A novel signal peptide toolbox for optimized heterologous expression of yeast pheromones in Bacillus subtilis
Tomislav Vološen, Henri Max Deda, Anica Walther, Philipp F. Popp, Thorsten Mascher, Diana Wolf

TL;DR
This paper introduces a toolbox to improve protein secretion in bacteria using yeast pheromones as test cases.
Contribution
A novel toolbox with 74 signal peptides for efficient heterologous protein secretion in Bacillus subtilis is developed.
Findings
Eight signal peptides enabled secretion of P-pheromone in B. subtilis.
Only three signal peptides effectively facilitated α-pheromone secretion.
Maximum secretion levels reached 43 nM for P-pheromone and 8 nM for α-pheromone.
Abstract
Optimising downstream processes and achieving high product yields are essential for cost-effective and scalable bioproduction, with efficient protein secretion being a critical factor in industrial applications. To address this, we leveraged the widely utilised Sec secretion system of Bacillus subtilis to enhance heterologous protein secretion. Given the lack of reliable in silico tools for predicting optimal combinations of Sec system signal peptides (SPs) with a protein of interest (POI), we aimed to optimise the secretion efficiency of SP–POI pairings by a toolbox approach. We developed an integrative evaluation vector in which the promoter, SPs, and a coding sequence (CDS) of the POI are easily exchangeable. Further, we generated a toolbox containing 74 SPs naturally present in B. subtilis that can easily be integrated into the evaluation vector and thereby fused to the POI. As…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
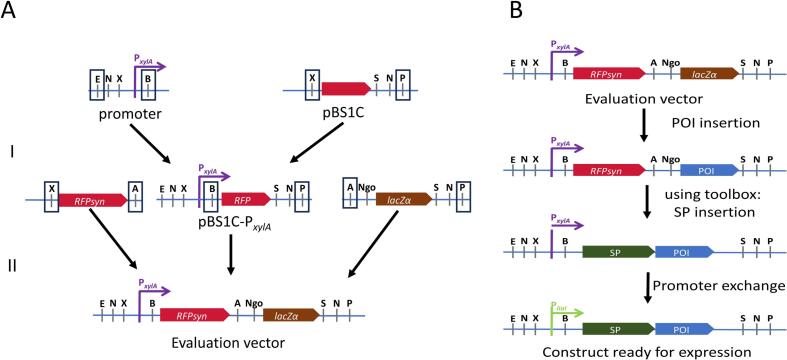 Figure 1
Figure 1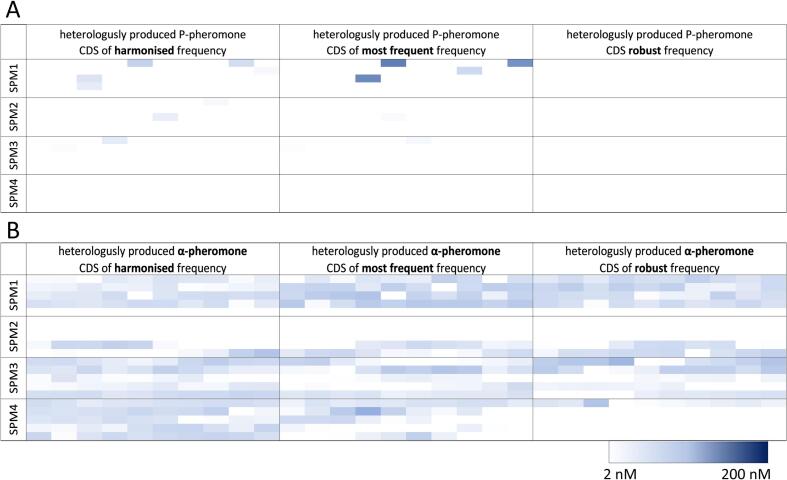 Figure 2
Figure 2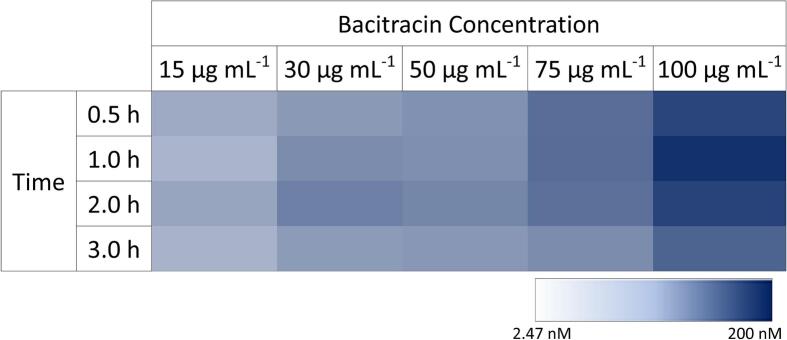 Figure 3
Figure 3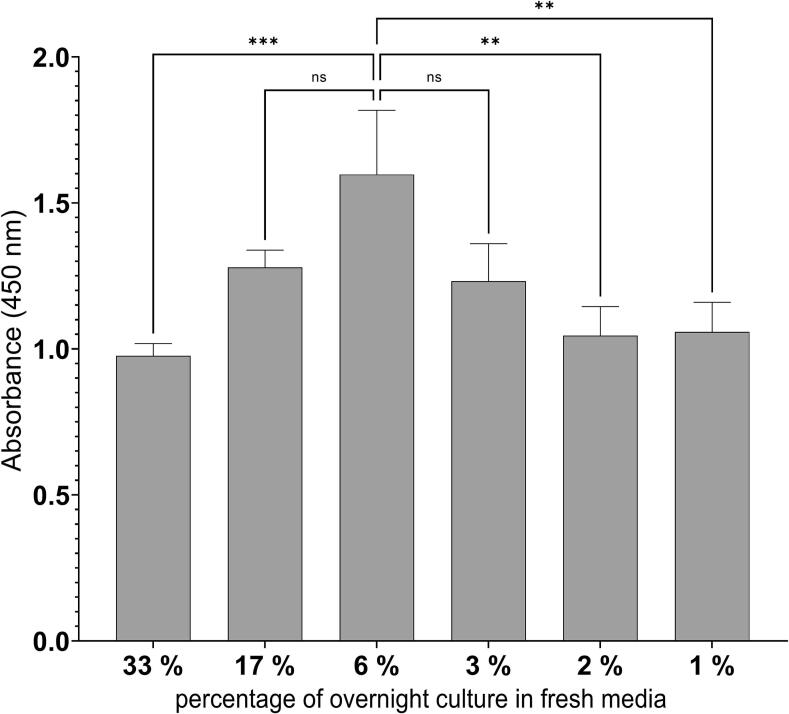 Figure 4
Figure 4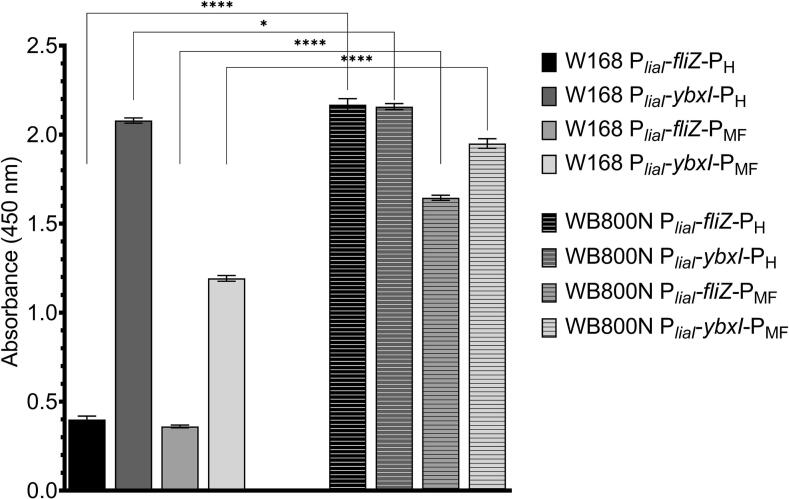 Figure 5
Figure 5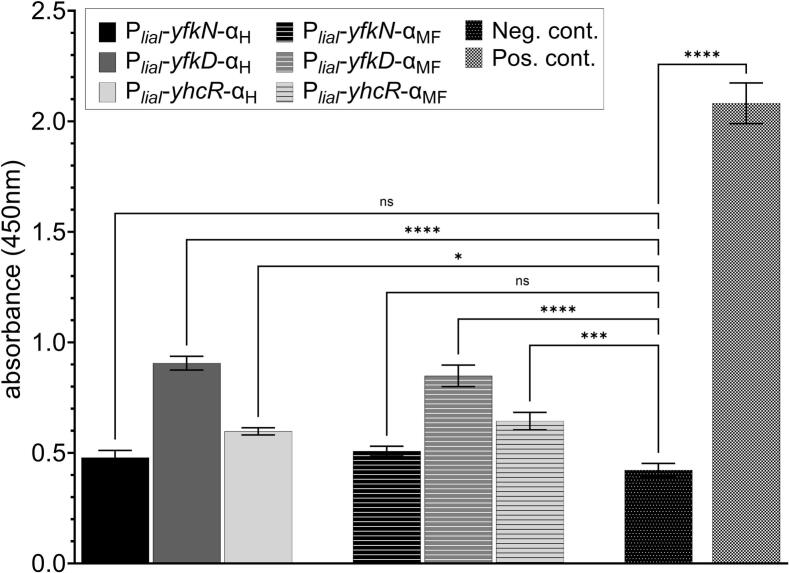 Figure 6
