The mitochondrial aspartate transporter Ucp4a regulates muscle aging and animal lifespan in Drosophila melanogaster
Minwoo Baek, Wijeong Jang, Changsoo Kim

TL;DR
A mitochondrial protein in fruit flies affects muscle aging and lifespan by reducing protein buildup.
Contribution
Ucp4a, a mitochondrial aspartate transporter, is shown to regulate muscle aging and lifespan in Drosophila.
Findings
Flies with Ucp4a mutations live longer.
Reducing Ucp4a in muscles removes protein aggregates linked to aging.
Lower cytosol aspartate levels may trigger a signaling process extending lifespan.
Abstract
Mild distress of mitochondria extends animal lifespan, yet the underlying mechanisms are not completely understood. Here we screened mitochondrial proteins for effects on longevity and found that flies mutant in Uncoupling protein 4a (Ucp4a), which encodes a mitochondrial aspartate transporter, have extended lifespans. Tissue-specific experiments revealed knockdown of Ucp4a in muscles, but not neurons, fat, or intestine, to extend lifespan and also eliminate polyubiquitinated protein aggregates, which accumulate with aging and are associated with lifespan. These findings suggest a retrograde mitochondrial signaling process initiated by reduced cytosol aspartate level culminates in muscle protein aggregate removal and lifespan extension.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
