Dynamics of postnatal bone development and epiphyseal synostosis in the caprine autopod
Christopher J. Panebianco, Maha Essaidi, Elijah Barnes, Ashley Williams, Karin Vancíková, Margot C. Labberté, Pieter Brama, Niamh C. Nowlan, Joel D. Boerckel

TL;DR
This study explores how bones in goats develop after birth, showing how movement affects bone structure and fusion in the lower forelimbs.
Contribution
The study identifies a novel postnatal synostosis event in the caprine metacarpal epiphysis linked to ambulatory loading.
Findings
P1 cortical bone densified rapidly after birth, with continual increases in cortical thickness through adulthood.
Metacarpal epiphyses undergo postnatal synostosis through fibrocartilaginous endochondral ossification prior to growth plate closure.
Abstract
Bones develop to structurally balance strength and mobility. Bone developmental dynamics are influenced by whether an animal is ambulatory at birth. Precocial species, which are ambulatory at birth, develop advanced skeletal maturity in utero and experience postnatal development under mechanical loading. Here, we characterized postnatal bone development in the lower forelimbs of precocial goats using microcomputed tomography and histology. Our analysis focused on the two phalanges 1 (P1) bones and the partially fused metacarpal bone of the goat autopod from birth through adulthood. P1 cortical bone densified rapidly after birth, but cortical thickness increased continually through adulthood. Upon normalization by body mass, the P1 normalized polar moment of inertia was constant over time, suggestive of changes correlating with ambulatory loading. P1 trabecular bone increased in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
