Unclassified Chromosomal Abnormalities as an Indicator of Genomic Damage in Survivors of Hodgkin’s Lymphoma
Sandra Ramos, Bertha Molina, María del Pilar Navarrete-Meneses, David E. Cervantes-Barragan, Valentín Lozano, Sara Frias

TL;DR
This study shows that Hodgkin’s lymphoma survivors have higher levels of chromosomal abnormalities after treatment, which could predict future cancer risks.
Contribution
The study identifies unclassified chromosomal abnormalities as potential early indicators of secondary cancer risk in Hodgkin’s lymphoma survivors.
Findings
Unclassified chromosomal abnormalities (UnCAs) were significantly higher in Hodgkin’s lymphoma patients than in healthy controls.
UnCA levels increased one year after treatment, driven by free chromatin and micronuclei clusters.
These abnormalities may serve as early biomarkers for secondary cancer risk in survivors.
Abstract
This study examined unclassified chromosomal abnormalities (UnCA) in patients with Hodgkin’s lymphoma (HL) before, during, and after anticancer treatment. HL is a rare cancer affecting 2–4 people per 100,000 individuals annually. Although treatment is highly effective, survivors face an increased risk of developing other cancers. In this study, we analyzed blood samples from HL patients and survivors at different stages of treatment, focusing on changes in their chromosomes that do not fit standard categories. These changes include alterations in chromosomes and nuclei. We found that UnCA levels were higher in HL patients than in healthy individuals at all stages; the general UnCA frequency decreased during treatment but increased again one year after treatment, at the expense of two specific UnCAs, free chromatin and micronuclei clusters. Given their persistence and association with…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