Figure 6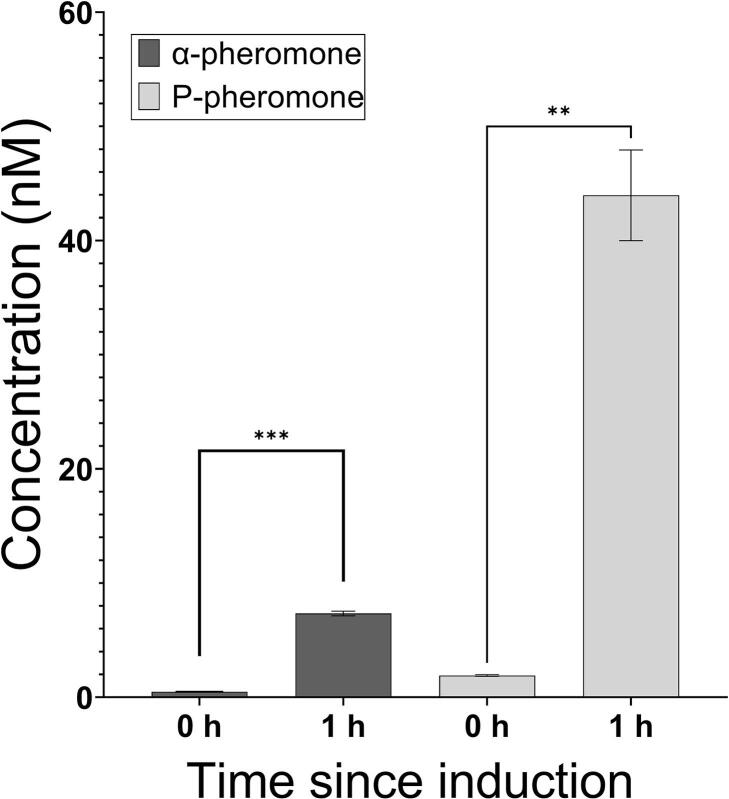 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA and protein synthesis mechanisms · Bacterial Genetics and Biotechnology · Biochemical and Structural Characterization
Introduction
1
The estimated market of technical bulk enzymes produced in industrial biotechnology doubled in the last ten years and was reaching approximately US$11.9 billion in the year 2022.1 This economic development increases the need for new technologies and platforms with enhanced speed and cost efficiencies for production. Hereby, extracellular protein secretion is a critical technology for reducing the costs of industrial downstream processing, product recovery, and energy consumption in various industrial applications.2 Not surprisingly, optimising protein secretion in industry-relevant organisms is a priority in White Biotechnology processes.3
The Gram-positive bacterium Bacillus subtilis includes the “generally recognised as safe” (GRAS) status and is widely used as the host for the production and secretion of commercial enzymes.4 In 2014, 17 % of total commercial enzymes were produced by B. subtilis.2 B. subtilis harbours four main secretion pathways, of which the Sec secretion pathway is the most prominently used.5 Proteins secreted by the Sec pathway carry a secretion-targeting peptide sequence in the N-terminus referred to as signal peptide (SP).6 Transmembrane protein translocation through the Sec-pathway in B. subtilis is controlled by at least four proteins, including the SecA translocation motor that drives preprotein through the SecY-SecG-SecE channel.7 After releasing the C-domain of the SP in the extracytoplasmic space, signal peptidase (SPase) and signal peptide peptidase (SPP) remove the SP from the secreted proteins. After full translocation of the preprotein through the cell membrane, proteins will fold into their native conformation with the help of folding catalysts (e.g., PrsA and SpoIIIJ) and quality control factors (e.g., HtrA-like proteases).8.