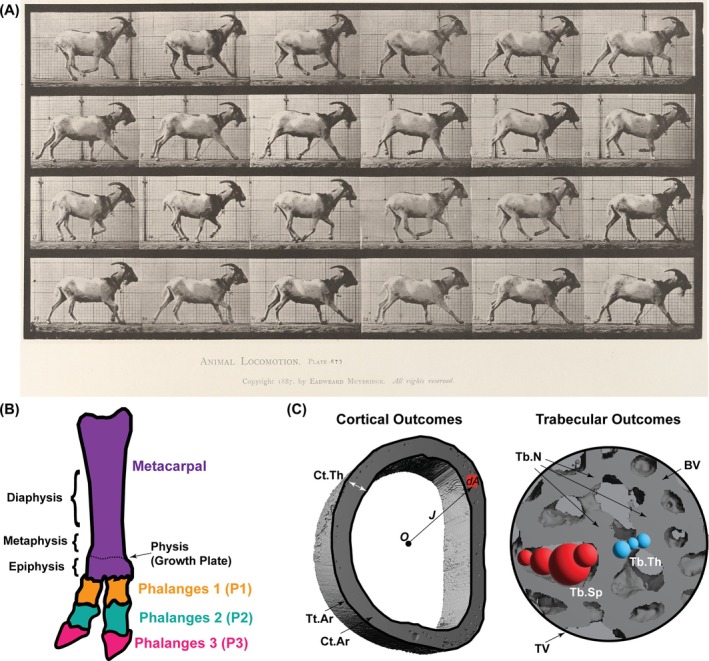 FIGURE 1
FIGURE 1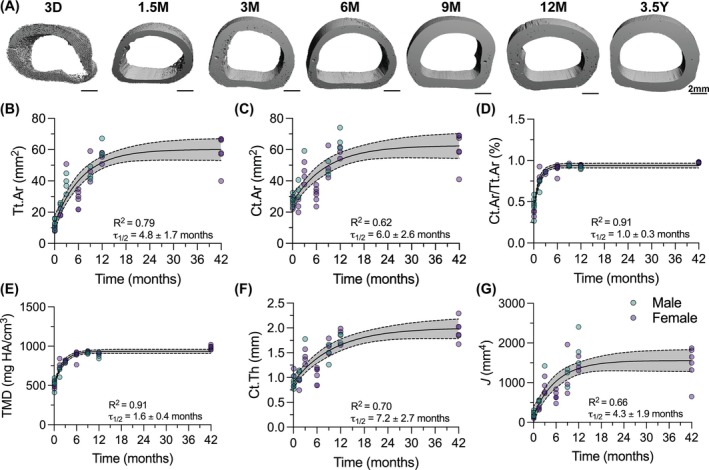 FIGURE 2
FIGURE 2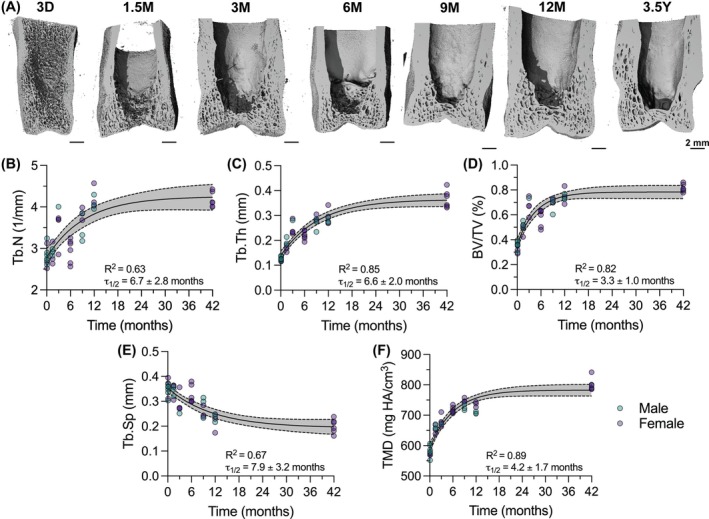 FIGURE 3
FIGURE 3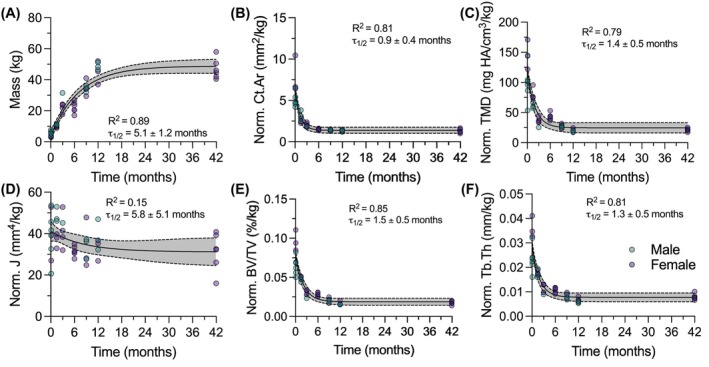 FIGURE 4
FIGURE 4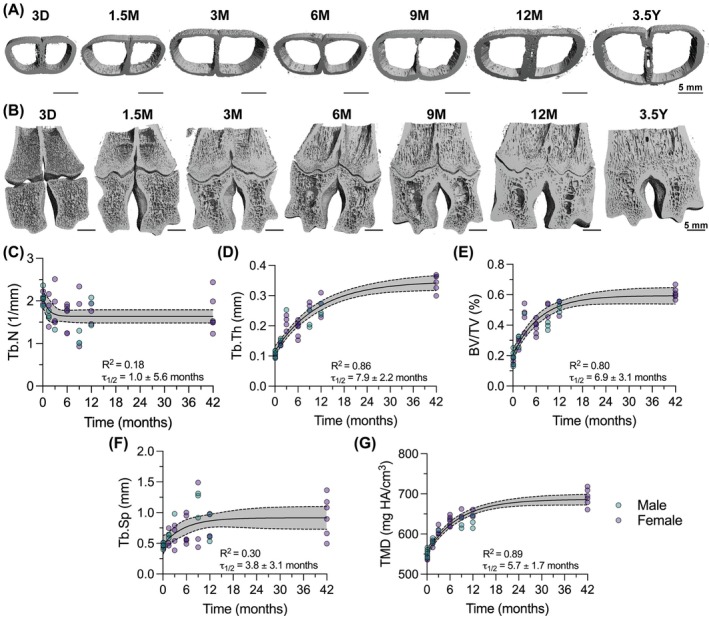 FIGURE 5
FIGURE 5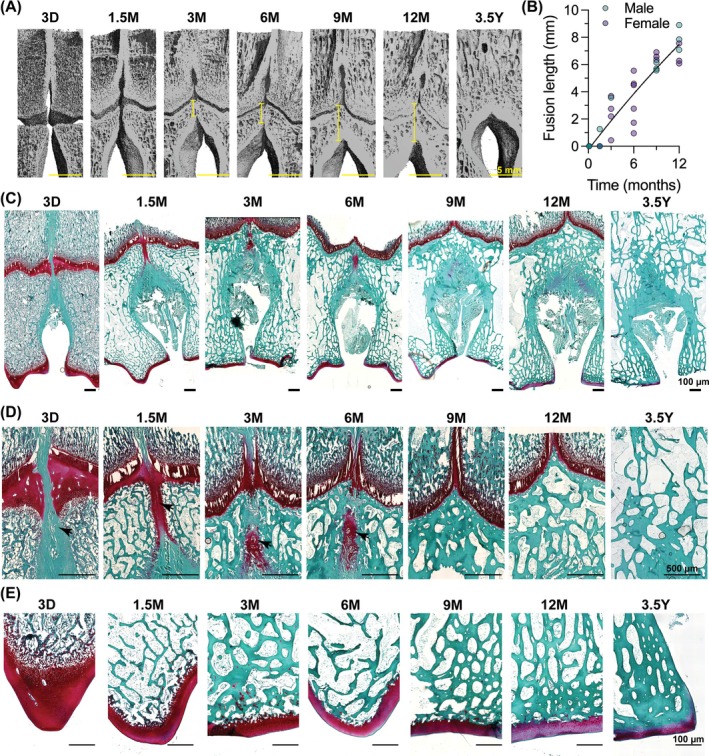 FIGURE 6
FIGURE 6- —Science Foundation Ireland10.13039/501100001602
- —Research Ireland
- —National Institute of General Medical Sciences10.13039/100000057
- —National Institute of Child Health and Human Development10.13039/100009633
- —HORIZON EUROPE European Innovation Council10.13039/100018703
- —National Science Foundation10.13039/100000001
- —National Institute of Arthritis and Musculoskeletal and Skin Diseases10.13039/100000069
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBone Metabolism and Diseases · Connective tissue disorders research · Bone and Dental Protein Studies
INTRODUCTION
1
The skeleton continuously remodels throughout mammalian growth and development, creating a structure optimized for high strength and low weight. While we have long known that mature bone adapts and remodels based on loading,1, 2 it has been difficult to decouple early bone development programs from adaptation to ambulatory loading. As precocial species, goats are ambulatory at birth, providing continuous loading during skeletal growth. Here, we mapped the developmental dynamics of the autopod of the caprine lower forelimb from birth through adulthood, focusing on the phalanges 1 (P1) and metacarpal bones in the autopod.