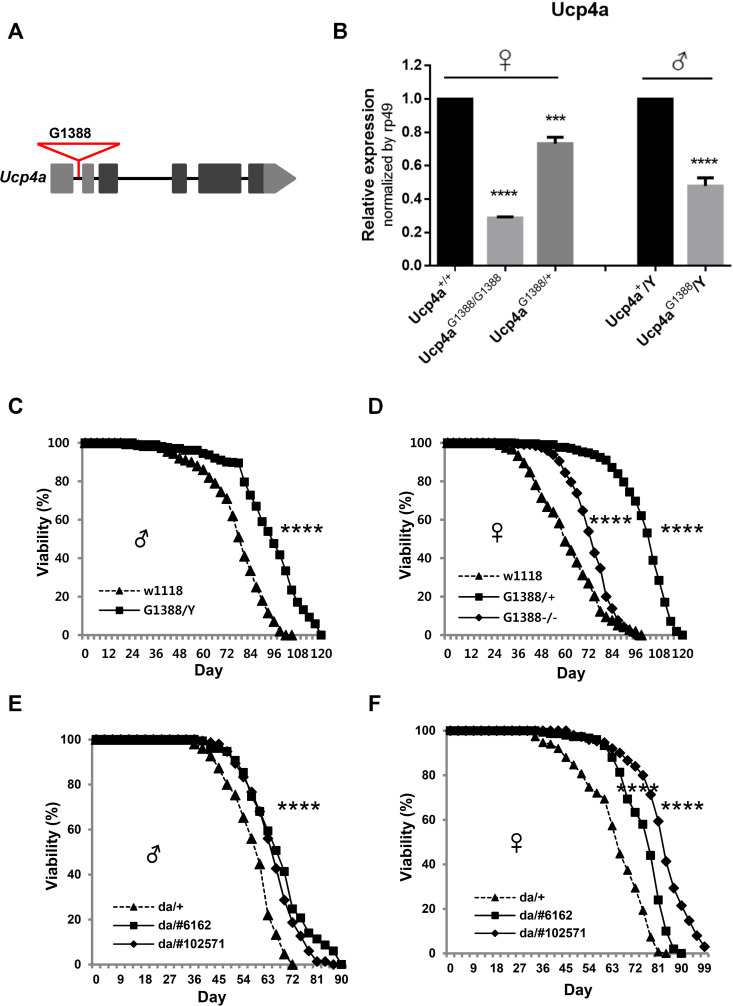 Fig 1
Fig 1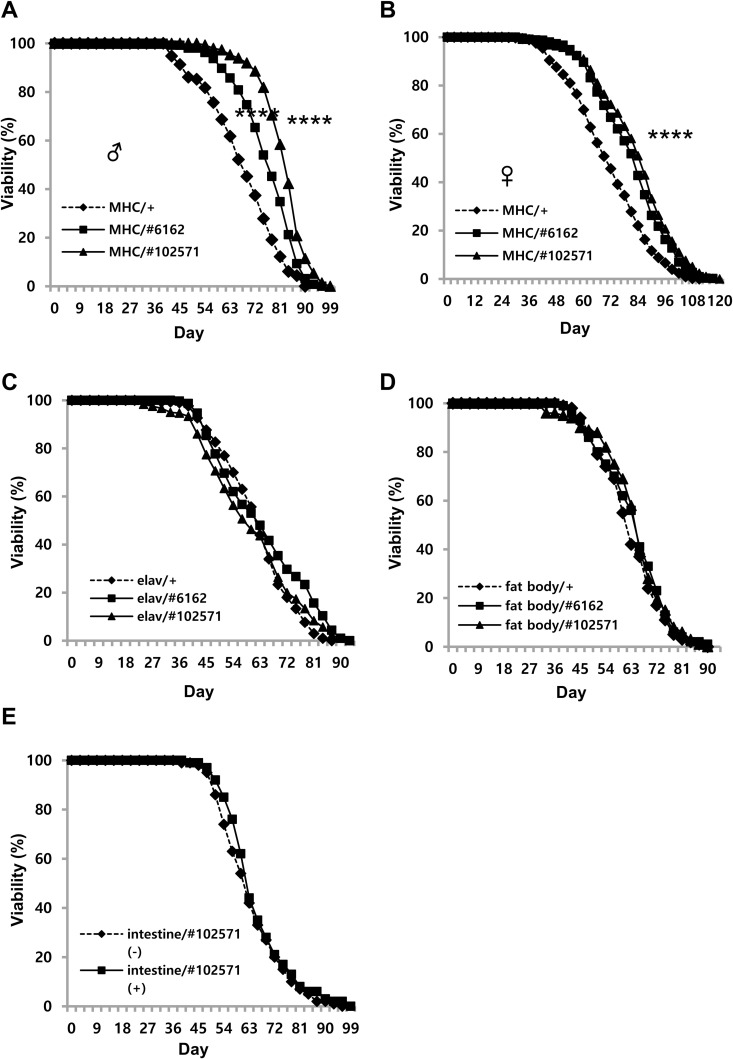 Fig 2
Fig 2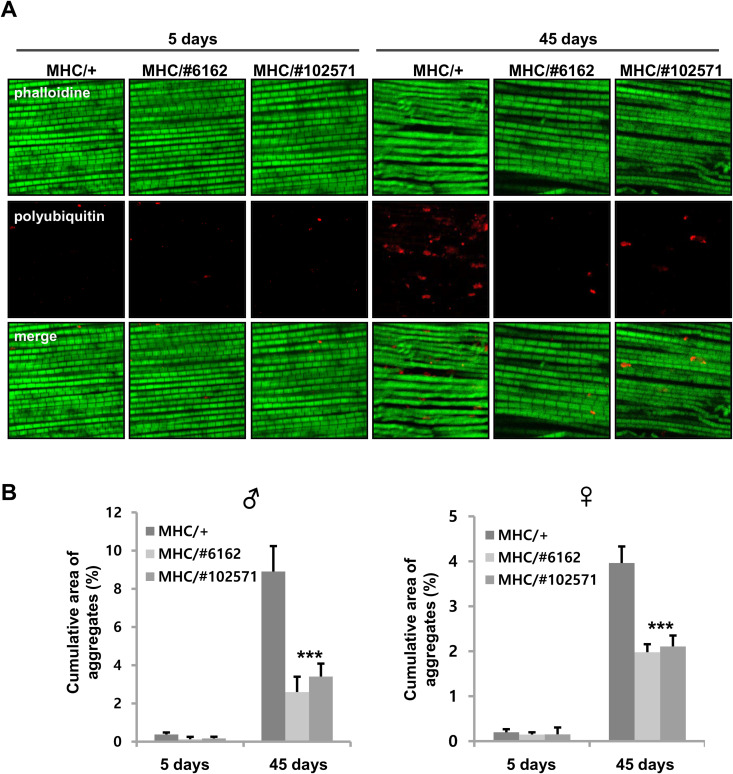 Fig 3
Fig 3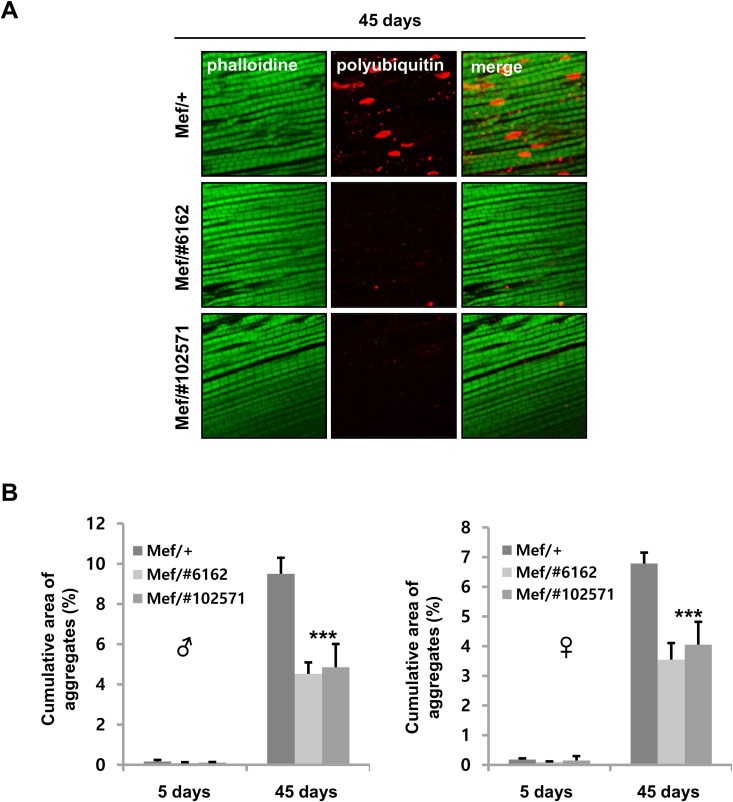 Fig 4
Fig 4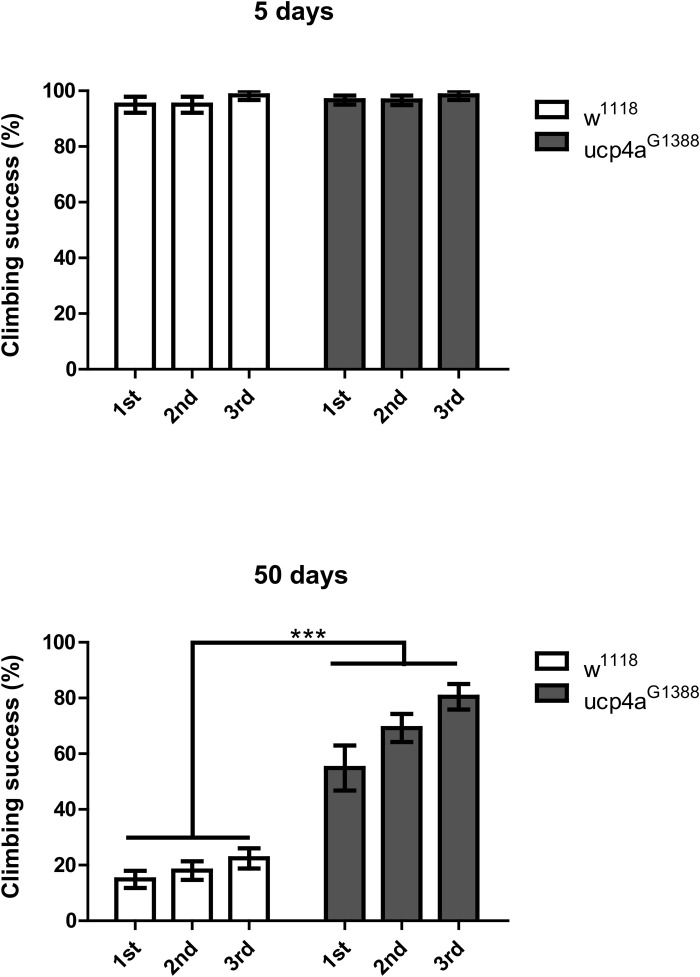 Fig 5
Fig 5- —http://dx.doi.org/10.13039/501100003725National Research Foundation of Korea
- —national research foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms · Mitochondrial Function and Pathology · Muscle Physiology and Disorders
Introduction
Mitochondria are subcellular organelles that utilize an electron transport chain (ETC) to produce cellular energy and also synthesize numerous metabolites that efflux to the cytosol [1–3]. Mild knockdown of mitochondrial ETC proteins prolongs lifespan, a phenomenon that has been observed in diverse organisms including C. elegans, Drosophila, and mice [4–7]. In mammalian cells, ETC perturbation or mitochondrial distress represses the mechanistic target of rapamycin complex 1 (mTORC1) pathway through activation of the transcription factor ATF4 [2,8–12]. Studies in