytokines 3 h and 24 h post-infection in mice infected with the parental invasive isolate 19–977 and its porB mutant CK8 and in non-infected mice from each experiment. Bars represent medians. Statistical significance was calculated using the Mann-Whitney U-test. (A) TNF-α levels were significantly higher in the mice infected with the parental isolate at 3 h (p < 0.0001), with no significant difference at 24 h. (B) IL-6 levels were significantly higher in the mice infected with the parental isolate at 24 h (p < 0.0001),with no significant difference at 3 h. (C) CXCL1/KC levels were significantly higher in the mice infected with the parental isolate at 24 h (p < 0.0001), with no significant difference at 3 h
Discussion
The use of this mouse model allowed a comparison of the virulence of invasive and carriage isolates of N. meningitidis. The invasive isolates were more virulent in mice, emphasizing intrinsic properties of the bacteria. The mouse model provided both clinical scores and biological scores using bacterial loads and inflammatory cytokines in the blood. There were some differences regarding porB class in these isolates; carriage isolates of porB class 3 were less virulent than invasive isolates of either class 2 or 3, but carriage isolates of porB class 2 showed no such difference. The invasive isolate that was mutated with inactivated porB was less virulent than its parental isolate, highlighting the importance of porB as a virulence factor.
The mice infected with the carriage isolates generally had a milder infection than the mice infected with invasive isolates, as revealed by several parameters. For instance, those with the invasive isolates had a significantly lower temperature at 24 h. This reflects a more severe infection in these mice, as hypothermia may be a result of severe meningococcal disease in mice [23]. Cytokine levels are generally correlated to the severity of IMD and fatality. Among these, the pro-inflammatory cytokines IL-6, which stimulates acute phase responses, haematopoiesis, and immune reactions [25], and TNF-α, a central early immune response activator [26], have both been related to meningococcal disease severity [27, 28]. The chemokine CXCL1/KC acts by recruiting and activating neutrophils as a host immune response to infection [29, 30], and has been linked to meningococcal infection in this mouse model [23]. TNF-α being an early immune response activator may explain why the difference was more significant at 3 h post-infection than at 24 h in all experiments where a rise in TNF-α levels was observed.
No differences in virulence were found when comparing isolates of porB class 2 with isolates porB class 3 on their own. However, when comparing carriage isolates of porB class 3 with invasive isolates of porB class 2, the carriage isolates caused significantly lower level of bacteriaemia at both 3 and 24 h, and when comparing them to invasive isolates of porB class 3, they were also significantly less virulent at 24 h. Such a difference was not observed when comparing carriage isolates of class 2 with invasive isolates of either porB class. These results are in line with the previous GWAS, which found a significant association between carriage isolates and porB class 3 [15].
The mouse model also made it possible to examine differences between invasive isolates, revealing that not all of the invasive isolates were equally virulent. There seemed to be two groups: one group of highly invasive serogroup C (ST-32 clonal complex) and serogroup W (ST-11 clonal complex) isolates, and one group of less invasive isolates. This points to the role of serogroups and clonal complexes. The evidently more invasive isolates in this study were exclusively of serogroups C and W, while the less invasive isolates were mainly of other serogroups (B, Y, cnl). Similarly, a previous study comparing differences in virulence between serogroups Y and W found that in mice expressing human transferrin, meningococci of serogroup W were more virulent than those of serogroup Y [31].
The C and the W isolates in our study belonged to the ST-32 clonal complex and ST-11 clonal complex respectively, which are both considered to be hyperinvasive [8]. Meanwhile, the Y isolates belonged to the ST-23 clonal complex, and the B isolates, though mainly of the hyperinvasive ST-41/44 clonal complex, showed weaker capsule expression based on the co-agglutination results, with several isolates marked as negative or +/-. These factors should all be taken into consideration when interpreting the results, since they might at least partly explain the outcome. Notably, these highly invasive isolates were of different porB classes; the W isolates were of porB class 2 and the C isolates were of porB class 3, and, as mentioned, no association was found between highly virulent isolates and porB class.
The results from the experiments with the porB mutants suggest that the porB gene itself plays a critical role for infection, as the mice infected with the mutants that had inactivated porB were less ill than the mice infected with the parental invasive isolates. This was reflected by higher clinical scores, significantly lower bacteraemia, and a lower inflammatory response. PorB is an outer membrane porin which plays a role in microbial virulence. The protein can insert itself into mitochondrial membranes, which induces cell apoptosis and enhances invasiveness through induction of Ca2 + influx and activation of toll-like receptors [2, 16]. This may explain why the mutated meningococci without the porB gene showed less virulence.
The invasive isolate 19–977 that was mutated was of porB class 3, and the carriage isolate Mcbar-21 was of porB class 2. It is still unclear whether the porB class itself is important for meningococcal virulence, and not just the presence of the gene. In the GWAS, porB class 3 was linked with carriage isolates [15]. Interestingly, in the previously mentioned mouse trial comparing W and Y strains, the W isolates were all of porB class 2 [31]. Furthermore, previous studies with transgenic rats have shown that isolates expressing porB class 2 exhibit higher virulence than those expressing porB class 3. This suggests that porB class 2 may enhance immune evasion capabilities [32, 33].