An animal's ability to ambulate at birth influences the dynamics of postnatal skeletal development. Humans, mice, and rats are examples of altricial species, meaning they are non‐ambulatory at birth. Altricial species initiate primary ossification in utero, under conditions of dynamic fetal movement,3, 4, 5, 6 then the majority of secondary ossification centers initiate after birth.5, 7, 8 Unlike altricial species, precocial species such as goats, horses, and sheep, ambulate from birth through adulthood (Figure 1A) and undergo primary and secondary ossification in utero as an anticipatory mechanism to prepare for future loading.9, 10, 11 It has been hypothesized that this early secondary ossification evolved to support the physis to withstand high mechanical loading shortly after birth.12 Though previous research has described gestational bone development of precocial species, few studies have investigated their postnatal development. Understanding postnatal developmental dynamics in precocial species may inform how mechanical cues influence skeletal morphogenesis.
Goats are precocial animals that ambulate from birth through adulthood. (A) Reproduction of 1887 Animal Locomotion Plate 676, captured by Eadweard Muybridge at the University of Pennsylvania. Plates provided by the University of Pennsylvania University Archives and Records Center. (B) Schematic representation of the caprine lower forelimb showing the different bones (i.e., metacarpal, phalanges 1 (P1), P2, and P3) and key anatomical regions of the metacarpal bone (i.e., physis/growth plate, epiphysis, metaphysis, and diaphysis). (C) Schematic representation of key cortical and trabecular outcomes. Cortical outcomes: Cortical tissue cross‐sectional area (Tt.Ar), cortical bone area (Ct.Ar), cortical area fraction (Ct.Ar/Tt.Ar), cortical thickness (Ct.Th), tissue mineral density (TMD), and polar moment of inertia (J). Trabecular outcomes: Bone volume fraction (BV/TV), trabecular number (Tb.N), trabecular thickness (Tb.Th), and trabecular separation (Tb.Sp). In the representation of J, O represents the origin of a polar coordinate system, and dA represents a differential area.
Diaphyseal synostosis has evolved in a variety of species, likely to provide structural resistance to high bending forces. For example, the jerboa, a bipedal rodent, evolved fused metatarsal bones that reduce peak bone stresses and provide a factor of safety to the hindlimb during bipedal ambulation.13 This evolutionary advantage contributes to the jerboa's ability to jump 10 times its hip height and withstand peak ground‐reaction forces that are five times its body weight.14, 15 As altricial species with limited hindlimb loading immediately after birth, jerboa metatarsal synostosis occurs postnatally.16, 17 Diaphyseal synostosis is also observed in some precocial species. For example, each goat forelimb has two independent metacarpal rudiments embryonically, but these rudiments undergo diaphyseal synostosis in utero to form one metacarpal bone at birth (Figure 1B). As distal elements (e.g., autopod) mature later in development than the more proximal zeugopod and stylopod, developmental reasoning suggests that the ungulate diaphyseal synostosis initiates in utero to provide increased strength and enable locomotion at birth.18 It remains unknown whether these fusion events continue through postnatal development. These findings may be useful for understanding how biomechanical pressures guide bone development in precocial species.
Here, we characterize the dynamics of postnatal bone development and epiphyseal synostosis in the caprine autopod from birth through adulthood. P1 cortical bone rapidly densified, then became continually thicker until sexual maturity (12 months), with structural distribution determined by ambulatory loading. Simultaneously, P1 trabecular bone increased in trabecular number and thickness until sexual maturity, while metacarpal trabecular number was constant and the network grew through trabecular thickening. Metacarpal synostosis began in utero; however, the epiphyses did not begin to fuse until after 1.5 months postnatally. Histological analyses indicated that this epiphyseal synostosis occurred through fibrocartilaginous endochondral ossification, prior to growth plate closure. Together, these data provide new insights into precocial bone development under ambulatory loading.
RESULTS
2
Phalanges 1 (P1) postnatal development dynamics
2.1
Phalanges 1 (P1) bones showed significant increases in all outcomes for cortical bone morphometry over postnatal development (Figure 2). To better understand the dynamics of caprine P1 development over time, each outcome measurement was fit with a one‐phase exponential decay model. Curves fit well for all outcomes with coefficients of determination (R ^2^) values between 0.62 and 0.91 (Figure 2B–G). By extracting the half‐life values (τ 1/2) from these curves, we could determine the characteristic time scale for each outcome. Outcomes with lower τ 1/2 values achieved 50% of their maximum value earlier in developmental time than outcomes with higher τ 1/2 values. Thus, outcomes with lower τ 1/2 values achieved their maximum value faster and the outcome plateaued earlier in developmental time than outcomes with higher τ 1/2 values. Cortical area fraction (Ct.Ar/Tt.Ar, τ 1/2 = 1.0 ± 0.3 mo, Figure 2D), a measure of cortical porosity, and tissue mineral density (TMD, τ 1/2 = 1.6 ± 0.4 mo, Figure 2E) plateaued most rapidly. Total cross‐sectional area (Tt.Ar, τ 1/2 = 4.8 ± 1.7 mo, Figure 2B), and cortical bone area (Ct.Ar, τ 1/2 = 6.0 ± 2.6 mo, Figure 2C), and cortical thickness (Ct.Th, τ 1/2 = 7.2 ± 2.7 mo, Figure 2F) plateaued less rapidly and continued to increase through 12 months. Polar moment of inertia (J, τ 1/2 = 4.3 ± 1.9 mo, Figure 2G), which measures the distribution of the cortical bone about the central axis, also plateaued less rapidly and continued to increase through 12 months. These results indicate that P1 bones undergo early cortical condensation followed by progressive cortical thickening into adulthood.