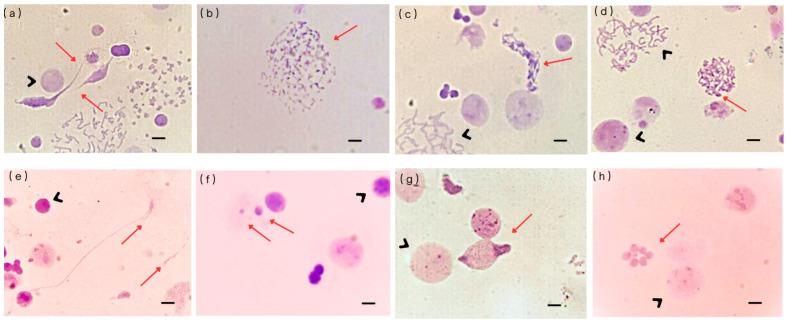 Figure 1
Figure 1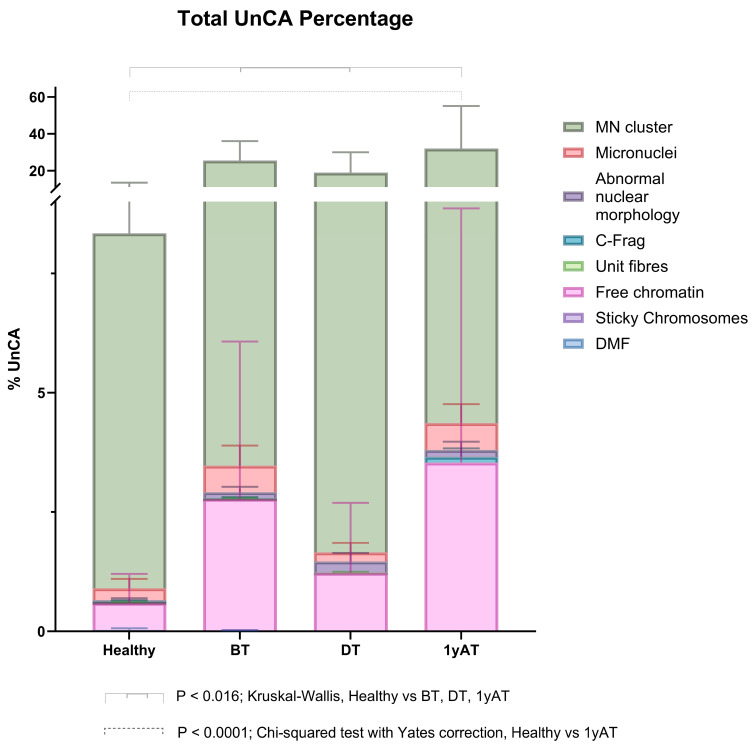 Figure 2
Figure 2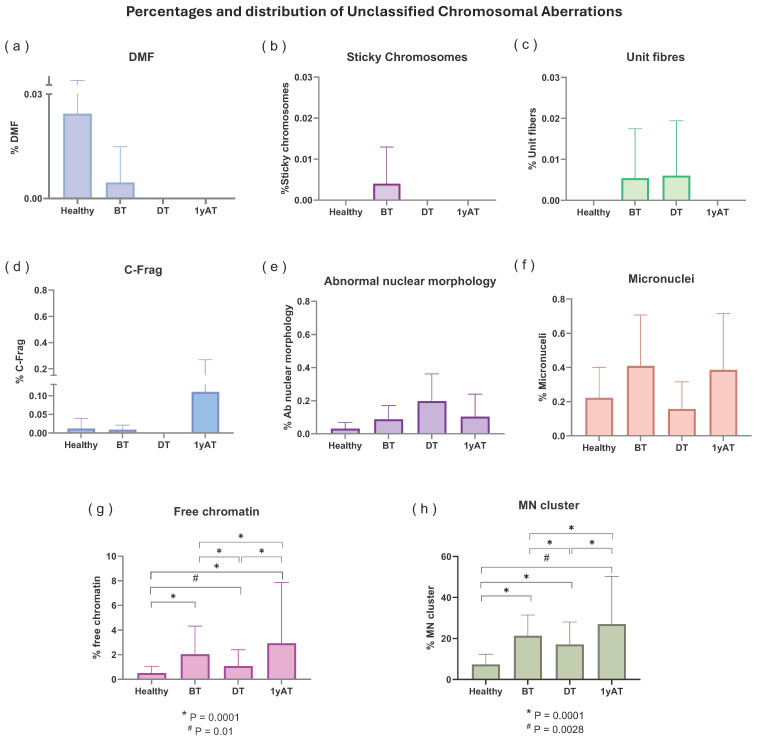 Figure 3
Figure 3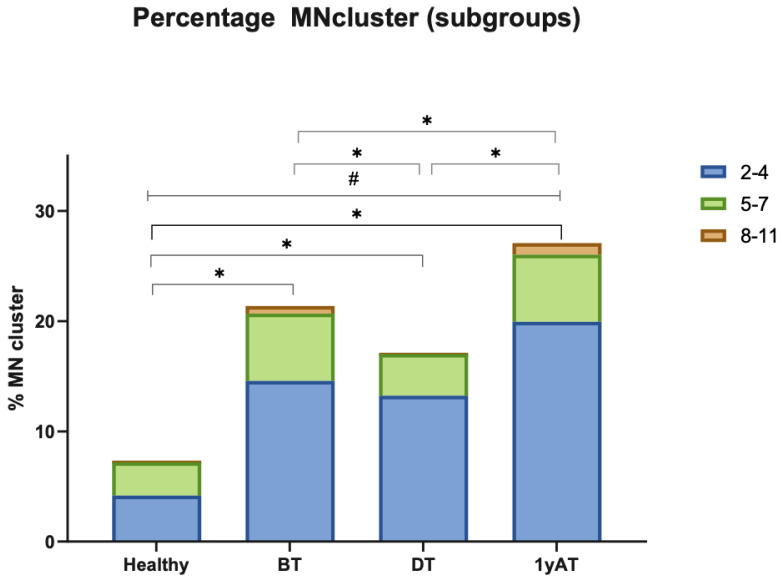 Figure 4
Figure 4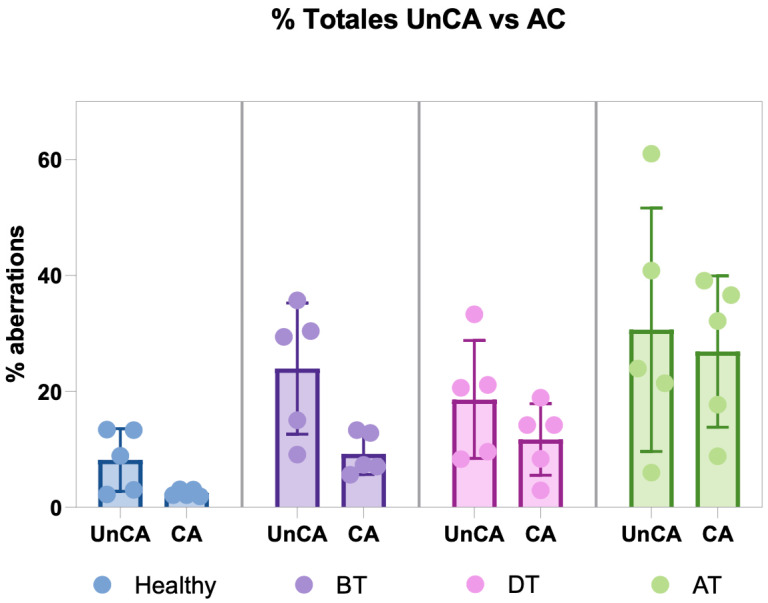 Figure 5
Figure 5- —Recursos fiscales, Instituto Nacional de Pediatría, México
- —Consejo Nacional de Humanidades, Ciencia y Tecnología (CONAHCYT, anteriormente CONACYT)
- —Fondos especiales de apoyo a la Ciencia y Tecnología, Project
- —Fondo Sectorial de Investigación en Salud y Seguridad Social
- —Fundación Miguel Aleman Valdes, Estimulo a la Investigación Médica “Miguel Alemán Valdes”
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDNA Repair Mechanisms · Lymphoma Diagnosis and Treatment · Carcinogens and Genotoxicity Assessment
1. Introduction
Hodgkin’s lymphoma (HL) is one of the most common pediatric cancers, accounting for approximately 4.9% of cases in children under 15 years of age [1]. HL is a hematological malignancy with distinct characteristics, including its frequent occurrence in young adults and the presence of rare malignant Reed–Sternberg cells—multinucleated, B-lymphocyte-derived cells—within a microenvironment rich in immune effector cells [2]. The global incidence rate of HL is approximately 1.2 per 100,000 individuals, with higher rates observed in Western Europe (3.0 per 100,000) [3]. Notably, HL has one of the highest survival rates among cancers, with a 