Drosophila muscle have found ETC perturbation to repress systemic insulin signaling through expression of ImpL2, an inhibitor of Drosophila insulin-like peptides (Dilps) [13,14]. Repression of either mTORC1 or insulin signaling is established to extend lifespan [15–19]; however, the mechanisms underlying this mitochondrial-distress-mediated life extension are not yet completely understood.
Aspartate (asp), a proteogenic amino acid, is synthesized in the mitochondrial matrix from glutamate and oxaloacetic acid (OAA), a tricarboxylic acid (TCA) cycle metabolite [20]. Of note, asp synthesis requires integral mitochondrial function. In mammalian cells, treatment with an ETC inhibitor depletes asp due to impairment of NADH flux, which is required for integrity of the TCA cycle [12,21]. Perturbation of asp synthesis impairs cell proliferation, and in endothelial cells impairs the cytosolic mTORC1 pathway [12,21–23]. Here we show that Uncoupling protein 4 a (Ucp4a), a mitochondrial asp transporter [24], regulates muscle aging and lifespan in Drosophila.
Materials and methods
2.1. Fly husbandry
Ucp4a^G1388^, da-, elav-, MHC-, mef-, cad- and C574-Gal4 lines were from the Bloomington Stock Center. UAS-Ucp4a RNAi (#6162 and #102571) were provided by the VDRC Stock Center. Fly culture and crosses were performed on standard fly food containing yeast, cornmeal, and sugar, and the flies were raised at 25 °C. The w^1118^ line was used as a wild-type genetic background.
2.2. Lifespan assay
All flies, including Ucp4a^G1388^, Gal4 lines and UAS-ucp4a RNAi #6162, #102571, were backcrossed to the w^1118^ line for six generations to minimize genomic background differences. Flies (1 ~ 2 days old) were collected by anesthetization with CO_2_ gas. All flies were kept in a humidified (50%), temperature-controlled incubator with a 12 h on/off light cycle at 25°C. Flies (20 per vial) were transferred to fresh food every three days and scored for death. Significant differences in lifespan were assessed using the log-rank test method in GraphPad Prism 5.
2.3. Capillary feeding (CAFE) assay
Quantification of food uptake was performed by capillary feeding assay as described previously [25] with slight modification. Briefly, five flies were placed in vials with wet glass paper for a water source and a capillary food source (5% sucrose, 5% yeast extract). Feeding was monitored every six hours, at which time fresh capillaries were placed and the feeding amount calculated.