B. subtilis secretes over 173 different proteins through the Sec secretion system. Identifying the optimal SP is a critical factor for the efficient heterologous production and secretion of a protein of interest (POI).9, 10 Due to the limited understanding of the factors influencing protein secretion and the challenge of identifying a functional pairing with a SP, in silico prediction tools for selecting SPs to optimise protein secretion are currently lacking.11 To address this, we designed and evaluated a versatile and cost-effectively toolbox to identify the ideal SP for the secretion of any POI in B. subtilis. As proof of concept, we chose to produce α- and P-pheromone from S. cerevisiae and S. pombe in B. subtilis to challenge the usability and effectiveness of the toolbox and its included expression evaluation vector. Yeast α- and P-pheromone are 13- and 23-amino acids long peptides, respectively and are functionally essential for mating of haploid yeast cells.12, 13 Additionally, recombinant secretion of short peptides has not been documented in B. subtilis yet*.* We were interested in the optimal secretion of these peptides in order to be able to successfully design an interkingdom cell–cell communication between yeast and B. subtilis by using these pheromones as part of a larger research project. To our knowledge, heterologous production of peptides smaller than 25 amino acids and originating from eukaryotes in prokaryotes is novel and can open a wide range of applications in White Biotechnology.
Materials and methods
2
Evaluation vector construction
2.1
All vectors and oligonucleotides used to create the evaluation vector are listed in Table S1 and Table S2. The evaluation vector was developed from the pBS1C backbone previously published in.14 To use this vector as the backbone of the evaluation vector, the BsaI restriction enzyme site was removed with PCR mutagenesis using the oligonucleotides TM3163 and TM3164. Next, xylose inducible promoter PxylA was amplified with oligonucleotides iG17P051 and iG17P052 from plasmid pBS2E-PxylA (9A’s),15 digested with EcoRI-HF and BsaI (XbaI 5′ CTAG…3′ overhang), and ligated into the BsaI free variant of pBS1C vector that was previously digested with EcoRI-HF and XbaI-HF. The resulting vector pBS1C-PxylA was confirmed by sequencing. pBS1C-PxylA vector was further developed by integrating rfp and lacZα genes inside the multiple cloning sites to achieve a three-colour colony screening (red-blue-white) of transformed E. coli cells. To do so, a rfp gene was synthesised and fused to the IPTG inducible promoter PlacI (Integrated DNA Technology’s (IDT, Coralville, IA, USA) gBlocks® Gene Fragments DNA synthesis) and inserted into the storage vector pSB1C3 (parts registry: BBa_K823055iGEM) following the BioBrick™ standard. Next, the rfp gene was amplified from pSB1C3-[RFP] by using the oligonucleotides iG17P110 and iG17P159 and digested with XbaI-HF and AgeI-HF. The lacZα gene was amplified with iG17P055 and iG17P056 from DNA storage vector pSB1C3-Bba_I732902(lacZα) and digested with AgeI-HF and PstI-HF. Genes rfp and lacZα were cloned into the pBS1C-PxylA vector, digested with BsaI (XbaI 5′ CTAG…3′ overhang) and PstI, resulting in the evaluation vector (pBS1C PxylA-[PlacI-RFP-lacZα]) that was verified by sequencing. Enzymes and buffers used here to develop the evaluation vector were purchased from New England Biolabs® (NEB, Ipswich, MA, USA).
Construction of the signal peptide toolbox
2.2
In this study, a toolbox with SPs from 74 Sec-system transported proteins was established. Oligonucleotides used for SP amplification (Table S3) are designed in BioBrick™ standard and contain a ribosome binding site (RBS) and a linker previously described in.15 Amplified SPs were cloned into the storage vector pSB1C3 and transformed with E. coli DH10β. Cloning was verified by sequencing. To establish the SP toolbox, all storage vectors containing the SP fused to RBS are isolated from E. coli, aliquoted in a concentration of 0.5 ng µL^−1^, and stored at −20 °C. A list of all SPs containing storage vectors is shown in Table S4. To use the toolbox, 4 SP mixes can be prepared. Each mix contains 20 different SPs. Every SP subset/mix was amplified by using the oligonucleotides TM4487 and iG17P039 in an adjusted multi-template PCR reaction shown in Table S5. Next, amplified SP mix was digested with XbaI and AgeI, cloned into the evaluation vector carrying the POI, and transformed with E. coli. Fifty white E. coli colonies per SP subset were picked for overnight cultures and plasmid isolation. Isolated plasmids were transformed with B. subtilis. Again, fifty colonies of each SP subset were picked, and a starch assay was performed to check the correct integration of constructed plasmids into the amyE locus of the B. subtilis genome. In the next step, screening for SPs that are responsible for POI secretion was performed by indirect ELISA assay. SPs of producing cells were identified by sequencing the PCR products of the SP region.