5-year survival exceeding 90% following treatment [4,5]. The treatment is based on the administration of chemotherapy, mainly using ABVD (Adriamycin, Bleomycin, Vinblastine, and Dacarbazine) alone or in association with radiotherapy [5,6]. Chemotherapeutic schemes are not specific for neoplastic cells; so, there may be cell damage and genotoxicity in normal tissues, indeed chromosomal instability has been reported in HL survivors treated with ABVD chemotherapy, represented as non-clonal chromosomal aberrations (NCCAs), both numerical and structural [7,8]. Chromosomal instability (CIN) plays a critical role in the generation of novel genomes and karyotypes, leading to a cell population with extensive genomic heterogeneity, an ideal substrate for malignant transformation [9]. Over the long term, this genomic instability can contribute to the development of second malignant neoplasms (SMNs). SMNs are a significant cause of mortality among HL survivors [10]. The incidence of SMNs is estimated to be approximately 14–17% at 20 years post-treatment, increasing to over 20% at 30 years following initial therapy for the primary tumor [4]. This elevated risk is closely associated with the presence of NCCAs in survivors, as these chromosome aberrations (CAs) are indicative of CIN, a widely recognized hallmark of cancer cells. NCCAs contribute significantly to the dynamic nature of the genome during tumorigenesis, with each aberration offering a distinct evolutionary potential. This genomic variability serves as the foundation for the macroevolutionary phase of cancer progression, enabling the emergence of more aggressive or resistant clones [11].