2.4. Quantitative RT-PCR
Total RNA was extracted with TRIzol reagent (Invitrogen) following manufacturer protocols. Samples were treated with DNase, and the RNA concentrations measured and equalized. Complementary DNA was synthesized using a cDNA synthesis kit (Bioneer). Quantitative RT-PCR was performed using QuantiTech SYBR Green (Qiagen), and DNA amount was monitored with a Rotor-Gene Q (Qiagen). Equalized amplicons of Rp49 were used to normalize. Primer sequences were as follows: Ucp4a, 5’-TTCGCCTGCACTTACATCGT-3’and 5’-CACCGCTATAGACGACGTGT-3’.
2.5. Immunostaining
Total polyubiquitinylated protein and the sarcomere structure of the indirect flight muscle were evaluated as previously described [26]. Briefly, adult flies were fixed with 4% paraformaldehyde in PBS for 1 h, embedded in OCT compound (Fisher Scientific), and frozen with liquid nitrogen. Samples were then processed by a cryomicrotome (Leica). After post-fixing with 4% paraformaldehyde in PBS, samples were permeabilized with 0.2% Triton X-100 buffer in PBS and blocked with 5% BSA solution in PBS for 1 h. Afterwards, the samples were incubated with anti-ubiquitinylated proteins antibody (Millipore, 1:200) and phalloidin AlexaFluor-488 (Invitrogen 1:1000) overnight at 4 °C. Anti-mouse IgG Alexa Flore 568 (Invitrogen) was used as the secondary antibody (1:1000). Samples were mounted with Prolong Gold Antifade Reagent with DAPI (Invitrogen) and imaged with a Leica confocal microscope (Leica); finally, the images were analyzed by the ImageJ software.
2.6. Locomotive assay
Flies (1 ~ 3 days old) were collected by anesthetization with CO_2_ gas. Twenty male flies per vial, which were kept in a humidified (50%) incubator at 25°C, were transferred to fresh food every three days. For locomotive assays, flies (5, 10, 30, 45, 50, 60 day old) were transferred to an empty vial directly from the food vial, idled for 10 min, and then tapped to the bottom. The number of flies that climbed to reach a 7 cm cutline within 10 sec was counted. Data are shown as mean±SEM and were analyzed using GraphPad Prism 6.0.
2.7. Data analysis
The statistical significance of differences in standard error of the mean (SEM) or standard deviation (SD) of the mean values was obtained using two-way ANOVA with Sidak’s multiple comparisons test. Kaplan-Meier survival curves was used for lifespan and survival analysis with log-rank tests. Prism software (GraphPad version 6.0, San Diego, USA) was used to calculate p-values. Results were considered statistically significant at levels *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
Results
We carried out a screening that identified a longevity line with an EP-element [27] inserted into the first intron of Ucp4a (hereafter we term the Ucp4a mutant line as Ucp4a^G1388^) (Fig 1A). qRT-PCR showed that Ucp4a mRNA was reduced in Ucp4a^G1388^ hemizygous males (Ucp4a^G1388^/Y), and heterozygous (Ucp4a^G1388^/^+^) and homozygous females (Ucp4a^G1388^/Ucp4a^G1388^ or Ucp4a ^-^/^-^) (Fig 1B). ucp4a hemizygous males (Ucp4a^G1388^/Y) exhibited longer lifespan, with an average 18.5% extension (Fig 1C), and ucp4a homozygous females (Ucp4a^G1388^/Ucp4a^G1388^ or Ucp4a ^-^/^-^) exhibited a comparable 19.0% increase (Fig 1D). Notably, ucp4a heterozygous females (Ucp4a^G1388^/^+^ or Ucp4a ^+^/^-^) exhibited a dramatic lifespan increase of 66.7% (Fig 1D).