One study investigated strains of meningococci that were engineered to lack certain molecules known to inhibit the alternative pathway of the immune system, and found that some strains still managed to inhibit the alternative pathway. These strains expressed PorB class 2 protein, which appeared to help the bacteria resist the immune response by binding certain parts of the human complement Factor H and by reducing the deposition of the immune protein complement component 3 (C3). This was linked to resistance to the body’s complement system, which is a key part of the immune defence against infections. This resistance mechanism was specific to humans, as animal experiments showed that the bacteria did not resist the immune system in rabbits or mice [32]. These findings could potentially explain why our study could not find stronger differences regarding porB class and outcome of infection in mice. Further investigations are needed to fully address the role of porB class.
A limitation of this study is that relatively few isolates and mice were used. Experiments with more isolates and more mice could generate even more significant data, although this must be considered in relation to the ethical imperative to use as few mice as possible while still yielding reliable results. Another limitation is the use of a mouse model. Although it is possible to sustain an infection with N. meningitidis in these mice, since they have been modified to express human transferrin, it is important to remember that the meningococcus is a human pathogen. The bacteria might act somewhat differently in mice, as observed in the abovementioned study regarding porB class, which might explain why we did not find a clearer link between porB class and outcome of infection. A third limitation of the study is that only a small number of successful porB mutants with inactivated porB were used. The mutation process was challenging, and thus only two isolates were successfully mutated. Ideally, more isolates would have been mutated through porB inactivation. To increase the number of mutations in future studies, the transformation procedure could be modified– for example, by replacing natural transformation with electroporation or conjugation, which may help circumvent restriction barriers related to DNA uptake. Additionally, creating mutations in other outer membrane proteins would provide a broader context for comparison and could help describe the specific role of PorB.
In another study using GWAS on N. meningitidis, several genetic variants, including SNPs, were associated with meningococci of carriage or invasive type. These genetic variants included a significant association of invasive isolates with polymorphisms in the fHbp gene. Notably, the strongest associations were found in genes involved in producing meningococcal surface structures that interact with host cells and immune molecules, as well as in genes linked to their expression [34]. In another study examining invasive strains in New Zealand, carriage strains from household contacts were analysed simultaneously. Although the isolates were very similar according to laboratory typing methods, they did have genetic differences. Several carriage strains had also lost their type IV pili, and this was correlated with reduced TNF-α expression when cultured with epithelial cells [35]. A study of an epidemic of meningococcal outbreak in West Africa revealed that invasive disease had resulted from a previously asymptomatic carriage strain that had acquired virulence genes, including capsule genes [36]. These findings are in line with our results, and support the notion that genetic factors, in addition to capsule expression, play a major role in disease development.
Other genes found to be associated with the carriage isolates in our previous GWAS [15] were tspB, encoding for T and B cell-stimulating protein B, which mediates the binding of human IgG and the formation of biofilm and is also involved in the pathogenesis of N. meningitidis [37], and the PilE/S genes, encoding a subunit of the pili. Two SNPs were also found to be associated with invasiveness or carriage; one SNP in the gene glmU, which encodes for an enzyme involved in synthesis of a substrate involved in the synthesis of lipooligosaccharide and the polysaccharide capsule, and one SNP in the gene fkpB, which encodes an enzyme involved in protein folding. The role of these genetic variants in promoting invasive infection or severity of infection still remains to be explored phenotypically in functional experiments.
Conclusion
This study confirms that in comparison to carriage isolates, invasive isolates of N. meningitidis have a greater ability to cause an invasive infection in transgenic mice, which emphasizes the capacity of the isolate itself to cause infection. However, this increased ability was most apparent among serogroup C and W isolates, while serogroup B isolates (both carriage and invasive) were overall of lower virulence and not significantly different from each other. The results highlight the importance of the intrinsic properties and the genetic signature of the bacteria, including the impact of the porB gene, on meningococcal virulence. Variants of the porB gene could also to some extent be linked to phenotypic outcome. However, since porB class alone did not consistently explain differences in virulence, its contribution may depend on the broader genomic context of the isolate. Our data underline that in addition to host and environmental factors, bacterial factors are key determinants in the outcome of N. meningitidis-host interaction.
Supplementary Information
Supplementary Material 1. Supplementary Material 2. Supplementary Material 3.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ren X, Eccles DA, Greig GA, Clapham J, Wheeler NE, Lindgreen S, et al. Genomic, transcriptomic, and phenotypic analyses of neisseria meningitidis isolates from disease patients and their household contacts. m Systems. 2017;2. 10.1128/msystems.00127.17.10.1128/m Systems.00127-17PMC 568652129152586 · doi ↗ · pubmed ↗
- 2Pub MLST. Neisseria spp. https://pubmlst.org/organisms/neisseria-spp (accessed 5 Nov 2024).