Phalanges 1 (P1) cortices exhibit rapid condensation and continuous expansion until adulthood. (A) Transverse views of 3D microcomputed tomography (microCT) reconstructions of P1 cortical mid‐shaft. Scale bars = 2 mm. (B) Cortical tissue cross‐sectional area (Tt.Ar), (C) Cortical bone area (Ct.Ar), (D) Cortical area fraction (Ct.Ar/Tt.Ar), (E) Tissue mineral density (TMD), (F) Cortical thickness (Ct.Th), and (G) Polar moment of inertia (J) throughout developmental time, fit with a one‐phase decay model. Gray bands indicate 95% confidence interval for each curve for the full dataset. Datapoint color indicates animal sex: Male = teal, female = purple. Goodness of fit (R 2) and half‐life (τ 1/2) are reported for each graph. D, days of age; M, months of age; Y, years of age.
P1 distal trabecular microarchitecture also showed significant changes in all outcomes over postnatal development (Figure 3). Trabecular number (Tb.N, τ 1/2 = 6.7 ± 2.8 mo, Figure 3B) and trabecular thickness (Tb.Th, τ 1/2 = 6.6 ± 2.0 mo, Figure 3C) increased continuously through 12 months. As a result of this increased formation of new trabecular networks and growth of existing trabecular networks, the bone volume fraction (BV/TV, τ 1/2 = 3.3 ± 1.0 mo, Figure 3D) increased and the trabecular separation (Tb.Sp, τ 1/2 = 7.9 ± 3.2 mo, Figure 3E) decreased continuously through 12 months. Simultaneously, the existing trabeculae were increasing in mineralization, as shown by the continuous changes in TMD (τ 1/2 = 4.2 ± 1.7 mo, Figure 3F). Curves fit well for all outcomes with R ^2^ values between 0.63 and 0.85 (Figure 3B–F). These data suggest that P1 postnatal cancellous bone formation is characterized by both the formation of new trabecular networks and the growth of existing trabecular networks.
Distal P1 bones grow by formation of new trabeculae and expansion of existing trabeculae. (A) Transverse views of 3D microCT reconstructions of P1 bones with a virtual slice through the mid‐coronal plane using microCT. Scale bars = 2 mm. (B) Trabecular number (Tb.N), (C) Trabecular thickness (Tb.Th), (D) Bone volume fraction (BV/TV), (E) Trabecular spacing (Tb.Sp), and (F) Tissue mineral density (TMD) throughout developmental time, fit with a one‐phase decay model. Gray bands indicate 95% confidence interval for each curve for the full dataset. Datapoint color indicates animal sex: Male = teal, female = purple. Goodness of fit (R 2) and half‐life (τ 1/2) are reported for each graph. D, days of age; M, months of age; Y, years of age.
P1 bone structure formed concurrently with precocial ambulatory weight‐bearing. Animals increased continuously in mass from birth through adulthood (τ 1/2 = 5.1 ± 1.2 mo, Figure 4A). To determine how bone morphometry changes with loading, we normalized all P1 cortical and trabecular outcomes by animal body mass (Figures 4B–F & S1). Most body mass‐normalized cortical and trabecular outcome measures plateaued rapidly: Ct.Ar (τ 1/2 = 0.9 ± 0.4 mo, Figure 4B), cortical TMD (τ 1/2 = 1.4 ± 0.5 mo, Figure 4C), BV/TV (τ 1/2 = 1.5 ± 0.5 mo, Figure 4E), Tb.Th (τ 1/2 = 1.3 ± 0.5 mo, Figure 4F), Tt.Ar (τ 1/2 = 2.2 ± 1.0 mo, Figure S1A), Ct.Ar/Tt.Ar (τ 1/2 = 2.3 ± 1.0 mo, Figure S1B), Ct.Th (τ 1/2 = 0.8 ± 0.5 mo, Figure S1C), Tb.N (τ 1/2 = 0.9 ± 0.3 mo, Figure S1D), Tb.Sp (τ 1/2 = 0.9 ± 0.4 mo, Figure S1E), and trabecular TMD (τ 1/2 = 1.0 ± 0.4 mo, Figure S1F). In contrast, the body mass‐normalized polar moment of inertia (*J/*mass) was constant over time (Figure 4D). These data suggest that while bone accrual was dominated by animal growth rather than changes correlated with ambulatory loading, the morphodynamics of cross‐sectional distribution, indicative of structure resistance to bending and torsional loading, were associated with body mass and thus ambulatory loading.