*Prolongation of lifespan by mutation and knockdown of Ucp4a.(A) The Ucp4a mutant line. G1388 is an EP element inserted into the first intron of the Ucp4a gene. Gray and black boxes denote noncoding and coding sequences, respectively. (B) qRT-PCR of Ucp4a normalized to rp49. Error bars indicate ±SEM of three independent experiments. Student’s t-test, ***, p < 0.001. ****, p < 0.0001. (C,D) Viability of Ucp4aG1388 hemizygous mutant (ucp4aG1388/Y) and w1118 (genetic control for EP line) male flies (C), and Ucp4aG1388 heterozygous mutant (ucp4aG1388/+), homozygous mutant (ucp4aG1388/ ucp4aG1388) and w1118 female flies (D). W1118 denotes W1118/Y. G1388/Y denotes Ucp4aG1388/Y. G1388/+ denotes Ucp4aG1388/+. G1388-/- denotes Ucp4aG1388/Ucp4aG1388. n = 400 for each curve. ****, p < 0.0001, log rank test. (E,F) Viability of Ucp4a whole-body knockdown male (E) and female (F) flies. da/#6162 and da/#102571 indicate da-Gal4 > UAS-Ucp4a RNAi #6162 and #102571, respectively. da/+ indicates da-Gal4/+ , used as control. n = 400 for each curve. ***, p < 0.0001, log rank test.
Knocking down of Ucp4a in whole cells with Ucp4a RNAi driven by da-Gal4, a ubiquitous Gal4 driver, in two distinct RNAi lines (da-Gal4 > UAS-Ucp4a RNAi #6162, #102571) extended lifespan in males and females by 20% and 30%, respectively (Fig 1E,F). Subsequently, elav-, MHC-, cad-, and c574-Gal4 tissue-specific drivers were used to drive Ucp4a RNAi in neurons, muscle, fat body, and intestine, respectively. Interestingly, the muscle-specific Gal4 driver was effective in extending male lifespan by 21.7% (Fig 2A), comparable to the ubiquitous driver (Fig 1E,F). Likewise, in females, the muscle Gal4 driver extended lifespan by 26%, similar to the 31% of the whole-body driver (Fig 2B). Other Gal4 drivers did not increase longevity (Fig 2C–E). UAS-Ucp4a #6162 was slightly less effective than Ucp4a RNAi #102571 (Figs 1F,2A). Taken together, these data suggest that muscle is the source of the Ucp4a-mediated effect on lifespan.
*Prolongation of lifespan by tissue-specific knockdown of Ucp4a.(A,B) Viability of Ucp4a muscle-specific knockdown male (A) and female (B) flies. MHC/+ denotes MHC-Gal4/+ . MHC/#6162 denotes MHC-Gal4/+ ; Ucp4a RNAi #6162. MHC/#102571 denotes MHC-Gal4/+ ; Ucp4a RNAi #102571. n = 400 for each curve. ***, p < 0.001, log rank test. (C) Viability of Ucp4a knockdown male by neuronal Gal4 (elav-Gal4). Elav/+ denotes elav-Gal4/+ . Elav/#6162 denotes elav-Gal4/+ ; ucp4a RNAi #6162. Elav/#102571 denotes elav-Gal4/+ ; ucp4a RNAi #102571. (D) Viability of Ucp4a knockdown male by fat body Gal4 (cad-Gal4). Fat body/+ denotes cad-Gal4/+ . Fat body/#6162 denotes cad-Gal4/+ ; ucp4a RNAi #6162. Fat body/#102571 denotes cad-Gal4/+ ; ucp4a RNAi #102571. (E) Viability of Ucp4a knockdown male by drug-inducible intestinal Gal4 (c574-Gal4). Intestine/#102571 denotes c574-Gal4/+ ; ucp4a RNAi #102571. (-), (+) denote administration of drug (RU486, 50 µg/ml) in food.
An association of muscle aging with lifespan has previously been demonstrated [13,28,29], prompting us to investigate whether Ucp4a reduction in muscle delays muscle aging. Accumulation of polyubiquitinated proteins is a hallmark of muscle aging [29]. We confirmed polyubiquitinated aggregates to be increased in normal aged muscles (Fig 3). Meanwhile, the ucp4a mutant (S1 Fig) and muscle-specific Ucp4a knockdown lines (MHC-Gal4 and mef-Gal4) exhibited reduced polyubiquitinated aggregates in aged muscles (Figs 3,4).
