Draft genomes of five bacteria isolated from Carcharodon carcharias (white shark) and Carcharhinus brachyurus (bronze whaler shark)
Emma N. Kerr, Ryan D. Hesse, Jessica A. P. Carlson-Jones, Bhavya Nalagampalli Papudeshi, Paul A. Butcher, Michael P. Doane, Elizabeth A. Dinsdale

TL;DR
This paper presents draft genomes of five bacteria found on the skin and cloaca of two shark species.
Contribution
The study provides new genomic data for bacteria isolated from white and bronze whaler sharks.
Findings
Five bacterial species were isolated from the skin and cloaca of two shark species.
Draft genomes of these bacteria were sequenced and reported.
Abstract
Here, we isolated five bacterial species from the skin and cloaca of Carcharodon carcharias (white shark) and Carcharhinus brachyurus (bronze whaler or copper shark).
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
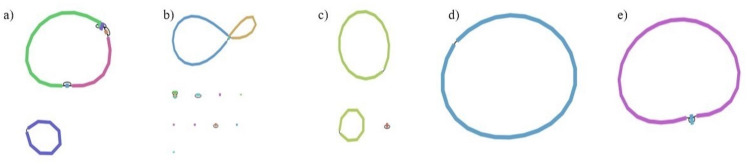 Fig 1
Fig 1| Species | Host | Genome length (bp) | Number of reads | Sequence N50 | Contigs | Assembly N50 | GC content (%) | Average coverage | Annotated genes | SRA | GenBank |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 4,962,167 | 294,978 | 7.957 | 3 | 1,510,937 | 43.92 | 23.3 | 4,906 |
|
| ||
| 4,750,424 | 735,427 | 8.892 | 11 | 2,972,306 | 38.42 | 52.4 | 4,982 |
|
| ||
| 4,983,971 | 1,060,052 | 6.743 | 3 | 3,372,530 | 44.07 | 98.3 | 4,709 |
|
| ||
| 4,871,077 | 342,618 | 8.310 | 1 | 4,871,077 | 40.83 | 29 | 4,537 |
|
| ||
| 5,949,190 | 1,352,592 | 6.366 | 1 | 5,949,190 | 43.43 | 84 | 5,784 |
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsIchthyology and Marine Biology · Genomics and Phylogenetic Studies · Aquaculture disease management and microbiota
ANNOUNCEMENT
Shark microbiology originated in response to shark bites (1). However, microbes are integral in shark health and ecology (2, 3). Elasmobranchs support biodiversity in marine ecosystems but are globally threated by overfishing and biomagnification of heavy metals (4–6). Biomagnification is reflected in shark microbial communities where heavy metal resistance genes are enriched (7–10). We sequenced the genomes of bacterial isolates from Carcharodon carcharias (white shark) and Carcharhinus brachyurus (bronze whaler shark).
Sharks were caught on SMART drumlines at Port Macquarie, New South Wales (NSW DPI scientific [Ref. P01/0059(A)], Marine Parks [Ref. P16/0145-1.1], and Animal Care and Ethics [ACEC Ref. 07/08]) (11, 12). Swabs were collected from the skin and cloaca, placed in a sterile seawater-filled microfuge tube and transported on ice. The bacterial slurry (50 µL) was plated on Thiosulfate–citrate–bile salts–sucrose agar (Bacto Laboratories, Australia) and incubated at 18°C until colonies formed. Colonies were isolated and picked for DNA extraction using the Nucleospin Tissue Kit (Macherey-Nagel, Germany). DNA concentrations were quantified with a Qubit 4 fluorometer (Invitrogen, USA). Libraries were prepared with raw DNA without size selection using the Nanopore Rapid Barcoding Kit and sequenced on the MinION (flow cell R9.4.1) (Oxford Nanopore, United Kingdom). Default parameters were used for all bioinformatics tools. Guppy basecalling (V 6.5.7) converted raw signal to base pairs (Oxford Nanopore Technology). Hybracter (V 0.10.1) quality-controlled sequences and assembled contigs (13). Average coverage is the mean coverage of all contigs in a genome. Taxonomic classification was determined using a BLASTN search of the entire genome sequence (14) and Bakta (V 1.10) for functional annotation (15). Bandage (v0.8.1) was used to visualize genomes (16).
