Barley roots require HvGLU3 for cellulose biosynthesis
Blanca Jazmin Reyes-Hernández

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
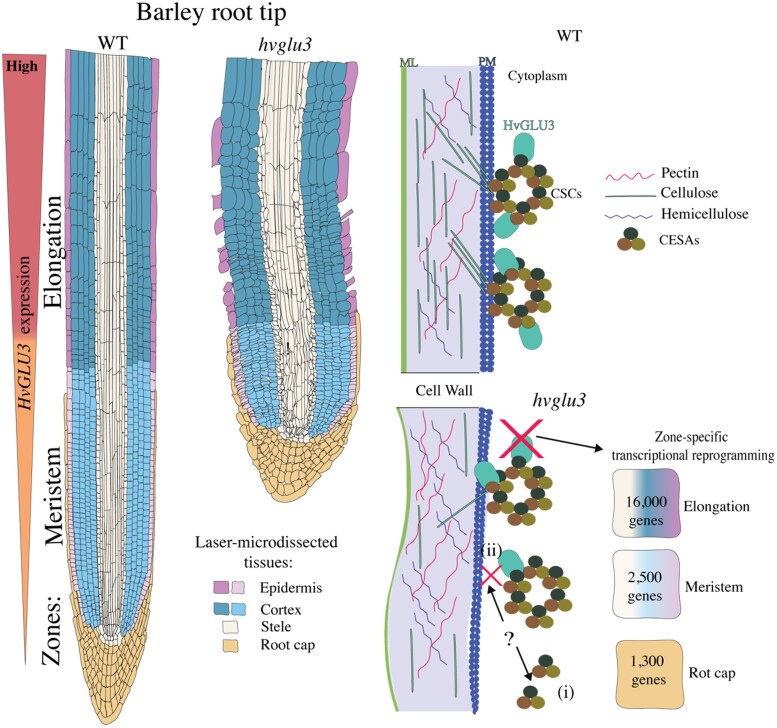 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant nutrient uptake and metabolism
Plant cells are enclosed by the cell wall, a complex polysaccharide structure that strengthens the plant body and supports plant development, cell communication, and immunity. The cell wall is composed of different layers. The outermost layer, the middle lamella, is shared between adjacent cells and holds them together. Between the middle lamella and the plasma membrane lies the primary cell wall. In some specialized cells that require greater mechanical strength, a secondary cell wall is developed inside the primary wall. Primary and secondary walls consist mainly of cellulose and hemicelluloses; the primary wall also contains pectin while the secondary is enriched in lignin (Cosgrove 2005).
Cellulose is synthesized at the plasma membrane by cellulose synthase complexes (CSCs). The resulting fibrils contain very ordered tough crystalline regions and looser amorphous parts, giving the cell wall its strength and some flexibility (Cosgrove 2005).
In Arabidopsis, the membrane-anchored endo-1,4-ß-D-glucanase KORRIGAN1(KOR1) is a key component of CSCs (Mansoori et al. 2014). Endo-1,4-ß-D-glucanases are enzymes that break down long chains of polysaccharides such as cellulose that are joined by a 1,4-ß-D glucan bond (Perrot et al. 2022). KOR1 is suggested to trim nascent 1,4-β-D glucan chains rather than break down mature tough crystalline cellulose (Mølhøj et al. 2001). Its correct delivery from the endoplasmic reticulum through the Golgi to the membrane is tightly regulated. When KOR1 is absent or mislocalized, cellulose fibers are poorly assembled, and the new nascent cell walls (cell plate) fails to close completely during cytokinesis, an effect that is more severe under salt stress (Nagashima et al. 2020). Despite the importance of KOR1 in cellulose biosynthesis and cell division, the mechanism by which endo-1,4-ß-D-glucanases regulate these processes remains unexplored.
