FvHsfB1a Gene Improves Thermotolerance in Transgenic Arabidopsis
Qian Cao, Tingting Mao, Kebang Yang, Hanxiu Xie, Shan Li, Hao Xue

TL;DR
This study shows that the FvHsfB1a gene from woodland strawberry improves heat tolerance in transgenic Arabidopsis plants.
Contribution
The study identifies FvHsfB1a as a positive regulator of heat stress and reveals its regulation by FvWRKY75.
Findings
Ectopic expression of FvHsfB1a in Arabidopsis improves thermotolerance with higher germination and survival rates.
FvWRKY75 activates the FvHsfB1a promoter through the W-box element, leading to heat-tolerant phenotypes in transgenic plants.
Abstract
Heat stress transcription factor (Hsf) families play important roles in abiotic stress responses. However, previous studies reported that HsfBs genes may play diverse roles in response to heat stress. Here, we conducted functional analysis on a woodland strawberry Class B Hsf gene, FvHsfB1a, to improve thermotolerance. The structure of FvHsfB1a contains a typical Hsf domain for DNA binding at the N-terminus, and FvHsfB1a belongs to the B1 family of Hsfs. The FvHsfB1a protein was localized in the nucleus. The FvHsfB1a gene was expressed in various strawberry tissues and highly induced by heat treatment. Under heat stress conditions, ectopic expression of FvHsfB1a in Arabidopsis improves thermotolerance, with higher germination and survival rates, a longer primary root length, higher proline and chlorophyll contents, lower malonaldehyde (MDA) and O2− contents, better enzyme activities,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
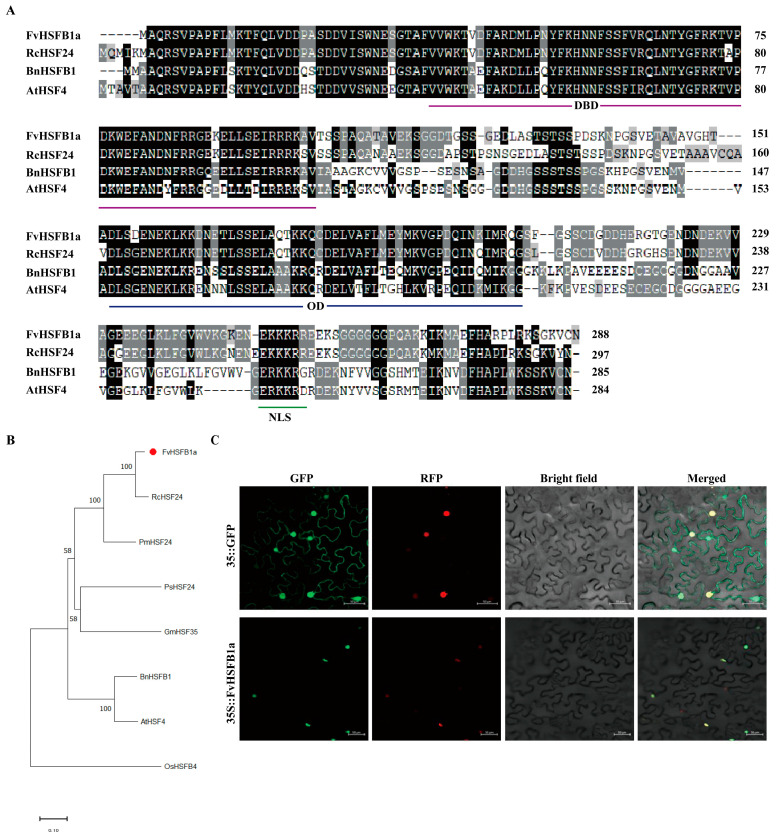 Figure 1
Figure 1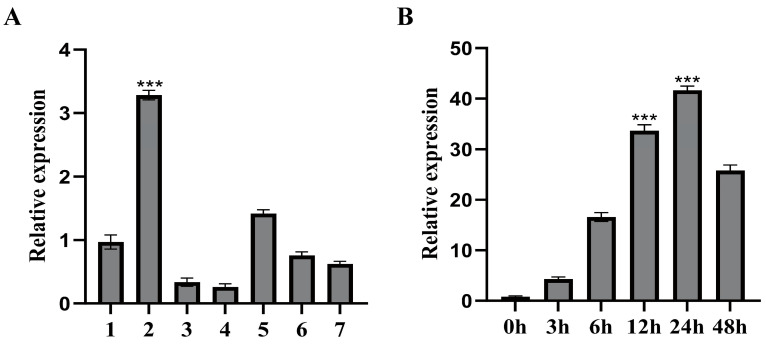 Figure 2
Figure 2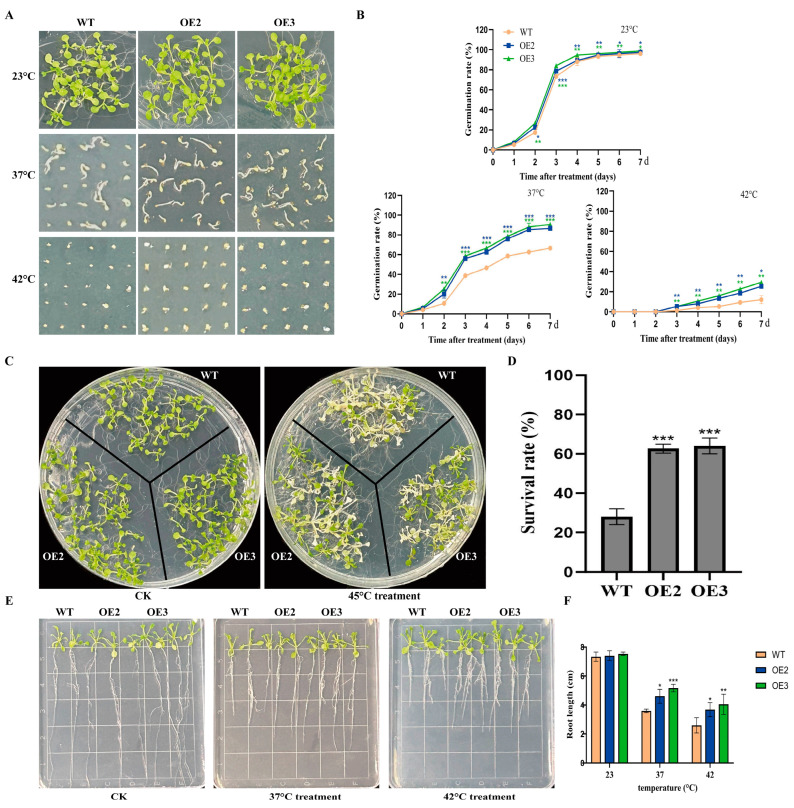 Figure 3
Figure 3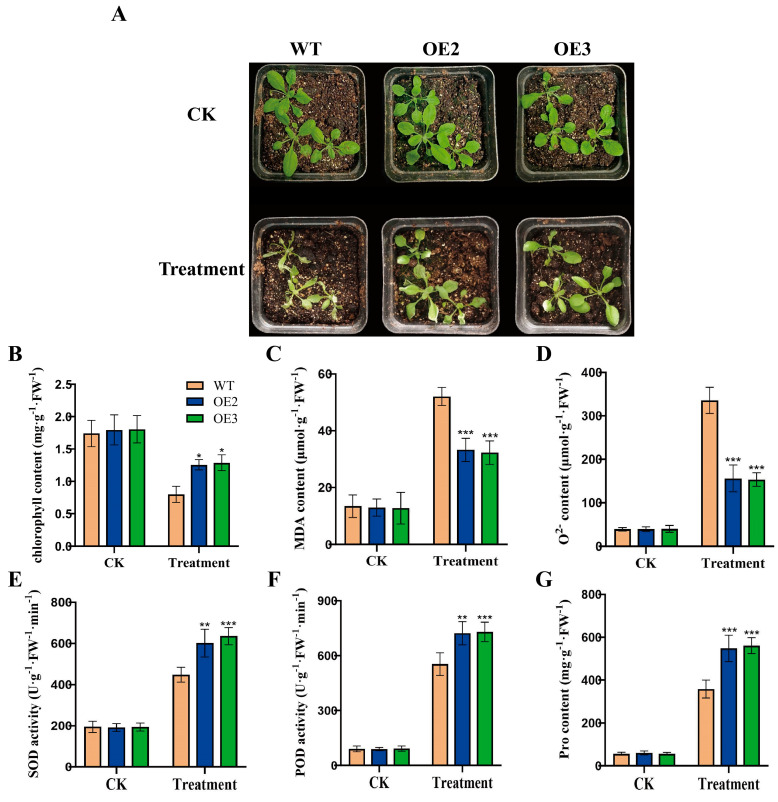 Figure 4
Figure 4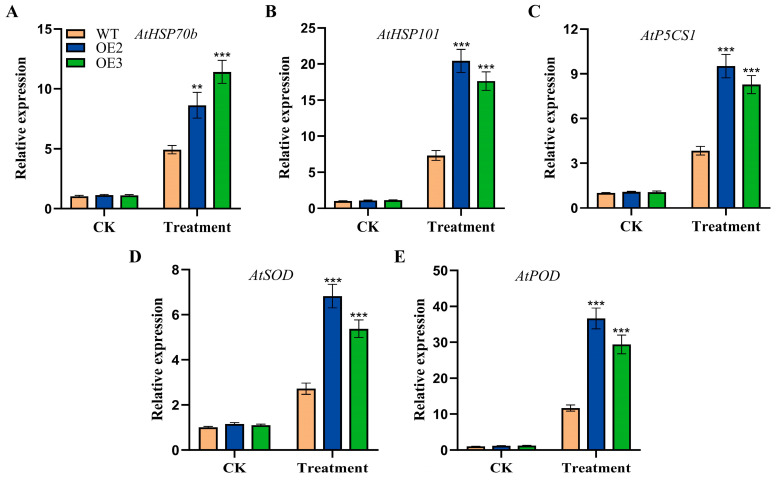 Figure 5