*Polyubiquitinated protein aggregates in muscle with age.(A) Confocal images of adult male thorax (indirect flight muscle) sections co-stained with polyubiquitinated protein antibody (red) and phalloidin (green). MHC/+ denotes MHC-Gal4/+ . MHC/#6162, 102571 indicates MHC-Gal4 > UAS-Ucp4a RNAi #6162, #102571, respectively. (B) Cumulative area of aggregates normalized to total visualized area from Fig 3A. Error bars indicate SD for n = 10 muscles. **, p < 0.001, two-way ANOVA with Sidak’s multiple comparisons test.
*Polyubiquitinated protein aggregates in muscle with age.(A) Immunohistochemical staining of adult thorax (indirect flight muscle) sections co-stained with polyubiquitinated protein antibody (red) and phalloidin (green). 45-day-old male flies. Mef/+ (control) denotes Mef-Gal4/+ . Mef/#6162, #102571 indicates Mef-Gal4 > UAS-Ucp4a RNAi #6162, #102571. (B) Cumulative area of aggregates normalized to total visualized area. Averages and SD of the mean were derived for n = 10 muscles. **, p < 0.001, two-way ANOVA with Sidak’s multiple comparisons test.
Longevity phenotypes have also been linked to food consumption [15,29]; accordingly, we explored this possibility in our flies, determining food uptake using the CAFÉ assay [25]. The results showed no reduction of food consumption in the ucp4a mutant (S2 Fig). We further tested whether reduction of protein aggregates leads to healthy muscles using climbing assays. In these assays, the ucp4a mutant exhibited better climbing ability, suggesting that ucp4a mutant flies possess healthier muscles (Fig 5). Flies with Ucp4a knockdown in muscle also exhibited better climbing ability (S3 Fig).
*Climbing assay.The number of male flies per vial that climbed 7 cm within 10 sec after tapping was counted. Ucp4aG1388 indicates Ucp4a hemizygous mutant flies (Ucp4aG1388/Y). 1st, 2nd, 3rd denote individual trials. Error bars indicate ±SEM. n = 100. **, p < 0.001, Student’s t-test.
Discussion
Here we show that mutation in the Ucp4a gene can extend lifespan without restricting feeding. Remarkably, the life-extension effect of Ucp4a mutation is specifically due to knockdown of Ucp4a in muscles; knockdown in other tissues was not effective in life-extension. We find that protein aggregates, a characteristic of muscle aging, are reduced by Ucp4a knockdown in muscles. Consistently, Ucp4a mutants and lines with Ucp4a knockdown in muscle maintain healthier muscle than control flies, as suggested by observation of enhanced locomotor activity in aged flies.
Our data show that Ucp4a hemizygous male and Ucp4a homozygous female flies still possess 20 ~ 40% the level of control Ucp4a transcripts, suggesting that they still possess some Ucp4a activity. Intriguingly, Ucp4a heterozygous females, which harbor one wild-type Ucp4a allele, exhibited longer lifespans compared to homozygous mutant females, suggesting that milder Ucp4a distress is more effective in lifespan extension. This finding is in line with previous reports that mild distress of mitochondria increases lifespan whereas severe distress shortens it [4]. In future, the effect of dose-dependency on lifespan could be tested using Ucp4a null mutants if available. Our present findings suggest that proper level of Ucp4a activity disturbance in the mitochondria might be a key determinant of lifespan.
Aspartate (Asp) is converted to asparagine (Asn) by the asparagine synthetase (ASNS) enzyme in the cytosol [12], suggesting that Ucp4a knockdown is likely to reduce cytosolic Asn. This reduction might be a signal that relates to inhibition of the mTORC1 pathway [12,30–32]. This possibility is supported by a recent finding that inhibition of glutaminolysis, which is required for asp synthesis, activates the ATF4-mediated pathway to suppress mTORC1 activity through expression of the mTORC1 negative regulators Sestrin2 and Redd1 [2,22]. Ultimately, asp reduction-mediated lifespan extension might require inhibition of the mTORC1 pathway. Further confirmatory research remains required.