Morphodynamic changes of P1 cross‐sectional distribution correlate with ambulatory loads. (A) Animal mass throughout developmental time. Body mass normalized (B) Cortical bone area (Ct.Ar), (C) Cortical tissue mineral density (TMD), (D) Polar moment of inertia (J), (E) Trabecular bone volume fraction (BV/TV), and (F) Trabecular thickness (Tb.Th). Prefix “Norm” indicates outcome is normalized to animal mass. Data graphed as individual data points vs. time fit with a one‐phase decay model. Gray bands indicate 95% confidence interval for each curve for the full dataset. Datapoint color indicates animal sex: Male = teal, female = purple. Goodness of fit (R 2) and half‐life (τ 1/2) are reported for each graph.
Metacarpal postnatal development dynamics
2.2
Synostosis of the metaphyseal diaphyses was uniformly present in neonates, indicating fusion during fetal development (Figure 5A). Cortical fusion was only observed in the diaphyseal region of the metacarpals and preserved the structure of two distinct distal physes. Additionally, metacarpal bones maintained distinct lateral and medial trabecular networks in the diaphysis and metaphysis.
Metacarpal cortices fuse prior to birth and distal metacarpal trabecular bone grows through trabecular thickness expansion. (A) 3D microCT reconstructions of cortices with a virtual slice through the transverse plane, showing cortical fusion prior to birth. Scale bars = 5 mm. (B) 3D microCT reconstructions of metacarpal bones with a virtual slice through the mid‐coronal plane. Scale bars = 5 mm. (C) Trabecular number (Tb.N), (D) Trabecular thickness (Tb.Th), (E) Bone volume fraction (BV/TV), (F) Trabecular spacing (Tb.Sp), and (G) Tissue mineral density (TMD) throughout developmental time, fit with a one‐phase decay model. Gray bands indicate 95% confidence interval for each curve for the full dataset. Datapoint color indicates animal sex: Male = teal, female = purple. Goodness of fit (R 2) and half‐life (τ 1/2) are reported for each graph. D, days of age; M, months of age; Y, years of age.
Bone formation in the epiphyseal metacarpal trabecular bone occurred by increasing trabecular thickness, rather than increasing trabecular number (Figure 5B–G). Tb.N changed minimally during postnatal metacarpal development (Figure 5C), while Tb.Th (τ 1/2 = 7.9 ± 2.2 mo, Figure 5D) increased continuously over 3.5 years, suggesting bone accrual on existing trabecular networks over time. Simultaneously, the trabecular bone matrix increased in mineral density, TMD (τ 1/2 = 5.7 ± 1.7 mo, Figure 5F). Despite limited changes in Tb.N, the continuously increasing Tb.Th was sufficient to increase trabecular BV/TV, but not Tb.Sp, over postnatal development (τ 1/2 = 6.9 ± 3.1 mo, Figure 5E,F). These findings suggest that the development of the epiphyseal metacarpal trabecular microarchitecture may feature mineral apposition on trabecular networks present at birth, rather than de novo trabecular formation.
Epiphyseal bone fusion dynamics in the metacarpal bones
2.3
Diaphyseal synostosis of the metacarpal bones occurred prenatally, but epiphyseal synostosis occurred postnatally. Diaphyseal synostosis could be observed as early as 3D (Figures 5A and 6A), suggesting this fusion event occurred prior to birth. In contrast, synostosis of the epiphyses distal to the growth plate did not initiate until after 3 months (Figure 6A,B). We quantified the extent of epiphyseal synostosis as the fusion length, which increased continuously between 3 months and 12 months. The distal metacarpal physis closed between 12 months and 3.5 years; thus, it was not possible to measure fusion length past 12 months. At 3.5 years, the metacarpal bone remodeled the growth plate into a mature network of cancellous bone.
Metacarpal epiphyses are separated by a non‐cartilaginous fibrous template at birth, but fuse by fibrocartilaginous endochondral ossification. (A) 3D microCT reconstructions of metacarpal bones with a virtual slice through the mid‐coronal plane. Yellow line indicates extent of epiphyseal fusion (i.e., “fusion length”). Scale. bars = 5 mm. (B) Quantification of the epiphyseal fusion length throughout developmental time. Datapoint color indicates animal sex: Male = teal, female = purple. (C) Safranin o/fast green histological micrographs of metacarpal bones cut through the mid‐coronal plane. Scale bars = 100 μm. (D) High‐magnification micrographs of the distal metacarpus. Black arrowheads indicate synonstosis via fibrocartilaginous endochondral ossification. Scale bars = 500 μm. (E) High‐magnification micrographs of articular cartilage region of the metacarpus. Scale bars = 100 μm. D, days of age; M, months of age; Y, years of age.
Histological analyses showed how the distinct metacarpal epiphyses underwent synostosis. At 3 days, the metacarpus had two distinct growth plates (i.e., physes) connected by a non‐ossified, non‐cartilaginous, periosteum‐like fibrous tissue (Figure 6C,D). Between 3 days and 1.5 months of age, the two physes joined, and the previous fibrous tissue exhibited robust glycosaminoglycan (GAG) deposition, which continued through the epiphyseal region. Between 1.5 and 3 months, endochondral ossification initiated in the interstitial fibrocartilaginous matrix, starting at the border with the conjoined physes. This process led to epiphyseal synostosis by 9 months of age. From 9 to 12 months old, the metacarpals continued longitudinal growth. Throughout this epiphyseal synostosis, the physes maintained a fibrocartilaginous bridge, devoid of chondrocyte columns between physes. Between 12 months and 3.5 years, the physis closed, and the distal metacarpal bone remodeled to form a singular continuous network of cancellous bone.