Figure 5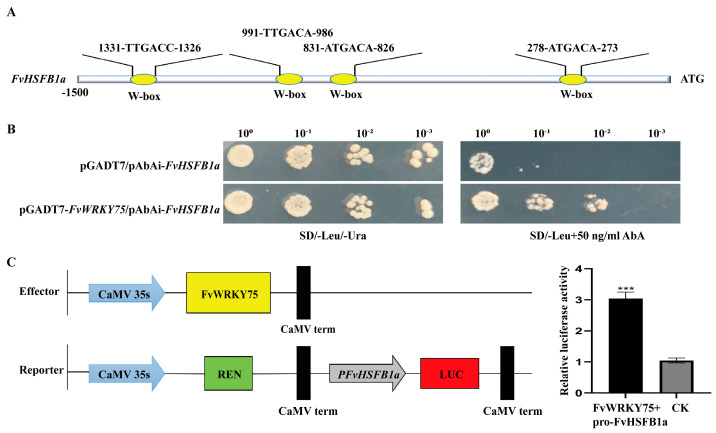 Figure 6
Figure 6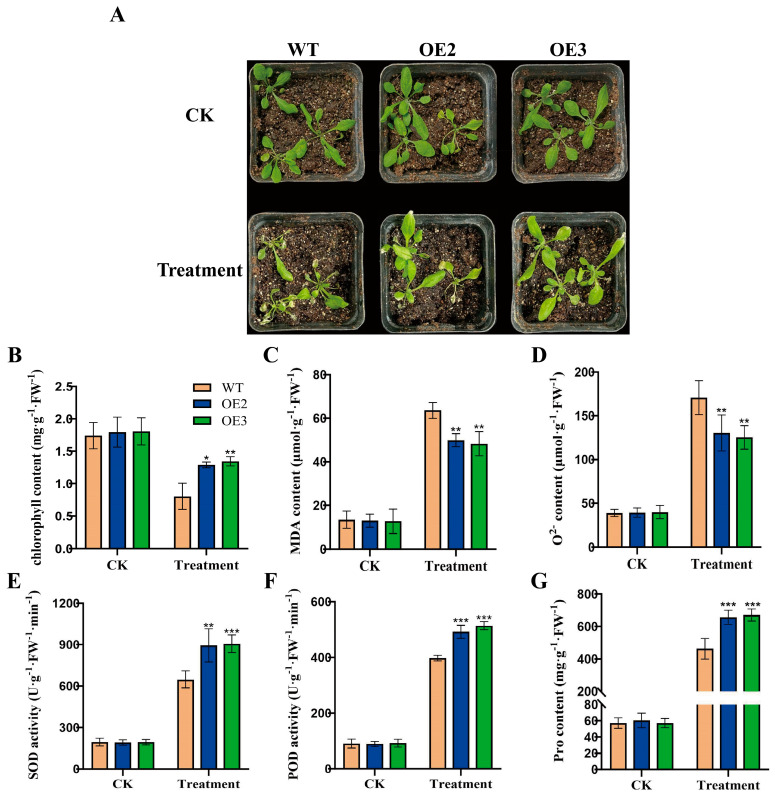 Figure 7
Figure 7- —Natural Science Foundation of Anhui Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Stress Responses and Tolerance · Plant Molecular Biology Research · Photosynthetic Processes and Mechanisms
1. Introduction
The growth and development of plants require suitable environmental conditions, generally including temperature, light, water, and soil nutrients [1]. Adverse environmental factors, especially heat stress, cause several morphological and physiological changes in plants. This includes protein denaturation, plasma membrane disruption, and reactive oxygen species (ROS) accumulation, and can lead to plant death in severe cases [2,3,4,5]. Plants have a series of complex physiological mechanisms and molecular regulatory networks to minimize the damage caused by the external environmental stresses [6,7].