Genome rearrangements resulting from CIN are not confined to the mitotic phase—where chromosomal abnormalities are most commonly observed—but can also arise during other stages of the cell cycle. In this context, additional forms of genomic alterations, referred to as chromosomal/nuclear unclassified chromosomal alterations (UnCAs), have been identified. Although long regarded as cytogenetic artifacts, UnCAs have gained recognition following studies analyzing the responses of oncological cell lines to antineoplastic agents [12]. These investigations have characterized distinct types of UnCAs, which are now considered manifestations of stochastic genomic alterations (NCCAs). Their presence is increasingly recognized as an indicator of genomic instability [13,14]. Heng et al. [15] identified a range of chromosomal/nuclear UnCAs and demonstrated a significant increase in these aberrations in both experimental and clinical samples exposed to genotoxic agents. The identified UnCAs included free chromatin, defective mitotic figures (DMFs), sticky chromosomes, unit fibers, chromosome fragmentation (C-Frag), micronuclei (MN), micronuclei clusters (MN clusters), and abnormal nuclear morphology. Although some of these chromosomal/nuclear UnCAs are extremely damaged genomes, such as chromosomal fragmentation or MN clusters, some of them may retain the ability to survive stress and maintain proliferative capacity. The progeny of these cells, carrying chaotic genomes, significantly contribute to karyotypic heterogeneity, which serves as the substrate for cellular macroevolution toward cancer [12,15,16,17].
Despite their potential relevance, chromosomal/nuclear UnCAs remain poorly characterized. To date, only micronuclei (MN) have been extensively studied and validated as reliable indicators of DNA damage and mitotic defects [18,19,20]. In contrast, other types of UnCAs lack a comparable body of research. Therefore, it is essential to conduct studies that establish baseline frequencies in healthy individuals and assess how these NCCA behave under genotoxic stress. Cancer survivors represent a highly relevant study population due to the lasting side effects that treatment-induced stress can have on their non-cancerous cells. Additionally, studying these patients offers a unique opportunity to explore the in vivo generation of UnCAs as a response to therapeutic stress.
The aim of this longitudinal study is to determine the frequencies of chromosomal/nuclear UnCAs in lymphocytes, non-cancerous somatic cells, from patients with HL, at three sampling times, before, during and after treatment with ABVD chemotherapy/radiotherapy, and to compare the frequencies of conventional CA, reported in a previous longitudinal study [7], with chromosomal/nuclear UnCAs, to assess their potential as complementary markers of genomic instability induced in vivo.
2. Materials and Methods
2.1. Study Population
HL survivors were recruited from the Survivor Clinic of the National Institute of Pediatrics and the National Cancer Institute in Mexico City. A total of five HL patients participated in this study, each providing peripheral blood samples at three time points: before chemotherapy (BT), during chemotherapy (between the third and fourth cycles; DT), and one year after completing treatment (1yAT). At the time of manuscript preparation, all participants were in remission, with no evidence of relapse or secondary malignancies. In addition, a control group of five healthy young individuals (mean age: 23.6 years) was recruited for comparative analysis.
This study was approved by the Ethics Review Board of the National Institute of Pediatrics. All participants provided written informed consent and completed a health and lifestyle questionnaire, which included information on alcohol consumption and smoking habits. To be eligible, the patients had to be diagnosed with Hodgkin’s lymphoma (HL) based on strict clinical and laboratory criteria and selected to receive ABVD chemotherapy and radiotherapy. The HL patients received between 6 and 8 cycles of ABVD. Additionally, the patients underwent field radiotherapy using a Clinac 2100D/C linear accelerator, with doses ranging from 30 to 45 Gray (Gy) of X-rays, delivered over 15 to 30 daily sessions. No participants were excluded from this study, as none reported the use of illicit or pharmaceutical drugs, had received a blood transfusion within six months prior to enrollment, or had a history of relapse or secondary malignancy.