The articular cartilage also matured over postnatal development. At 3 days, the distal portion of the metacarpal bone exhibited a coherent articular cartilage surface with robust GAG content (Figure 6E). The articular cartilage matured and condensed throughout postnatal development, forming superficial, transitional, and deep zones by 3 months of age and reducing GAG content by 12 months of age.
DISCUSSION
3
Here, we characterized the dynamics of postnatal bone development and epiphyseal synostosis in the caprine autopod from birth through adulthood. P1 bone accrual was driven by growth patterning, while cross‐sectional structural morphodynamics were maintained as ambulatory loading increased. While P1 distal trabeculae increased in thickness and number, metacarpal epiphyseal trabecular bone increased primarily by the thickening of pre‐existing trabeculae. The two bones that fuse to form the caprine metacarpus underwent diaphyseal synostosis prenatally, but epiphyseal synostosis postnatally, prior to growth plate closure, through fibrocartilaginous endochondral ossification. Together, these data provide new insights into precocial bone development under ambulatory loading. Our findings may guide future caprine mechanobiology studies to mechanistically understand bone development and adaptation to ambulatory loading.
P1 cortical bone development was driven by both patterned growth and ambulatory loading. Cortical bone rapidly condensed within months, followed by continuous cortical bone accrual through adulthood. Notably, the body mass‐normalized polar moment of inertia (J/mass) was maintained over developmental time, suggesting morphodynamic changes in structural distribution to resist bending deformation by ambulatory loading. These findings demonstrate the early functional capacity of the precocial skeleton, which persists into adulthood. The precocial skeleton develops so robustly that it can even develop normally after preterm birth and reduced birthweight.19 In humans, which are altricial, peripubertal physical activity increases bone formation and establishes bone structural properties that maintain bone mass and strength,20, 21 which persist into adulthood.22, 23 These findings support the notion that early and peripubertal impact loading can preserve adult bone strength and reduce fracture risk.24, 25 Future studies can use the caprine model system to determine how altered perinatal mechanical loading impacts long‐term skeletal health.
Trabecular architecture matured in P1 and metacarpal bones at different rates, but both were significantly different from altricial developmental dynamics. Trabecular bone only makes up 20% of total bone mass,26 but the trabecular microarchitecture is crucial for determining bone mechanical properties.27, 28 The developing caprine autopod of the lower forelimb is useful for studying adaptive trabecular bone growth because precocial species experience progressive biomechanical loading from gestation, through ambulation at birth, and into adulthood. Metacarpal distal epiphyseal BV/TV increased at a slower rate than P1 distal BV/TV, likely because metacarpal trabecular bone formed by trabecular bone apposition, rather than de novo trabecula formation. Thus, the metacarpal trabecular network may have formed in utero and only developed by bone accrual postnatally. Despite different mechanisms of development, trabecular BV/TV increased continuously after birth in the P1 and metacarpal bones, which is distinct from altricial trabecular bone dynamics. Altricial species experience a reduced loading state from birth until the onset of ambulation, which may explain the loss in trabecular bone volume fraction during early postnatal development.29 Once humans begin walking, trabecular BV/TV continuously increases coincidentally with progressive loading.30, 31 These post‐ambulatory BV/TV increases mirror the progressive increases in trabecular bone observed in our study. Our findings contribute to the growing body of literature describing how precocial animals adapt to the biomechanical loading of ambulation.9, 10, 11
The epiphyseal synostosis of the caprine metacarpal bones is distinct from other known mechanisms of endochondral bone formation or bone fusion. Here we show that the metacarpal physes and epiphyses are initially separated by a non‐cartilaginous fibrous template, which becomes chondrogenic and subsequently undergoes fibrocartilaginous endochondral ossification. This rudimentary fibrous tissue is distinct from the cartilaginous rudiments that give rise to endochondral primary bone development,32 or the cartilage template that initiates the fibrocartilaginous enthesis.33, 34, 35 The epiphyseal synostosis is also distinct from the fusion of flat bones of the skull. Like the metacarpal epiphyses, the sutures of the skull feature a flexible fibrous tissue, which allows for brain growth and development. However, sutures fuse by intramembranous ossification, without a cartilaginous intermediate.36 Synostosis of metacarpal epiphyseal bone is perhaps most analogous to endochondral bone fracture healing.37, 38 The progression of fibrous to cartilaginous to bone tissue is like fracture healing, though fracture features an acute inflammatory phase and less robust fibrous tissue formation. Biomechanical stimuli are key regulators of endochondral ossification,5, 6 suture fusion,39, 40 enthesis development,33 and fracture healing41, 42, 43, 44, 45 and may play a role in metacarpal epiphyseal synostosis. Interrogating the mechanical basis of metacarpal epiphyseal synostosis could lead to novel insights into the underlying mechanisms.