In a recent study published in Plant Physiology (Guo et al. 2025), a barley mutant with short roots caused by repressed cell division and elongation was identified. This mutant, which originated from chemical mutagenesis, displayed twisted roots with a rough surface resembling roots with problems in cellulose biosynthesis. Gene mapping showed that the affected gene was HvGLU3, which encodes for a putative ß-endoglucanase. This mutant was designated as hvglu3-1 and presents a single-nucleotide substitution causing an amino acid change from serine to asparagine residue at position 178 of the protein.
A second allele was identified with a premature stop codon and a more severe phenotype, confirming that the mutant phenotype was caused by the loss of HvGLU3 function. Although the aerial parts of the hvglu3-1 mutant look normal, they are smaller than the wild type. Cross sections in the root of the mutant indicated that, despite an overall normal pattern, (i) the epidermis was thicker in the meristem region and (ii) the cells of this tissue were lost in the elongation zone above. Reverse transcription quantitative PCR experiments demonstrated that HvGLU3 expression was normally high in the elongation and meristematic zones, while it was lower in shoot and leaf, supporting the idea that its role could be relevant in the root (Fig.).
Phylogenetic and sequence analyses revealed that HvGLU3 homologs are present throughout the plant kingdom and that the gene is highly conserved within the barley germplasm*. HVGLU3* is ortholog of the rice gene OsGLU3, which is involved in cellulose biosynthesis (Zhang et al. 2012). Rice OsGLU3 mutants display opposite phenotypes: some accumulate cellulose; others lose it (Inukai et al. 2012). Consistent with the cellulose-deficient root phenotypes observed in osglu3-1, osglu3-2 (Zhang et al. 2012), and their Arabidopsis homolog kor1 (Takahashi et al. 2009), hvglu3-1 mutant also exhibits reduced cellulose content.
To investigated HvGLU3 function in barley, the authors treated seedlings with isoxaben, a cellulose biosynthesis inhibitor. Wild type roots stopped elongating under treatment, while hvglu3-1 roots hardly changed, meaning that hvglu3-1 already has a cell wall defect. Biochemical assays confirmed a 60% drop in cellulose content in hvglu3-1. At the same time, changes in sugar composition shifted, indicating an increase in arabinoxylan hemicellulose and pectin levels, likely a compensatory mechanism to reinforce the weakened cell wall. In addition, suberin levels were reduced in hvglu3-1, pointing to a compromised protective barrier. Histologic analysis supported these biochemical results, altogether indicating that HvGLU3 is required for proper normal cellulose deposition and for maintaining the correct balance of structural and protective components in the barley root cell wall.
To gain insight into the function of HvGLU3, the authors transiently expressed it in tobacco leaves using RFP-HvGLU3 with mVenus-tagged cellulose synthases (CESAs), revealing colocalization at the plasma membrane. These interactions were confirmed by a bimolecular fluorescence complementation assay, suggesting that HvGLU3 interacts with CESAs and could be part of the cellulose biosynthesis machinery in barley.
To explore how defective cellulose biosynthesis in hvglu3-1 affects the plant at a molecular level, the authors performed RNA sequencing on specific root tissues using laser capture microdissection. They isolated the stele, cortex, epidermis, and root cap, separating cells from the meristem and elongation zones. The elongation zone exhibited the strongest differential transcriptional response when compared with wild type roots, with >16,000 genes differentially expressed, as opposed to about 2,500 in the meristem and 1,300 in the root cap. In all tissues, upregulated genes were 2 to 3 times more abundant than downregulated ones. Genes involved in cellulose biosynthesis were mostly downregulated, while lignin-related genes were upregulated in mutant roots. These results indicate that the defect in cellulose biosynthesis in the mutant hvglu3-1 leads to a substantial transcriptomic reprogramming across various root tissues.