Growth conditions
2.3
All strains used in this study are listed in Table 1. B. subtilis was grown at 37 °C with aeration in Lysogeny broth LB broth (Luria/Miller, Carl Roth) [tryptone 10 g L^-1^, yeast extract 5 g L^-1^, NaCl 10 g L^-1^] or MNGE media [88.2 % 1 × MN media (1.36 % (w/v) dipotassium phosphate x 3 H_2_O, 0.6 % (w/v) monopotassium phosphate, 0.1 % (w/v) sodium citrate·H_2_O)), 1.9 % glucose, 0.19 % potassium glutamate, 0.001 % (w/v) ammonium ferric citrate, 0.005 % (w/v) tryptophan, and 0.035 % (w/v) magnesium sulphate] or Edinburgh Minimal Medium (EMM) containing 3.75 g L^-1^ glutamic acid as the nitrogen source.16 E. coli was grown at 37 °C with aeration in Lysogeny broth (LB broth). Solid media contained 1.5 % (w/v) agar. Selection media for E. coli contained ampicillin (100 µg mL^−1^). To have a blue-red-white screening of successful E. coli transformation, agar plates contained 1 mM IPTG, 100 µg mL^−1^ X-Gal, and 100 µg mL^−1^ ampicillin. Selection media for B. subtilis contained chloramphenicol (5 µg mL^−1^) or kanamycin (10 µg mL^−1^). Genetic modification of E. coli and B. subtilis was performed as described previously.17, 18 Colonies were transferred to starch agar plates to show successful integration of the pBS1C constructs into the amyE locus of the B. subtilis genome by starch assay with Lugol’s solution (Sigma Aldrich, Germany).Table 1B. subtilis strains used in this study.StrainDescriptionSource/ReferenceB. subtilis W168WT, trpC2Laboratory stockB. subtilis WB800NnprE aprE epr bpr mpr::ble nprB::bsr Δvpr wprA::hyg (Neo^R^)19TME4009E. coli DH10β pUC57-α-pheromone_H_This studyTME4010E. coli DH10β pUC57-α-pheromone_MF_This studyTME4011E. coli DH10β pUC57-α-pheromone_R_This studyTME4012E. coli DH10β pUC57-P-pheromone_R_This studyTME4013E. coli DH10β pUC57-P-pheromone_H_This studyTME4014E. coli DH10β pUC57-P-pheromone_MF_This studyTMB6298W168 amyE::pBS1C-PliaI-fliZ-P-pheromone_MF_This studyTMB6301W168 amyE::pBS1C-PliaI-ybxI-P-pheromone_MF_This studyTMB6315W168 amyE::pBS1C-PliaI-fliZ-P-pheromone_H_This studyTMB6318W168 amyE::pBS1C-PliaI-ybxI-P-pheromone_H_This studyTMB6423WB800N amyE::pBS1C-PliaI-yhcR- P −pheromone_MF_This studyTMB6424WB800N amyE::pBS1C-PliaI-ybxI-P-pheromone_MF_This studyTMB6425WB800N amyE::pBS1C-PliaI-fliZ-P-pheromone_H_This studyTMB6426WB800N amyE::pBS1C-PliaI-ybxI-P-pheromone_H_This studyTMB6427WB800N amyE::pBS1C-PliaI-yhcR-α-pheromone_H_This studyTMB6428WB800N amyE::pBS1C-PliaI-yfkD-α-pheromone_H_This studyTMB6429WB800N amyE:: pBS1C-PliaI-yfkN- α-pheromone_H_This studyTMB6430WB800N amyE::pBS1C-PliaI-yhcR- α-pheromone_MF_This studyTMB6431WB800N amyE::pBS1C-PliaI-yfkD- α-pheromone_MF_This studyTMB6432WB800N amyE::pBS1C-PliaI-yfkN- α-pheromone_MF_This studyMF-most frequent codon usage frequency; H-harmonised codon usage frequency.
Codon adaptation
2.4
Codon sequences of α- and P-pheromones from S. cerevisiae and S. pombe were adapted for expression in B. subtilis. Codon optimisation was performed by comparing codon usage frequencies between B. subtilis and the yeasts S. cerevisiae and S. pombe. Codon usage frequencies were obtained from Codon Usage Databases.20 Three variations of codon-optimised sequences for α- and P-pheromones were designed. The first sequence contains the most frequent codon sequence that has the highest codon usage frequency in B. subtilis, which indicates the highest speed of translation in B. subtilis. The robust codon sequence has the lowest possible codon usage frequency, indicating the slowest speed of translation. Furthermore, a harmonised coding sequence was designed by comparing and matching the codon usage frequency of the S. cerevisiae and S. pombe to the codon usage frequency in B. subtilis, resulting in a translation speed similar to both yeast and bacteria. α- and P-pheromone protein sequence, codon sequence, and optimised codon sequences are shown in Fig. S1 and Fig. S2. Designed codon sequences were artificially synthesised by BioCat (Heidelberg, Germany) and donated in the pUC57 vector for storage.
Construction of B. subtilis α- and P-pheromone secretion strains
2.5
B. subtilis was genetically modified to secrete the α- and P-pheromones by using the evaluation vector and the SP toolbox. Strain construction occurred in three steps. First, restriction cloning replaced the lacZ gene in the evaluation vector with α- or P-pheromone coding sequences. To do so, the coding sequence and evaluation vector were digested with PstI and NgoMIV, ligated and transformed with E. coli. Blue-red screening of clones and sequencing of plasmid constructs confirmed the successful cloning. Second, we determined the SPs that potentially secrete α- and P-pheromone by using the SP toolbox developed in this study. Finally, we exchanged the promoter in the plasmids, including the optimal SP from PxylA to the PliaI, to ensure antibiotic inducible expression of the pheromones and circumvent leakiness of PxylA. The evaluation vector carrying PxylA, RFP gene, and α- or P-pheromone was digested with EcoRI-HF and NgoMIV-HF. The desired SP was digested with XbaI*-HF* and AgeI-HF. At the same time, the promoter PliaI was amplified with oligonucleotides TM2895 and TM2896 (Table S2) and digested with EcoRI-HF and SpeI-HF. Constructs were confirmed by sequencing and transformed with B. subtilis W168 (laboratory standard strain) and WB800N strain (an eight-protease-deficient strain).19
Heterologous expression of α- and P-pheromones in B. subtilis
2.6
Established α- and P-pheromone-producing B. subtilis strains were used to perform expression studies. Overnight cultures, including appropriate antibiotic chloramphenicol (5 µg mL^−1^) for selection, were diluted 1:100 in fresh LB or EMM media.21 200 µL of the culture dilution was aliquoted in wells of a transparent 96-well microplate (Greiner Bio-One, Kremsmünster, Austria) and cultivated at 37 °C for 3 h with agitation. Then, peptide expression was induced by adding xylose in a final concentration of 0.5 % (w/v) or 100 µg mL^−1^ of bacitracin zinc salt (Sigma), and cultivation with agitation was continued for 3 h. Expression was stopped by adding 200 nM Tris-HCl pH 8.0 buffer, 10 mM EGTA pH 8.0, and cOmplete Protease inhibitor cocktail (Merck) or Halt™ Protease-Inhibitor Cocktail (Thermo Fischer Scientific). Finally, the cultivation was inactivated by heating the 96-well plate to 80 °C for 3 min, and cells were removed by centrifugation at 1800 × g for 15 min. 100 µL of the remaining supernatant was used for an indirect ELISA assay to check and quantify the pheromone secretion.