Together with a growing body of literature on precocial bone formation, these development dynamics data may form a foundation on which to uncover how mechanical forces regulate skeletal development, disease, and regeneration. A key limitation of this study is that we could not account for sex differences. To enable sufficient numbers and ages, given housing and breeding constraints,46 we used male and female goats from the Translational Research Unit (TRU) goat herd at University College Dublin Lyons Research Farm. This led to different numbers of male and female goats in each group. Further study will be required to directly interrogate sex as a biological variable among animals of more homogenous backgrounds.47
EXPERIMENTAL PROCEDURES
4
Study design
4.1
We performed bone morphometry on the autopods of the lower forelimbs of 44 Saanen goats at the following ages: 3 days (3D, N = 6 male, N = 3 female), 1.5 months (1.5M, N = 3 male, N = 3 female), 3M (N = 1 male, N = 4 female), 6M (N = 6 female), 9M (N = 3 male, N = 3 female), 12M (N = 3 male, N = 3 female), and 3.5 years (3.5Y, N = 6 female). Each animal was considered an experimental unit, and goats were assigned to groups in cohorts. Goats were continually assessed by a veterinarian, and only animals in good physical health were included in the study. At the specified timepoints, animals were euthanized, and the phalanges 1 (P1) and metacarpal bones were harvested from autopods of the lower forelimbs for microcomputed tomography (μCT) and histological analyses.
Animal husbandry and care
4.2
This animal experiment complied with the ARRIVE guidelines. Ethical evaluation and approval were provided by the Animal Research Ethics Committee (AREC‐21‐22) of University College Dublin (UCD) and the Lyons UCD Research Farm Animal Welfare Board (Health, Husbandry and Monitoring plans: 201,907). Animals were selected at predetermined age points from the Translational Research Unit (TRU) goat herd at University College Dublin Lyons Research Farm (Breeding license AREC‐22‐408,929). In principle, all animals genetically originated from the same high health closed goat herd of our sole supplier.
The TRU Saanen goat herds were kept under normal goat husbandry conditions. Herds had loose housing and pasture access when weather conditions were appropriate. Daily health checks of the herd were performed by appropriately trained husbandry staff, with weather conditions and barn climate parameters documented on the daily health check sheets. Animals were reared with their respective mothers when available or at a lambing bar with milk replacer to suckle milk until weaning at approximately 60–90 days (depending on body condition score and weight). Ad libitum concentrates and hay/silage were provided from approximately 14 days of age. After goats were 3 months, hay/silage remained ad libitum, but concentrates were provided based on their daily requirements and weight gain to prevent overfeeding. Veterinary staff of the TRU performed weekly monitoring health checks on the goat herds and were available 24/7 in case a veterinary consultation was required.
When animals reached their predetermined age endpoint, a normal general health check and orthopedic assessment were conducted. Only animals that met these requirements were included in the study. Included animals underwent humane euthanasia under sedation with a barbiturate overdose. Following euthanasia, the P1 and metacarpal bones were harvested for microcomputed tomography (μCT) and histological analyses. Samples were fixed using 10% neutral buffered formalin (NBF) for 3 days, then 5% NBF for 3 weeks. Fixed samples were washed with 1× phosphate buffered saline (PBS), then frozen at −20°C in a solution of 30% sucrose (w/v) in 1× PBS prior to analyses.
Microcomputed tomography
4.3
The night before scanning, frozen samples were thawed overnight at 4°C. Thawed samples were wrapped in PBS‐soaked gauze and scanned using a SCANCO μCT 45 desktop microCT scanner (SCANCO Medical, Wangen‐Brüttisellen, Switzerland). X‐ray images were acquired using an X‐ray intensity of 145 μA, an energy of 55 kVp, an integration time of 400 ms, and a resolution of 10.4 μm. Reconstructed scans were semi‐automatically contoured and analyzed using the SCANCO microCT image analysis interface (SCANCO Medical) according to morphometric standards.48
Regions of interest using SCANCO 3D morphometry software, then SCANCO software calculated cortical and trabecular outcomes using a model‐independent algorithm. Cortical tissue cross‐sectional area (Tt.Ar) and cortical bone area (Ct.Ar) were calculated using average cross‐sectional geometries for each contoured slice. Tissue mineral density (TMD) is calculated from the average attenuation value of the bone tissue and does not include non‐bone voxels. Polar moment of inertia (J) measures the distribution of the cortical bone about the central axis. Trabecular thickness (Tb.Th) is the average thickness of trabeculae. It is calculated by determining the diameter of the largest possible sphere that can be fitted through each voxel that is completely contained within the object, then averaging these diameters. Trabecular separation (Tb.Sp) represents the difference between trabeculae. Like Tb.Th, Tb.Sp is calculated using a sphere fitting model to the background. Trabecular number (Tb.N) is the average number of trabeculae per unit length. It is computed as the inverse of the mean distance between the mid‐axes of the structure. Schematic representations of these standard microCT outcomes are shown in Figure 1C.
For P1 bones, approximately 200 slices of the midshaft cortical region were contoured to measure cortical tissue cross‐sectional area (Tt.Ar), cortical bone area (Ct.Ar), cortical area fraction (Ct.Ar/Tt.Ar), cortical thickness (Ct.Th), tissue mineral density (TMD), and polar moment of inertia (J). Additionally, the entire P1 distal trabecular region was contoured to measure trabecular bone volume fraction (BV/TV), trabecular number (Tb.N), trabecular thickness (Tb.Th), and trabecular separation (Tb.Sp). For each goat, the cortical and trabecular outcome measurements were averaged for the medial and lateral P1 bones, so each goat was an experimental unit.