The heat shock transcription factors (Hsfs) family is generally known to play a central role in regulating abiotic stress response [8,9,10]. Based on the conserved Hsf DNA-binding domain (DBD) and oligomerization domain (OD) regions, Hsfs can be divided into three classes in higher plants, namely HsfA, HsfB, and HsfC [11,12]. In a previous study, HsfAs were found to be master regulators of thermotolerance by regulating the expression of downstream genes in plants [13,14,15,16]. In contrast to HsfAs, HsfBs have no transcriptional activity, because they lack a C-terminal activator domain (CTAD) [17]. Previous studies reported that HsfBs are induced by heat stress and involved in the regulation of thermotolerance in many plant species [18,19]. For example, AtHsfB1 and AtHsfB2b act as suppressors of the expression of Hsfs and several heat shock protein genes but are necessary for acquiring thermotolerance [20]; PpHsf5 in peaches and HsfB1 in grapes were reported to play a positive role in thermotolerance [21,22]. In addition, some HsfBs genes, such as OsHsfB2b from rice, CarHsfB2 from chickpeas, GmHsfB2b from soybeans, and ZmHsf08 (Class B gene) from maize, play important roles in drought and salt tolerance responses [23,24,25,26]. For HsfCs members, it is demonstrated that HsfC2a from wheat and HsfC1b from tall fescue regulate the heat-responsive genes to increase heat tolerance, and VaHsfC1 from grapes is involved in multiple abiotic stresses [27,28,29,30].
The strawberry is cultivated widely in the world and is threatened by a range of adverse environmental factors. With global warming, the increasing number of extreme temperature events has severely affected the growth, development, productivity, and quality of strawberry, posing a considerable challenge to modern agricultural farming [31,32]. Seventeen Hsf genes have been identified in the diploid woodland strawberry (Fragaria vesca), and different FvHsfs have distinct responses to various biotic and abiotic stresses at the transcriptional level [33]. However, the molecular mechanisms of the FvHsfs genes involved in abiotic stress remain unclear. Herein, we cloned the FvHsfB1a gene from ‘Ruegen’ strawberry leaves and found that FvHsfB1a was induced by heat stress treatment. We then explored its function in heat stress response. The overexpression of the FvHsfB1a gene enhances heat stress resistance. Compared with WT plants, heat-responsive and stress-related genes in FvHsfB1a-OE lines were rapidly and strongly induced by heat stress treatment. Furthermore, the FvHsfB1a gene was directly regulated by the FvWRKY75 TF. The overexpression of the FvWRKY75 gene in Arabidopsis also improved heat tolerance. Overall, we have provided evidence that FvHsfB1a may play an important role in improving strawberry thermotolerance.
2. Results
2.1. Isolation and Characterization of FvHsfB1a
The FvHsfB1a gene was isolated from ‘Ruegen’ leaves and the CDS sequence of FvHsfB1a was 873 bp, encoding a protein containing 290 amino acids (Figure 1A). The predicted molecular weight and isoelectric point of FvHsfB1a protein were predicted to be 32.01 kDa and 5.96 M. The structure of FvHsfB1a contained a typical HSF domain for DNA binding at the N-terminus (Figure 1A). Phylogenetic analysis showed that the sequence of FvHsfB1a was the closest homolog to RcHSF24 (class B1) (Figure 1B).