Indirect ELISA assay
2.7
Visualisation and quantification of α- and P-pheromone concentration were conducted by a modified indirect ELISA protocol adopted from.22 In brief, 100 µL of culture supernatant was transferred to each well of an ELISA plate (High Binding 96 Chimney Well Bottom Plate, Greiner Bio-One, Kremsmünster, Austria). Additionally, 100 µL of synthetic α- or P-pheromones (Peptides & Elephants GmbH, Berlin, Germany and David’s Biotechnology GmbH, Regensburg, Germany were added in a concentration range from 2.47 nM to 200 nM. After the generation of a four-parameter-dose–response calibration curve, synthetic peptides were added in a range from 0.21 nM to 50 nM. As a positive control, the supernatant of the B. subtilis wild type strain was supplemented with synthetic α- and P–pheromone and added to the ELISA plate. Next, the plate was covered with a lid and stored at 4 °C overnight or at room temperature for 2 h. After 2 h, wells were washed three times with 200 µL of PBST buffer (8 g NaCl, 0.2 g KCl, 1.44 g Na_2_HPO_4_, 0.24 g KH_2_PO_4_, 0.1 ml Tween20, pH 7.2) and residual binding sites on ELISA plates were blocked with 100 µL of 1 % (w/v) BSA (bovine serum albumin) dissolved in PBST buffer for 2 h at room temperature. The ELISA plate was rewashed three times with PBST buffer, and 100 µL of 0.4 µg mL^−1^ α- or P-pheromone antibody from rabbit (Peninsula Laboratories International, Inc. (currently BMA Biomedicals, Switzerland) and David’s Biotechnology) dissolved in 0.5 % (w/v) BSA in PBST was added. After 1 h, wells were rewashed and 100 µL of Anti-Rabbit IgG (H + L) HRP Conjugate (working solution 1:2500 diluted in 0.5 % BSA in PBST buffer) (Promega) was added. After 1 h, the last washing step was performed and the peroxidase reaction was started by adding 100 µL of Pierce™ TMB substrate kit solution in each well. The reaction was stopped by adding 100 µL of 2 M sulfuric acid. Subsequently, absorbance was measured at 450 nM in a Synergy Neo3 Hybrid Multimode Microplate Reader from BioTek (Winooski, VT, USA).
Results
3
Establishment of the evaluation vector and signal peptide toolbox
3.1
To develop a highly versatile and easy-to-use vector for heterologous protein expression in B. subtilis, we utilised the well-established pBS1C.14 Desired characteristics of this vector are a moderate size (<7 kb), a copy number of 20–30 in E. coli, and the possibility of integration into the amyE locus of B. subtilis, for which the starch assay is a low-cost screening method to determine B. subtilis mutants. First, we inserted the well-characterised xylose inducible promoter, PxylA, in the adopted BioBrick^TM^ RFC10 standard into pBS1C to enable the exchangeability of the promoter if desired (Fig. 1A). In the next step, we placed gene sequences of rfp and lacZα into the vector to establish the final evaluation vector pBS1C-PxylA-rfp-lacZα, in which the SP and the sequence of POI can be cloned via the restriction sites AgeI and NgoMIV (Fig. 1A and Fig. S3).Fig. 1. Evaluation vector establishment and application. A Schematic overview of the multiple cloning site development into the evaluation vector. (I) Xylose inducible promoter was fused into the BsaI-free pBS1C background vector. (II) Multiple cloning site was enriched with synthesized (fused to P_lacI_) rfp_syn_ and lacZα to establish the evaluation vector. Black box indicate which restriction enzyme was used for cloning. B Schematic overview of evaluation vector application. POI-protein of interest, SP-signal peptide, E-EcoRI, N-NotI, X-XbaI, B-BsaI (XbaI overhang), A-AgeI, Ngo-NgoMIV, S-SpeI and P-PstIA.
Based on the rfp and lacZα genes in the MCS, the evaluation vector provides a quick identification of integrated SP and POI inserts via red-blue colony screening in E. coli (Fig. 1B). When desired, the inducible PxylA promoter that drives the POI expression in the evaluation vector can be exchanged easily due to the presence of the BsaI restriction enzyme cleavage site (Fig. 1B). For this study, we first introduced α- and P-pheromone gene sequences from S. cerevisiae and S. pombe as POI into the evaluation vector. Codon adapted sequences of both pheromones were utilised to ensure efficient heterologous expression in B. subtilis. Thus, we designed a set of coding sequences adapted for heterologous expression in B. subtilis (Fig. S1 and S2). After coding sequences were inserted into the evaluation vector, the SP toolbox was used to randomly introduce SP sequences in front of α- and P-pheromone encoding sequences, respectively. In general, the SP toolbox is not a predictive tool designed to directly estimate SP-POI relationships. However, to address the lack of computational SP prediction tools, we developed an experimental approach that is both fast and easily implementable, enabling the efficient identification of the optimal SP-POI pairing. 74 SP sequences, divided into four SP-mix libraries, were generated (Table 2). Each library contains 0.5 ng µl^−1^ DNA concentration of each SP pre-cloned in pSB1C plasmids (Table 2).Table 2. Sec secretion signal peptides used in this study are divided alphabetically in signal peptide mix subsets.Signal peptide mix: Subset 1AmyEAspBBglSBprCccACitHCsnDacBDacFDltDEprFliLFliZGlpQLipALytBLytCLytDSignal peptide mix: Subset 2LytRMdrMprNprEPbpXPelPelBPenPPhoAPhoBPhrAPhrCPhrFPhrGPhrKRpmGSignal peptide mix: Subset 3SacBSacCSleBSpollDSpollPSpollQSpollRTyrAVprWapAYbbCYbbCYbbRYbdGYbdNYbfOYbxIYdbKYdhTYdjMSignal peptide mix: Subset 4YdjNYfhKYfjSYfkDYfkNYhcRYhdCYhfMYhjAYjcNYjdBYjfAYjiAYkoJYkvVYkwDYlaEYlbLYlqBYlxF
SP names derive from the protein names for which the SPs designate secretion in their native context. Additionally, each SP sequence carries an RBS, a spacer sequence optimised for expression in B. subtilis, followed by the start codon. Protein sequences of all 74 signal peptides can be found in Table S5. Furthermore, each SP sequence holds identical flanking DNA sequences up- and downstream that allow multi-template PCR to enrich the SP mix subsets while using the toolbox.