For metacarpal bones, BV/TV, Tb.N, Tb.Th, and Tb.Sp were calculated for the combined distal trabecular network in both epiphyses. Additionally, the maximum epiphyseal fusion length was calculated for each metacarpal bone using the mid‐sagittal virtual slice numbers for the initiation of epiphyseal synostosis and the physis.
Histology
4.4
Representative P1 and metacarpal samples (N = 3 per timepoint) were used for paraffin histology. P1 samples were decalcified for 3.5 weeks using Formical‐4 Decalcifier (StatLab Medical Products, McKinney, Texas, USA), with weekly reagent changes. Metacarpal samples were decalcified for 4 weeks using Formical‐4, then an additional 2 weeks using Formical‐2000 (StatLab Medical Products), with weekly reagent changes. Decalcified samples were trimmed using a Tissue‐Tek® Accu‐Edge® Trimming Knife (Electron Microscope Sciences, Hatfield, PA, USA) and processed for paraffin histology using a Thermo Scientific Excelsior AS Tissue Processor (Thermo Fisher Scientific, Waltham, MA). Processed samples were paraffin‐embedded and sectioned to 10 μm using a Leica RM 2030 Microtome (Leica, Bala Cynwyd, PA, USA). Sections were stained for safranin‐O/fast green using standard protocols. Stained sections were imaged using a ZEISS Axio Scan.Z1 Slide Scanner (ZEISS, Oberkochen, Germany).
Statistical analysis
4.5
The “pwr2” package in R (Indianapolis, IN, USA) was used to conduct a power analysis for large effect sizes (α = 0.05, β = 0.20, f = 0.4–0.6). GraphPad Prism software Version 10.3.1 (GraphPad Software, San Diego, CA, USA) was used to conduct all statistical analyses. A one‐way analysis of variance (ANOVA) with Tukey's multiple comparison test was used to determine significant differences in all outcome measurements over developmental time. Prism was also used to fit all outcome measurements with a one‐phase exponential decay model (y=span*e−kt+plateau). Graphical results are reported as a one‐phase decay model with the goodness of fit (R ^2^) and half‐life (τ 1/2). τ 1/2 is the amount of developmental time, in months, for each outcome to achieve 50% of its maximum value.
CONFLICT OF INTEREST STATEMENT
The authors declare that they have no competing interests.
Supporting information
FIGURE S1. Morphodynamic changes of P1 cross‐sectional distribution correlate with ambulatory loads. Body mass normalized (A) Cortical tissue cross‐sectional area (Tt.Ar), (B) Cortical area fraction (Ct.Ar/Tt.Ar), (C) Cortical thickness (Ct.Th), (D) Trabecular number (Tb.N), (E) Trabecular spacing (Tb.Sp), and (F) Trabecular tissue mineral density (TMD). Prefix “Norm” indicates outcome is normalized to animal mass. Data graphed as individual data points versus time fit with a one‐phase decay model. Gray bands indicate 95% confidence interval for each curve for the full dataset. Datapoint color indicates animal sex: male = teal, female = purple. Goodness of fit (R ^2^) and half‐life (τ 1/2) are reported for each graph.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wolff J . Das gesetz der transformation der knochen. Dtsch Med Wochenschr. 1893;19(47):1222‐1224. doi:10.1055/s-0028-1144106 · doi ↗
- 2Frost HM . Wolff's law and bone's structural adaptations to mechanical usage: an overview for clinicians. Angle Orthod. 1994;64(3):175‐188. doi:10.1043/0003-3219(1994)064<0175:WLABSA>2.0.CO;2 8060014 · doi ↗ · pubmed ↗
- 3Nowlan NC . Biomechanics of foetal movement. Eur Cell Mater. 2015;29:1‐21. doi:10.22203/ecm.v 029a 01 25552425 · doi ↗ · pubmed ↗
- 4Beall MH , van den Wijngaard JPHM , van Gemert MJC , Ross MG . Amniotic fluid water dynamics. Placenta. 2007;28(8–9):816‐823. doi:10.1016/j.placenta.2006.11.009 17254633 · doi ↗ · pubmed ↗
- 5Kozhemyakina E , Lassar AB , Zelzer E . A pathway to bone: signaling molecules and transcription factors involved in chondrocyte development and maturation. Development. 2015;142(5):817‐831. doi:10.1242/dev.105536 25715393 PMC 4352987 · doi ↗ · pubmed ↗
- 6Collins JM , Lang A , Parisi C , et al. YAP and TAZ couple osteoblast precursor mobilization to angiogenesis and mechanoregulation in murine bone development. Dev Cell. 2024;59(2):211‐227.e 5. doi:10.1016/j.devcel.2023.11.029 38141609 PMC 10843704 · doi ↗ · pubmed ↗
- 7Breeland G , Sinkler MA , Menezes RG . Embryology, bone ossification. Stat Pearls. Stat Pearls Publishing; 2025.30969540 · pubmed ↗
- 8Gardner E , Gray DJ . The prenatal development of the human femur. Am J Anat. 1970;129(2):121‐140. doi:10.1002/aja.1001290202 5473773 · doi ↗ · pubmed ↗