The subcellular localization of FvHsfB1a protein was detected via green fluorescent protein (GFP) and red fluorescent protein (RFP) as a nuclear location marker. As shown in Figure 1C, the recombinant pCAMBIA1302-FvHsfB1a-GFP was localized only in the nucleus.
2.2. Expression Profiles of FvHsfB1a in Diploid Strawberry
The tissue-specific expression patterns of FvHsfB1a were measured in the diploid strawberry ‘Ruegen’. The FvHsfB1a gene was differentially expressed in different tissues, with the lowest expression in leaves and the highest expression in the stem (Figure 2A). Then, we investigated the expression profiles of the FvHsfB1a gene by using mature strawberry leaves under abiotic stress conditions. Under heat stress treatment, FvHsfB1a gene expression was significantly induced and reached a peak with a 43.7-fold increase after 24 h of treatment (Figure 2B).
2.3. FvHsfB1a Improves Thermotolerance in Arabidopsis
To examine the function of FvHsfB1a under heat stress, we generated FvHsfB1a transgenic Arabidopsis plants, and two FvHsfB1a-OE T3 homozygous lines were selected by using PCR and qRT-PCR for future tests (Figure S1). As shown in Figure 3, no significant differences in seed germination and seedling growth were observed between the WT and OE lines under normal conditions. After treatment with heat stress for 7 d, the WT seeds exhibited the lowest germination rates, especially under the 42 °C treatment (Figure 3A,B). In contrast, the FvHsfB1a-OE seeds had higher germination rates in 37 °C and 42 °C treatments for 7 d compared to WT (Figure 3A,B). Meanwhile, the performance of FvHsfB1a-OE and WT Arabidopsis seedlings was evaluated after 45 °C treatment for 3 h. Compared with WT, there were better performances, higher survival rates, and longer root lengths in FvHsfB1a-OE seedlings observed (Figure 3C,D). Additionally, the heat tolerance of WT and FvHsfB1a-OE mature plants was also detected. Compared with WT, FvHsfB1a-OE lines showed a lower degree of leaf wilting, higher chlorophyll and proline contents, lower MDA and O_2_^−^ contents, and better superoxide (SOD) and peroxidase (POD) activities after heat stress treatment (Figure 4A–G). These results suggest that the FvHsfB1a gene could improve thermotolerance in Arabidopsis.
2.4. Overexpression of FvHsfB1a Regulates the Expression of Stress-Related Genes
To further investigate the regulation of FvHsfB1a in response to heat stress, the expression profiles of several heat-responsive and stress-related genes were detected by using qRT-PCR in the WT and FvHsfB1a-OE lines. After treatment with heat stress, the expression levels of heat-responsive genes (AtHSP70b and AtHSP101) and stress-related genes (AtP5CS1, AtSOD, and AtPOD) were significantly upregulated in FvHsfB1a-OE lines as compared with the WT (Figure 5).
2.5. FvWRKY75 Can Active the Promoter of FvHsfB1a Gene
Bioinformatics analysis showed that the promoter of the FvHsfB1a gene contains several WRKY binding elements (Figure 6A). Based on previous findings, FvWRKY70 and FvWRKY75 were selected for a yeast one-hybrid assay to determine their relationship with FvHsfB1a. As shown in Figure 6B,C, the FvWRKY75 directly bound to the FvHsfB1a promoter and influenced its expression in the yeast one-hybrid and LUC assays.
2.6. FvWRKY75 Enhances Thermotolerance in Arabidopsis
Previously, we cloned the FvWRKY75 gene and generated two FvWRKY75-OE transgenic Arabidopsis plants [34]. In this study, we examined the role of the FvWRKY75 gene in the response to high-temperature stress. During heat treatment, the FvWRKY75-OE transgenic plants exhibited greater insensitivity to heat stress with a better phenotype compared to the WT (Figure 7A). Compared with the WT, lower MDA and O_2_^−^ contents, higher chlorophyll and proline contents, and better peroxidase activity were observed in the FvWRKY75-OE plants (Figure 7B–G), which suggested that FvWRKY75 is also a positive regulator of heat tolerance.