Evaluation of the secretion peptide toolbox
3.2
Enriched peptide mix subsets were digested with XbaI and AgeI, cloned into the evaluation vector already carrying the yeast pheromones, and transformed with E. coli. Afterwards, white colonies, potentially carrying correct SP-POI constructs, were picked for plasmid isolation. Plasmids were transformed to B. subtilis and 50 colonies for each SP subset were transferred to starch agar plates to check the successful plasmid integration into the B. subtilis genome with Lugoĺs solution. B. subtilis colonies, including the artificial SP-POI construct within the genome, were then transferred to 5 µg mL^−1^ chloramphenicol agar plates for storage and subsequent expression/secretion experiments. To evaluate expression of SP-POI constructs, generated B. subtilis strains were grown to the exponential phase and induced with the addition of 0.5 % (w/v) xylose. Supernatants were collected after 3 h and used for indirect ELISA assays to probe successful secretion of α- and P-pheromones (Fig. 2). Successful expression and secretion of P-pheromone was conducted by using SP-mix subsets 1, 2, 3, and 4 in combination with the harmonised and most frequent coding sequence of P-pheromone (Fig. 2A). Among SP mixtures, the majority of secreting P-pheromone clones belong to SP-mix subset 1. On the other hand, no secretion of P-pheromone for SP-mix subset 4 was observed (Fig. 2A). Further, only 14 clones out of 600 tested were able to secrete any P-pheromone, highlighting that the optimal SP-POI combination is critical for secretion of heterologously expressed proteins in B. subtilis (Fig. 2 and Table 3). However, the pheromone coding sequence and therein embedded codon usage of B. subtilis play an important role as well in the expression and secretion of artificial proteins. The darker colour in the heat map indicates a higher concentration of P-pheromone encoded by the most frequent adapted coding sequence compared to the harmonised coding sequence. For the robust coding sequence, no secreted P-pheromone could be detected (Fig. 2A). To identify the SPs yielding in P-pheromone secretion, the 14 clones were identified by sequencing. In the case of α-pheromone secretion, we detected the secreted peptide for every optimised coding sequence across each SP-mix subset (Fig. 2B). However, all determined yields for α-pheromone production resulted in low concentrations at the limit of detection (Fig. 2B). Since none of the tested colonies showed a high concentration of secreted α-pheromone, random clones were chosen for sequencing and thereby the identification of SP responsible for peptide secretion (Table 3). In total, seven SPs that contribute to the secretion of P-pheromone and three SPs responsible for α-pheromone secretion in B. subtilis were identified (Table 3). In summary, the toolbox developed in this study provides a powerful tool to screen a versatile variety of SPs in a single step and subsequently identify promising pairs of SP(s) and POI for successful secretion.Fig. 2P- and α-pheromone secretion screening by ELISA assay. ELISA assay was performed on 1200 colonies to screen the SP-Mix subsets (SPM1-4), each fused to the three optimised coding sequences (CDS), for (A) P-pheromone and (B) α-pheromone secretion in B. subtilis. Cultures were grown in LB media and expression/secretion was induced with 0.5 % (v/v) xylose. Heat map represents the average value of biological duplicates. Colour gradient is in the range from a minimum pheromone concentration of 2.47 nM to a maximum concentration of 200 nM.Table 3. Identified signal peptides responsible for α- and P-pheromone peptide secretion.SP for P-pheromone secretionSP for α-pheromone secretionBglSYfkDBprYfkNDacFYhcRFliZSacCYbxIYdjNThe signal peptide protein sequence is shown in Table S5.
Enhancement of pheromone production by fine-tuning expression parameters
3.3
The next step in secretion improvement consisted of the heterologous pheromone expression under the control of an antibiotic-regulated promoter. Here, we chose the well-studied antibiotic inducible promoter PliaI of B. subtilis to increase the secretion of α- and P-pheromone. The PliaI promoter has been selected due to the high induction activity (∼100-fold) after adding the antibiotic bacitracin.23, 24 Reasoned by the low secretion of α–pheromone, we decided to optimise and test the expression/secretion level of P-pheromone only by using PliaI. The optimised (harmonised and most frequent) coding sequence of P-pheromone fused to the seven selected SPs (Table 3) were fused to PliaI. Then, generated strains were grown and induced with the antibiotic bacitracin with concentrations ranging from 15 to 100 µg mL^−1^ to start heterologous expression and secretion of P-pheromone in B. subtilis. We found that expression of the construct PliaI-ybxI-P_H_ resulted in the highest concentration of secreted P-pheromone (Fig. S4). Additionally, the next best-performing expression cassette is PliaI-ybxI-P_MF,_ indicating optimal SP for P-pheromone secretion is YbxI, which will be used for further experiments in this study (Fig. S4). After the identification of YbxI-SP as the best-performing SP for P-pheromone secretion, we started to characterise the concentration of bacitracin that works best for optimal promoter induction and peptide expression time interval (Fig. 3). In general, increasing bacitracin concentrations resulted in increasing P-pheromone secretion in a time range between 0.5 and 3 h. The highest amount of secreted P-pheromone was detected after the addition of 100 µg mL^−1^ of bacitracin and an expression time of 1 h (Fig. 3). A longer period (> 3h) of expression cultivation resulted in a decrease of P-pheromone secretion, which can be observed for all tested bacitracin concentrations. The next question we asked ourselves was to what extent the optical density of the cells to be induced affects P-pheromone secretion (Fig. 4). To do so, different amounts of B. subtilis overnight culture (33 %, 17 %, 6 %, 3 %, 2 % and 1 %) were diluted in fresh media and cultivated for 3 h before 100 µg mL^−1^ bacitracin was added. One hour after induction, supernatants were collected and analysed by ELISA. We found that an amount of 3 % to 17 % of overnight cells, diluted into fresh media and induced with bacitracin, resulted in the highest levels of P-pheromone secretion (Fig. 4). The most substantial presence of P-pheromone in culture supernatant was detected when 6 % of cells were used to inoculate the fresh media.Fig. 3. Characterisation of bacitracin-induced P-pheromone production in B. subtilis. Strain B. subtilis W168 P_liaI_-ybxI-P_H_ was induced with bacitracin (concentration range 15–100 µg mL^−1^) and samples were collected after 0.5, 1.0, 2.0, 3.0 h and analysed by indirect ELISA assay. Heat map represents the average value of triplicates. Colour gradient is in the range from minimum P-pheromone concentration 2.47 nM to maximum concentration of 200 nM P-pheromone.Fig. 4. Influence of the B. subtilis culture start optical density on the P-pheromone production. Overnight culture of B. subtilis W168 PliaI-ybxI-P_H_ was mixed with fresh media in different ratios (shown as the percentage of overnight culture in fresh media) and expression started after 3 h of cultivation by induction with bacitracin (100 µg mL-1). Samples were harvested 1 h after the induction and analyzed with indirect ELISA. Experiments were performed as technical triplicates. Statistical difference was determined using one-way ANOVA followed by Bonferroni’s multiple comparisons test to do pairwise comparison between start optical densities. Error bars present the standard deviation from the mean value. ns = not significant, ** = p < 0.01, and *** = p < 0.001.