To understand broader patterns of gene expression, the authors used coexpression network analysis and found 18 gene modules linked to specific tissue types. Each module included hub genes highly connected to key biological functions, such as those related to the elongation zone, meristem cortex, and root cap. A separate analysis focused on genotype differences indicated 9 modules associated with hvglu3-1 or wild type. Some of these overlapped with tissue-specific modules, showing that gene expression changes in the mutant depend on tissue type and genotype.
This study highlights HvGLU3 as a key endoglucanase needed for cellulose production and root development in barley. Its loss not only reconfigures the transcriptome but also leads to epidermal damage. As yet, it is not known whether HvGLU3 works by cutting glucan chains enzymatically or by helping the delivery or function of CSCs. The authors also raise new questions—for example, what signals cause the increase in lignin and drop in suberin in mutant roots? The hub genes found in the elongation zone epidermis may play a role in these shifts. Overall, these findings link cell wall building blocks to tissue development and open new possible ways to tune plant resilience.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cosgrove DJ . Growth of the plant cell wall. Nat Rev Mol Cell Biol. 2005:6(11):850–861. 10.1038/nrm 174616261190 · doi ↗ · pubmed ↗
- 2Guo L, Rosignoli S, Rasmussen MW, Suresh K, Sangiorgi G, Camerlengo F, Zeisler-Diehl VV, Schreiber L, Dockte C, Pauly M, et al Hv GLU 3 encodes a putative endo-1,4-β-D-glucanase required for cellulose biosynthesis in barley root. Plant Physiol. 2025:198(3):311–326. 10.1093/plphys/kiaf 311PMC 1231130040674700 · doi ↗ · pubmed ↗
- 3Inukai Y, Sakamoto T, Morinaka Y, Miwa M, Kojima M, Tanimoto E, Yamamoto H, Sato K, Katayama Y, Matsuoka M, et al ROOT GROWTH INHIBITING, a rice endo-1,4-β-d-glucanase, regulates cell wall loosening and is essential for root elongation. J Plant Growth Regul. 2012:31(3):373–381. 10.1007/s 00344-011-9247-3 · doi ↗
- 4Mansoori N, Timmers J, Desprez T, Alvim-Kamei CL, Dees DCT, Vincken J-P, Visser RGF, Höfte H, Vernhettes S, Trindade LM. KORRIGAN 1 interacts specifically with integral components of the cellulose synthase machinery. P Lo S One. 2014:9(11):e 112387. 10.1371/journal.pone.011238725383767 PMC 4226561 · doi ↗ · pubmed ↗
- 5Mølhøj M, Ulvskov P, Dal Degan F. Characterization of a functional soluble form of a Brassica napus membrane-anchored endo-1,4-beta-glucanase heterologously expressed in Pichia pastoris. Plant Physiol. 2001:127(2):674–684. 10.1104/pp.01026911598241 PMC 125102 · doi ↗ · pubmed ↗
- 6Nagashima Y, Ma Z, Liu X, Qian X, Zhang X, von Schaewen A, Koiwa H. Multiple quality control mechanisms in the ER and TGN determine subcellular dynamics and salt-stress tolerance function of KORRIGAN 1. Plant Cell. 2020:32(2):470–485. 10.1105/tpc.19.0071431852774 PMC 7008481 · doi ↗ · pubmed ↗
- 7Perrot T, Pauly M, Ramírez V. Emerging roles of β-glucanases in plant development and adaptative responses. Plants. 2022:11(9):1119. 10.3390/plants 1109111935567119 PMC 9099982 · doi ↗ · pubmed ↗
- 8Takahashi J, Rudsander UJ, Hedenström M, Banasiak A, Harholt J, Amelot N, Immerzeel P, Ryden P, Endo S, Ibatullin FM, et al KORRIGAN 1 and its aspen homolog Ptt Cel 9A 1 decrease cellulose crystallinity in Arabidopsis stems. Plant Cell Physiol. 2009:50(6):1099–1115. 10.1093/pcp/pcp 06219398462 · doi ↗ · pubmed ↗