3. Discussion
Heat stress severely affects the growth, development, production, and yields of most agricultural crops [35,36,37,38]. The Hsf family has been well-identified in many plant species and functions as a key regulatory component involved in various stress responses [10,12,39]. In strawberries, 17 Hsfs were identified based on the wild diploid woodland strawberry (Fragaria vesca) [33]. Here, we cloned an Hsf TF gene, FvHsfB1a, from ‘Ruegen’ leaves, and explored its characterization. The amino acid sequence of FvHsfB1a showed all the characteristics of Class B HSFs, and the sequence of FvHsfB1a showed high homology with that of RcHsf24, which belongs to Class B1 Hsfs (Figure 1A,B). The FvHsfB1a protein was localized only in the nucleus (Figure 1C,D). Some Hsf B TF genes, including OsHSF2b, CarHSFB2, and ZmHsf08, were upregulated under abiotic stresses and proved to play a role in heat stress response [23,24,26]. In the current study, we found that the FvHsfB1a gene was induced by heat stress (Figure 2), which suggested that the FvHsfB1a gene may be involved in the heat stress response in strawberry.
Previous studies reported that HsfBs genes may play diverse roles in abiotic stress responses. For example, the overexpression of VdHSFB1 and VvHSFB1 in grapes improved heat tolerance, and the overexpression of CarHSFB2 and GmHsfB2b improved resistance to drought and salt stress in transgenic plants [22,23,25]. However, OsHsfB2b and ZmHsf08 (a member of HSFB1) negatively regulated drought and salt tolerance in rice and maize [24,26]. To better understand the function of the FvHsfB1a gene in strawberry, two FvHsfB1a-OE Arabidopsis lines were generated and used for future tests (Figure S1). After heat stress treatment, FvHsfB1a-OE lines showed increased insensitivity to high temperature, longer root length, higher germination rates, and higher survival rates than the WT (Figure 3E,F), which suggested that FvHsfB1a may regulate the growth of roots in response to heat stress. Under stress conditions, chlorophyll degradation and membrane lipid peroxidation occur in plants, resulting in a decrease in chlorophyll content and an increase in ROS (O^2−^ and H_2_O_2_) and MDA content. Therefore, chlorophyll, MDA, O^2−^, and H_2_O_2_ contents are commonly used to assess the severity of stress damage in plants [40,41,42]. In addition, antioxidant enzymes, such as POD, CAT, and APX, play important roles in scavenging oxidative damage caused by ROS accumulation [43,44]. In this study, it was noticed that FvHsfB1a-OE lines had a lower MDA content and higher chlorophyll and proline contents than WT after stress treatment; meanwhile, a lower O_2_^−^ content and higher SOD and POD activities were presented for the former rather than the latter (Figure 4), which suggested that FvHsfB1a could attenuate heat stress damage to plants by modulating ROS scavenging and increasing antioxidant enzyme activity.
Heat shock proteins (HSPs) were known as the target genes of HSFs and play important roles in heat stress regulation [45,46]. HSP70 and HSP101 are induced by heat stress and confer improved heat tolerance in Arabidopsis [47,48]. In the current study, AtHSP70b and AtHSP101 were significantly induced in FvHsfB1a-OE lines compared to in WT plants under heat stress (Figure 5A,B). This suggests that FvHsfB1a could influence the expression of these HSP genes to improve thermotolerance in transgenic plants. The P5CS gene participated in the synthesis of proline, which contributes to the positive regulation of plant dehydration and osmotic stress [49]. The upregulation of the AtP5CS1 gene in the FvHsfB1a-OE lines under heat treatment may contribute to the accumulation of proline, which can reduce the injury caused by stress (Figure 5C). Many studies reported the connection between antioxidant enzyme genes and Hsfs in scavenging ROS [50,51]. In the current study, the stress-related genes (AtSOD and AtPOD) showed higher expression levels in transgenic plants under heat stress conditions (Figure 5D,E). All of the results suggest that the overexpression of the FvHsfB1a gene improves thermotolerance by upregulating the expression of heat-responsive genes (AtHSP70b and AtHSP101) and stress-related genes (AtP5CS1, AtSOD, and AtPOD).