Supporting information
S1 FigPolyubiquitinated protein aggregates in muscle.Confocal images of adult thorax (indirect flight muscle) sections co-stained with polyubiquitinated protein antibody (red) and phalloidin (green, sarcomere structure). 50-day-old male flies. Ucp4a denotes Ucp4a^G1388^/Y.(TIF)
S2 FigQuantification of food uptake by capillary feeding assay.Food uptake was measured for five flies (5 ~ 7 day old) over six hours in a vial. Each bar is an average from a triplicated experiment. Two trials are shown. W1118 indicates W^1118^/Y (male), W^1118^/ W^1118^ (female). G1388 indicates Ucp4a^G1388^/Y(male), Ucp4a^G1388^/Ucp4a^G1388^ (female).(TIF)
S3 FigClimbing assay.The number of flies per vial that climbed 7 cm within 10 sec after tapping was counted. Male flies of different ages. MHC/+ indicates MHC-Gal4/+ . Ucp4a RNAi/+ indicates UAS-Ucp4a RNAi #6162/+ . MHC/Ucp4a RNAi indicates MHC-Gal4/UAS-Ucp4a RNAi #6162. Error bars indicate ±SEM. n = 100. *, p < 0.05, Student’s t-test.(TIF)
S1 DataIndividual numerical values displayed in all graphs.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Liu Y, Birsoy K. Metabolic sensing and control in mitochondria. Mol Cell. 2023;83(6):877–89. doi: 10.1016/j.molcel.2023.02.016 36931256 PMC 10332353 · doi ↗ · pubmed ↗
- 2Condon KJ, Orozco JM, Adelmann CH, Spinelli JB, van der Helm PW, Roberts JM, et al. Genome-wide CRISPR screens reveal multitiered mechanisms through which m TORC 1 senses mitochondrial dysfunction. Proc Natl Acad Sci U S A. 2021;118(4):e 2022120118. doi: 10.1073/pnas.2022120118 33483422 PMC 7848693 · doi ↗ · pubmed ↗
- 3Chakrabarty RP, Chandel NS. Beyond ATP, new roles of mitochondria. Biochem (Lond). 2022;44(4):2–8. doi: 10.1042/bio_2022_119 36248614 PMC 9558425 · doi ↗ · pubmed ↗
- 4Copeland JM, Cho J, Lo T Jr, Hur JH, Bahadorani S, Arabyan T, et al. Extension of Drosophila life span by RN Ai of the mitochondrial respiratory chain. Curr Biol. 2009;19(19):1591–8. doi: 10.1016/j.cub.2009.08.016 19747824 · doi ↗ · pubmed ↗
- 5Dell’agnello C, Leo S, Agostino A, Szabadkai G, Tiveron C, Zulian A, et al. Increased longevity and refractoriness to Ca(2+)-dependent neurodegeneration in Surf 1 knockout mice. Hum Mol Genet. 2007;16(4):431–44. doi: 10.1093/hmg/ddl 477 17210671 · doi ↗ · pubmed ↗
- 6Parkhitko AA, Filine E, Mohr SE, Moskalev A, Perrimon N. Targeting metabolic pathways for extension of lifespan and healthspan across multiple species. Ageing Res Rev. 2020;64:101188. doi: 10.1016/j.arr.2020.101188 33031925 PMC 9038119 · doi ↗ · pubmed ↗
- 7Feng J, Bussière F, Hekimi S. Mitochondrial electron transport is a key determinant of life span in Caenorhabditis elegans. Dev Cell. 2001;1(5):633–44. doi: 10.1016/s 1534-5807(01)00071-5 11709184 · doi ↗ · pubmed ↗
- 8Fessler E, Eckl E-M, Schmitt S, Mancilla IA, Meyer-Bender MF, Hanf M, et al. A pathway coordinated by DELE 1 relays mitochondrial stress to the cytosol. Nature. 2020;579(7799):433–7. doi: 10.1038/s 41586-020-2076-4 32132706 PMC 7116715 · doi ↗ · pubmed ↗