Genomes were complete and contained 4,750,424 to 5,949,190 base pairs in 1–11 contigs (Fig. 1, Table 1). Antimicrobial and heavy metal resistance genes were present in all genomes. Cloaca isolates have class A beta-lactamase, heavy metal efflux pump, trimethylamine-N-oxide (TMAO) reductase, and urease genes which are lacking from skin isolates. TMAO and urea are used to maintain osmotic balance by Elasmobranchs (17). Presence of these genes may indicate compounds utilized by shark microbes. Cation Diffusion Facilitator (CDF) family efflux transporter DmeF gene was present in skin isolates and may provide resistance to heavy metals (18).
Visualization of genome assemblies. (a) Vibrio toranzoniae strain ENK1. (b) Aliivibrio fischeri ES114 strain ENK5. (c) Vibrio atlanticus strain ENK4. (d) Shewanella sp. WPAGA9 strain ENK2. (e) Shewanella woodyi strain ENK6.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Buck JD, Spotte S, Gadbaw JJ. 1984. Bacteriology of the teeth from a great white shark: potential medical implications for shark bite victims. J Clin Microbiol 20:849–851. doi:10.1128/jcm.20.5.849-851.19846511869 PMC 271456 · doi ↗ · pubmed ↗
- 2Doane MP, Morris MM, Papudeshi B, Allen L, Pande D, Haggerty JM, Johri S, Turnlund AC, Peterson M, Kacev D, et al.. 2020. The skin microbiome of Elasmobranchs follows phylosymbiosis, but in teleost fishes, the microbiomes converge. Microbiome 8:93. doi:10.1186/s 40168-020-00840-x 32534596 PMC 7293782 · doi ↗ · pubmed ↗
- 3Perry CT, Pratte ZA, Clavere-Graciette A, Ritchie KB, Hueter RE, Newton AL, Fischer GC, Dinsdale EA, Doane MP, Wilkinson KA, Bassos-Hull K, Lyons K, Dove ADM, Hoopes LA, Stewart FJ. 2021. Elasmobranch microbiomes: emerging patterns and implications for host health and ecology. Anim Microbiome 3:61. doi:10.1186/s 42523-021-00121-434526135 PMC 8444439 · doi ↗ · pubmed ↗
- 4Henderson CJ, Gilby BL, Turschwell MP, Goodridge Gaines LA, Mosman JD, Schlacher TA, Borland HP, Olds AD. 2024. Long term declines in the functional diversity of sharks in the coastal oceans of eastern Australia. Commun Biol 7:611. doi:10.1038/s 42003-024-06308-038773323 PMC 11109089 · doi ↗ · pubmed ↗
- 5Wosnick N, Niella Y, Hammerschlag N, Chaves AP, Hauser-Davis RA, da Rocha RCC, Jorge MB, de Oliveira RWS, Nunes JLS. 2021. Negative metal bioaccumulation impacts on systemic shark health and homeostatic balance. Mar Pollut Bull 168:112398. doi:10.1016/j.marpolbul.2021.11239833906010 · doi ↗ · pubmed ↗
- 6Weijs L, Briels N, Adams DH, Lepoint G, Das K, Blust R, Covaci A. 2015. Bioaccumulation of organohalogenated compounds in sharks and rays from the southeastern USA. Environ Res 137:199–207. doi:10.1016/j.envres.2014.12.02225569844 · doi ↗ · pubmed ↗
- 7Doane MP, Haggerty JM, Kacev D, Papudeshi B, Dinsdale EA. 2017. The skin microbiome of the common thresher shark (Alopias vulpinus) has low taxonomic and gene function β-diversity. Environ Microbiol Rep 9:357–373. doi:10.1111/1758-2229.1253728418094 · doi ↗ · pubmed ↗
- 8Doane MP, Johnson CJ, Johri S, Kerr EN, Morris MM, Desantiago R, Turnlund AC, Goodman A, Mora M, Lima LFO, Nosal AP, Dinsdale EA. 2023. The epidermal microbiome within an aggregation of leopard sharks (Triakis semifasciata) has taxonomic flexibility with gene functional stability across three time-points. Microb Ecol 85:747–764. doi:10.1007/s 00248-022-01969-y 35129649 PMC 9957878 · doi ↗ · pubmed ↗