In this study, the Y1 H and LUC assays showed that the FvWRKY75 transcription factor directly binds to the promoter region of the FvHsfB1a gene (Figure 6). It has been reported that the WRKY TFs play important roles in abiotic stress responses, especially WRKY75 [52,53,54]. For example, the overexpression of PtrWRKY75 in poplar and AhWRKY75 in peanuts improves heat, drought, and salt tolerance; the overexpression of MdWRKY75 in apple regulates the expression of Hsf genes to enhance thermotolerance [55,56,57]. In a previous study, we cloned the FvWRKY75 gene and found that the FvWRKY75 gene plays an active role in response to salt stress [34]. Herein, we explored the role of the FvWRKY75 gene in the response to high-temperature stress. It was noticed that the FvWRKY75-OE transgenic Arabidopsis plants exhibited a better phenotype compared to the WT under heat treatment, and lower MDA and O_2_^−^ contents, higher chlorophyll and proline contents, and better SOD and POD activities were observed in the former than in the latter (Figure 7). This suggests that FvWRKY75 is also a positive regulator of thermotolerance in strawberry.
In conclusion, we suggest a model of the WRKY-HSF transcriptional regulatory cascade for regulating heat stress in strawberry. In response to heat stress, FvHsfB1a acts as a positive regulator to improve thermotolerance by modulating ROS scavenging, enhancing antioxidant enzyme activity, and being directly regulated by the FvWRKY75 TF. These results provide physiological and molecular evidence highlighting the significance of FvHsfB1a in plant responses to heat stress.
4. Materials and Methods
4.1. Plant Materials, Growth Conditions, and Stress Treatment
Diploid strawberry cultivar ‘Ruegen’ (Fragia vesca), Nc89 tobacco (Nicotiana tabacum), and Arabidopsis thaliana (Columbia) plants were grown in a growth chamber at 23 °C, a 12 h light (4000 lux)/12 h dark photoperiod, and 60% relative humidity in Anhui Agricultural University (Hefei, China). The leaf, stem, root, flower, green fruit, and mature fruit of ‘Ruegen’ plants were harvested and used to detect the tissue specificity of the FvHsfB1a gene. The two-month-old seedings were subjected to 42 °C treatment in an artificial climate incubator, and mature leaves were collected at a series of time points (0 h, 3 h, 6 h, 12 h, 24 h, and 48 h).
4.2. Structure and Sequence Analysis of FvHsfB1a
The CDS sequence of FvHsfB1a (XM_004288037.1) was collected from NCBI (www.ncbi.nlm.nih.gov, accessed on 10 April 2022) and cloned from ‘Ruegen’ cDNA using the specific primers (Supplemental Table S1). DANMAN (version 6.0) software was used to analyze the homologs of FvHsfB1a and other species (accessed on 20 May 2022). MEGA (version 7.0) software was used to construct the phylogenetic tree by the maximum-likelihood (ML) method (accessed on 20 May 2022).
4.3. Subcellular Localization of FvHsfB1a Protein
The CDS region without the stop codon of FvHsfB1a was cloned and used to construct the fusion vector 35S::FvHsfB1a-eGFPm and then introduced into the A. tumefaciens GV3101 strain to infiltrate the tobacco leaves. Leica fluorescence confocal microscopy (Leica TCS SP8) was used to observe the GFP signal. The subcellular localization assays were used according to the methods of a previous study [58]. The primers used are listed in Table S1.
4.4. Genetic Transformation of Arabidopsis
The CDS sequence of FvHsfB1a was cloned and inserted into the pRI101-AN vector to construct the pRI101-FvHsfB1a fusion vector. The fusion vector was transferred into the A. tumefaciens GV3101 strain and then transferred into the Arabidopsis Columbia-0 (Col-0, abbreviated WT) plants at flower budding by the floral dip method [59]. T3 homozygous transgenic lines were measured by PCR and qRT-PCR with the specific primers, and used for further tests. All of the primers used are listed in Table S1.
4.5. RNA Extraction and RT-qPCR
The CTAB methods were used to isolate the total RNA of strawberry and Arabidopsis plants. Each cDNA sample was reverse-transcribed and then used for quantitative real-time PCR (qRT-PCR) according to the established protocols. Three technical replicates of each sample were analyzed for qRT-PCR. The primers used are listed in Table S1.