In line with our observation of decreasing P-pheromone concentration in supernatants after an expression time of more than 1 h and 75 or 100 µg ml^−1^ of bacitracin (see Fig. 3), we thought about the possibility of peptide instability or degradation by extracellular proteases of B. subtilis W168. To investigate if the extracellular proteases are a reason for lower P-pheromone amounts in supernatants, we transferred the most promising plasmids containing P-pheromone secretion constructs to the strain B. subtilis WB800N that is deficient for eight extracellular proteases.25 Heterologous expression and secretion of P-pheromone in B. subtilis WB800N was compared to the secretion conducted in B. subtilis W168 (Fig. 5). We found a significantly higher amount of P-pheromone in supernatants of B. subtilis WB800N strains compared to B. subtilis W168 strains. These results indicated that, even though we added a protease inhibitor cocktail directly after harvesting the supernatants of B. subtilis W168, protease activity was high enough to reduce the pheromone concentration over time of cultivation. However, wild-type and WB800N strains including PliaI-ybxI-P_H_ show similar secretion levels (Fig. 5). This result is due to the saturation limit (outside of the linear working range) of the plate reader and the necessary optimisation of the indirect ELISA assay. Nevertheless, these results again indicate that expression and secretion of P-pheromone under PliaI and SP YbxI seem to be the best. Additionally, B. subtilis WB800N was the strain of choice for further pheromone secretion experiments in this work.Fig. 5. Difference in secretion performance between B. subtilis strains W168 and WB800N. Production of P-pheromone with best performing signal peptides with either one of two coding sequences (H-harmonised, MF-Most frequent) was analysed in B. subtilis strains WB800N and W168. Indirect ELISA was used to detect the secreted P-pheromone. Experiments were performed as technical triplicates. Statistical difference was determined using an unpaired t-test with Welch's correction test to do comparison between two B. subtilis species. Error bars present standard deviation from the mean value, * = p < 0.05, and **** = p < 0.0001.
As next step, the optimisation and characterisation results of the strain B. subtilis WB800N, including the bacitracin-inducible promoter PliaI fused to the SP P-pheromone (harmonised or most frequent sequence), were transferred to improve the expression and secretion of α-pheromone. Thus, the SP-α–pheromone secreting cassette was fused with PliaI and transformed with B. subtilis WB800N. Again, secretion studies were performed by indirect ELISA and the results of the α-pheromone secretion are shown in Fig. 6. In general, α–pheromone was successfully secreted by the used strains (see Fig. 6). However, the amount of secreted α–pheromone is low compared to the negative control for which supernatant of B. subtilis WB800N non-producer strain was used. The highest amount of α–pheromone was achieved with YfkD SP. Furthermore, in contrast to the P-pheromone secretion strains, there is no dominating α-pheromone coding sequence and similar amounts of α–pheromone were observed for both the most frequent (MF) and harmonised (H) sequence. It was decided to use the harmonised coding sequence of α-pheromone for further experiments.Fig. 6α-pheromone secretion in B. subtilis WB800N. α-pheromone detected in supernatant of B. subtilis WB800N strains with indirect ELISA. Negative control (Neg. cont.) is supernatant of B. subtilis WB800N non-producer strain. Strain is missing the expression/secretion cassette. Positive control (Pos. cont.) is the standard concentration (100 nM) of synthetic α −pheromone added into the supernatant of B. subtilis WB800N non-producer strain. Experiments were performed as technical triplicates. Statistical difference was determined using one-way ANOVA followed by Bonferroni’s multiple comparisons test to do pairwise comparison between negative control and α-pheromone producing strains. Error bars present standard deviation from the mean value, ns = not significant, * = p < 0.05, *** = p < 0.001, and **** = p < 0.0001.
Finally, to determine the precise concentration of secreted α- and P-pheromone in supernatants, a four-parameter-dose–response curve was established, which maps a concentration range from 0.21 nM up to 50 nM of α- and P-pheromone (Fig. S5). Concentration determination of α- and P-pheromone in supernatants of B. subtilis WD800N PliaI-ybxI-P_H_ for P-pheromone and PliaI-yfkD-α_H_ for α-pheromone, respectively, are shown in Fig. 7. One hour after induction with bacitracin, 43 nM P-pheromone could be determined in supernatants of strain PliaI-ybxI-P_H,_ while strain PliaI-yfkD-α_H_ secreted 8 nM of α-pheromone. Even though secreted pheromone peptide concentration is low compared to industry standards, production was repeatable and robust in B. subtilis. This lets us conclude that the developed evaluation vector and the SP toolbox have proven to be useful for heterologous protein secretion in B. subtilis.Fig. 7. Concentration of secreted α- and P-pheromone at various time points. Detected concentration of heterologously secreted α- and P- pheromone in B. subtilis WB800N PliaI-ybxI-P_H_ for P-pheromone and PliaI-yfkD-α_H_ for α-pheromone. Indirect ELISA was used to determine the α- and P-pheromone concentrations. Experiments were performed as technical triplicates. Error bars present the standard deviation from the mean value. Statistical significance was determined with an unpaired t-test with Welch’s correction (** = p < 0.01, *** = p < 0.001).
Discussion
4
B. subtilis has already been used as a workhorse of White Biotechnology for a long time. Wide application of B. subtilis was secured due to its GRAS (generally recognised as safe) status and ability to secrete high amounts of enzymes in relatively cheap culture media.4 However, Fu et al., 2018 highlight that a universal SP for protein secretion does not exist, leaving library construction and high-throughput screening approaches the only way to identify, boost expression and render secretion for POI production via SPs.9.
In this work, we wanted to develop a faster way to find the optimal SP for protein secretion in B. subtilis. Therefore, we established the SP toolbox, including an evaluation vector and 74 SPs. The evaluation vector based on the backbone vector pBS1C14 proved to be a feasible and easy-to-use vector in this study. The SP toolbox offers a shotgun approach for high-throughput screening to identify the optimal SP for the POI secretion. Furthermore, the advantage of the developed toolbox here is the design in which SPs were fused with optimised ribosome binding site, spacer and start codon from B. subtilis within the storage vector. In future, a toolbox can be easily expanded and more SPs can be added to test their secretion capacities. In this work, eight out of 74 SP are successfully secreting the P-pheromone. The identified SPs responsible for P-pheromone secretion do not have any observable pattern in common. Random correlation between SPs and secreted protein was already observed previously.9, 11 Unpredictability and unavailability of secretion standardisation (in silico prediction tools) are the reasons for the development of the toolbox in this study.
It is worth noticing the two amino acid linkers consisting of threonine and glycine between SPs and the protein of interest, which is a product of restriction cloning with enzymes AgeI and NgoMIV. Two-amino acid linker was removed in Fu et al., 2018, due to the hypothesis that additional residues influence the cleavage efficiency of SPe for protein secretion.9 However, we didn’t test the influence of two amino acid residues on the secretion efficiency because of the wide use of restriction cloning in the development of similar SP libraries.11.
By using the newly developed toolbox for secretion of heterologous expressed proteins and peptides, we were able to identify eight SPs for effective P-pheromone secretion, whereas three SPs out of 74 were responsible for α-pheromone secretion. Interestingly, all 11 SPs (see sequence in table S6) vary in their length and share the characteristic α-helix structure of the signal peptide. The structure of the signal peptides can be seen by using the database SubtiWiki.26 Additionally, after identifying SPs necessary for pheromone secretion, secretion can be maximised through directed evolution and high-throughput screening, as in Shi et al., 2025. Furthermore, we utilised the well-characterised antibiotic-inducible promoter PliaI to optimise peptide secretion in the B. subtilis.23, 24, 27 High inducibility of PliaI with the antibiotic bacitracin boosts peptide expression within the exponential growth phase and could prevent or reduce the degradation process of secreted peptides by growth-dependent accumulation of extracellular proteases (e. g. Bpr, Npr, Epr, or Vpr) of B. subtilis. To circumvent the mentioned degradation problem, we also exchanged the production host strain from wild-type to the protease-deficient strain B. subtilis WB800N that shows an 8-fold lower protease activity.19 As expected, we observed a higher peptide concentration when strain B. subtilis WB800N was used for overexpression and secretion of P-pheromone. Since B. subtilis WB800N is deficient for only a fractional number of available proteases in B. subtilis8 we assumed that protease activity is still present and will lower the concentration of expressed and secreted P- and α-pheromones. Here, specialised degradation proteases like Clp, ClpQ or Lon, or protein quality control factors like HtrA and HtrB can contribute to degrade heterologously expressed peptides.8, 28, 29 Improvement in protein expression and preventing degradation of expressed proteins and peptides can be achieved by deleting additional protease-encoding genes from strain B. subtilis WB800N. An alternative strategy is to use ‘midiBacillus’ or ‘miniBacillus’ strains.30, 31 These strains lacking around 36 % of the genome and almost all proteases are characterised by giving the advantage to the production of “difficult-to-produce proteins” but not yet small peptides. However, in case of the expression and secretion of α-pheromone, additional removal of more proteases from the host strain will be most beneficial due to the fact that the 13 amino acid long α-peptide is active as an unfolded protein.32 Unfolded secreted proteins are targets for B. subtilis extracytoplasmic folding catalysts and quality control proteases, which efficiently remove misfolded or incompletely synthesised proteins.8 Unfolded state of α-pheromone and weak secondary structure of P-pheromone is also a reason why we choose Sec secretion system and not TAT-pathway. The TAT pathway is used for the secretion of fully folded proteins with tightly bound cofactors, while proteins secreted by the Sec pathway are unfolded.33
In summary, efficient protein expression and secretion in B. subtilis is easily possible by using our developed SP-toolbox. Furthermore, the toolbox represents a strategically important application to circumvent a main problem in protein secretion: lack of knowledge regarding the optimal and functional SP essential for effective secretion. In the future, SP toolbox can be improved by expanding it and adding more SPs (up to 173 available in B. subtilis, Fu et al., 2018) to the library or developing the protease-free strain of B. subtilis.9
CRediT authorship contribution statement
Tomislav Vološen: Writing – review & editing, Writing – original draft, Visualization, Validation, Methodology, Investigation, Formal analysis, Data curation. Henri Max Deda: Writing – review & editing, Validation, Software, Investigation, Formal analysis, Data curation. Anica Walther: Writing – review & editing, Validation, Resources, Methodology, Investigation. Philipp F. Popp: Writing – review & editing, Supervision, Project administration, Funding acquisition, Formal analysis. Thorsten Mascher: Writing – review & editing, Supervision, Project administration, Conceptualization. Diana Wolf: Writing – review & editing, Writing – original draft, Validation, Supervision, Project administration, Funding acquisition, Conceptualization.
Declaration of competing interest
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Diana Wolf reports financial support was provided by European Social Fund. Tomislav Volosen reports financial support was provided by European Social Fund. Henri Max Deda reports financial support was provided by iGEM Foundation. Anica Walther reports administrative support and writing assistance were provided by European Social Fund. Philipp F. Popp reports administrative support and writing assistance were provided by iGEM Foundation. Thorsten Mascher reports administrative support was provided by iGEM Foundation. If there are other authors, they declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Global technical enzymes market research report [Internet]. [cited 2024 Dec 20]. Available from: https://www.sphericalinsights.com/reports/enzymes-market.
- 2Hemmerich J.Rohe P.Kleine B.Use of a Sec signal peptide library from Bacillus subtilis for the optimization of cutinase secretion in Corynebacterium glutamicum Microb Cell Fact 15120162082792720810.1186/s 12934-016-0604-6PMC 5142396 · doi ↗ · pubmed ↗
- 3Corrêa G.G.da Lins M.R.C.R.Silva B.F.de Paiva G.B.Zocca V.F.B.Ribeiro N.V.A modular autoinduction device for control of gene expression in Bacillus subtilis Metab Eng 6120203263343237109010.1016/j.ymben.2020.03.012 · doi ↗ · pubmed ↗
- 4Schallmey M.Singh A.Ward O.P.Developments in the use of bacillus species for industrial production Can J Microbiol 50120041171505231710.1139/w 03-076 · doi ↗ · pubmed ↗
- 5Ejaz S.Khan H.Sarwar N.Aqeel S.M.Al-Adeeb A.Liu S.A review on recent advancement in expression strategies used in Bacillus subtilis Protein Pept Lett 29920227337433592781110.2174/0929866529666220803163335 · doi ↗ · pubmed ↗
- 6Kang Z.Yang S.Du G.Chen J.Molecular engineering of secretory machinery components for high-level secretion of proteins in bacillus species J Ind Microbiol Biotechnol 41112014159916072521224610.1007/s 10295-014-1506-4 · doi ↗ · pubmed ↗
- 7Tsirigotaki A.De Geyter J.Šoštaric N.Economou A.Karamanou S.Protein export through the bacterial Sec pathway Nat Rev Microbiol 151201721362789092010.1038/nrmicro.2016.161 · doi ↗ · pubmed ↗
- 8Tjalsma H.Antelmann H.Jongbloed J.D.H.Proteomics of protein secretion by Bacillus subtilis: Separating the “secrets” of the secretome Microbiol Mol Biol Rev 68220042072331518718210.1128/MMBR.68.2.207-233.2004 PMC 419921 · doi ↗ · pubmed ↗