4.6. Heat Stress Assay of Transgenic Arabidopsis Plants
To test the function of FvHsfB1a and FvWRKY75 genes, WT, FvHsfB1a-OE, and FvWRKY75-OE lines were treated with heat stress. At first, seeds of WT and FvHsfB1a-OE lines were treated in a chamber at 37 °C and 42 °C in darkness for 7 days for heat stress. After heat treatment, all Arabidopsis seeds were transferred into a growth chamber for 4 days under normal growing conditions (23/18 °C day/night), and then the germination rates were measured. Meanwhile, seeds of FvHsfB1a-OE lines and WT were sterilized and seeded on an MS medium plate. A sample of 7-day-old Arabidopsis seedlings was subjected to heat stress at 45 °C for 3 h, and then underwent recovery for 5 days under normal conditions to record phenotypes and count survival rates. Furthermore, 3-week-old WT, FvHsfB1a-OE, and FvWRKY75-OE lines were subjected to heat stress at 42/37 °C (day/night) for 7 days, and samples were collected to measure the physiological parameters. A sample of 3-week-old plants was subjected to heat stress at 42 °C for 3 h, and the leaves were harvested to analyze the expression profiles of related genes.
4.7. Yeast One-Hybrid Assay
The CDS region of FvWRKY75 (XM_004310052.2) was cloned and inserted into the plasmid vector pGADT7 with the GAL4 activation domain. PlantCARE online software (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 20 May 2022) was used to identify the cis-acting elements in the promoter region. The promoter region (1500 bp) of FvHsfB1a was cloned and inserted into the pAbAi vector. Yeast one-hybrid assays were conducted according to the previously reported methods [58]. The primers used for the yeast one-hybrid assays are listed in Table S1.
4.8. Dual Luciferase Activity Assay
The CDS region of FvWRKY75 was cloned and inserted into the pGreenII-62SK vector, named pGreenII-62SK-FvWRKY75, and defined as an effector. The promoter region (1500 bp) of FvHsfB1a was inserted into the pGreenII-0800-LUC vector and named pGreenII-0800-FvHsfB1a, which acts as a reporter. Dual luciferase activity assays were conducted according to the previously reported methods [58]. All of the primers used are listed in Table S1.
4.9. Data Analysis
The data are the average of three repeated experiments and are expressed as the mean ± SD. The statistical analysis was determined by using SPSS system (version 20) software. Duncan’s multiple range test was used to identify significant differences among treatment means (* p < 0.05, ** p < 0.01, and *** p < 0.001).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Choudhury F.K. Rivero R.M. Blumwald E. Mittler R. Reactive oxygen species, abiotic stress and stress combination Plant J.20179085686710.1111/tpj.1329927801967 · doi ↗ · pubmed ↗
- 2Wahid A. Gelani S. Ashraf M. Foolad M.R. Heat tolerance in plants: An overview Environ. Exp.20076119922310.1016/j.envexpbot.2007.05.011 · doi ↗
- 3Hasanuzzaman M. Nahar K. Alam M.M. Roychowdhury R. Fujita M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants Int. J. Mol. Sci.2013149643968410.3390/ijms 1405964323644891 PMC 3676804 · doi ↗ · pubmed ↗
- 4Gechev T. Petrov V. Reactive oxygen species and abiotic stress in plants Int. J. Mol. Sci.202021743310.3390/ijms 2120743333050128 PMC 7588003 · doi ↗ · pubmed ↗
- 5Zandalinas S.I. Fritschi F.B. Mittler R. Global warming, climate change, and environmental pollution: Recipe for a multifactorial stress combination disaster Trends Plant Sci.20212658859910.1016/j.tplants.2021.02.01133745784 · doi ↗ · pubmed ↗
- 6Zhu J.K. Abiotic stress signaling and responses in plants Cell 201616731332410.1016/j.cell.2016.08.02927716505 PMC 5104190 · doi ↗ · pubmed ↗
- 7Erpen L. Devi H.S. Grosser J.W. Dutt M. Potential use of the DREB/ERF, MYB, NAC and WRKY transcription factors to improve abiotic and biotic stress in transgenic plants Plant Cell Tissue Organ Cult.201713212510.1007/s 11240-017-1320-6 · doi ↗
- 8Wang W. Vinocur B. Shoseyov O. Altman A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response Trends Plant Sci.2004924425210.1016/j.tplants.2004.03.00615130550 · doi ↗ · pubmed ↗