2.2. Lymphocyte Culture
Approximately 4 mL of peripheral blood was collected via venipuncture by a certified phlebotomist. The blood samples were cultured for 48 h in RPMI medium (Gibco^TM^, Grand Island, NY, USA), supplemented with phytohemagglutinin (PHA-M; Gibco^TM^, Grand Island, NY, USA) to stimulate lymphocyte proliferation. After culture, colchicine [0.08 µg/mL] (Sigma, St. Louis, MO, USA) was added three hours prior to harvesting. The cells were then exposed to a hypotonic treatment with KCl [0.075 M] (Sigma, St. Louis, MO, USA) for 45 min, followed by washing and fixation using Carnoy’s fixative (methanol/acetic acid, 3:1 Merck& Co. Inc., Rahway, NJ, USA). Slides previously processed for M-FISH and analyzed as described by Ramos et al. [7] were reused by first removing the coverslips and Vectashield mounting medium with three washes in 2X SSC (Sigma, St. Louis, MO, USA). The slides were then dehydrated in graded ethanol (Merck& Co. Inc., Rahway, NJ, USA) solutions (70%, 80%, and 100%) for 2 min each. Finally, the slides were stained using Wright–Giemsa stain (Gibco^TM^, Grand Island, NY, USA), following standard protocols and analyzed for UnCAs using a Zeiss Axiolab 5 microscope at 100× magnification (Zeiss, Oberkochen, Germany).
2.3. Cell Analysis
For each sample, the number and types of UnCAs were recorded, 3105–7760 nuclei were analyzed, and the results were expressed as a percentage of UnCAs. To ensure objectivity, the slides were coded by an independent individual not involved in this study, allowing for blinded analysis. In cases of uncertainty regarding the classification of any UnCA, a second reviewer was consulted to confirm the abnormality type. Following the criteria described by Heng et al. [15], eight distinct types of UnCAs were identified in cultures prepared using conventional cytogenetic methods (including hypotonic treatment, fixation, and air-drying): free chromatin; defective mitotic figures (DMFs); sticky chromosomes; unit fibers; chromosome fragmentation (C-Frag); abnormal nuclear morphology; micronuclei (MN) and MN clusters. These alterations are illustrated in Figure 1.
Definition of Unclassified Chromosomal Alterations [15]:
- (a)Free chromatin: These structures consist of interphase chromatin that is not enclosed by a nuclear envelope. They have been observed in various cell lines exposed to chemotherapeutic agents, showing a clear dose–response relationship [12]. Their formation is likely related to nuclear envelope instability, primarily because of underlying genomic instability. Another possible origin of free chromatin is the disruption of micronuclei (MN), which has been identified as one of their possible fates [20].
- (b)Defective mitotic figures (DMFs): These structures are characterized by abnormal mitotic figures in which chromosomes and chromatin fibers coexist, indicating incomplete or faulty chromatin condensation. The primary mechanism underlying DMF formation involves defects in chromatin condensation combined with G2–M checkpoint failure. DMFs have been frequently observed in oncological cell lines, supporting the hypothesis that aberrant chromosomal condensation contributes to cancer development.
- (c)Sticky chromosomes: These aberrations involve chromosomes that adhere to one another, entangled by chromatin fibers, often appearing as fuzzy or clumped structures. Their formation is linked to abnormal methylation patterns that disrupt proper chromatin condensation. There appears to be a strong association between sticky chromosomes and defective mitotic figure formation. Sticky chromosomes are commonly observed in cells treated with ethidium bromide for high-resolution chromosome preparations, or in cells exposed to agents that interfere with DNA replication, methylation, or chromosomal compaction.
- (d)Unit fibers: These are substructures of metaphase chromosomes with a diameter approximately five times smaller than that of a fully condensed chromatid, around 0.4 µm. Unit fibers were first observed by Back [21], who interpreted them as intermediates in chromosome condensation, likely resulting from the coiling of the 250–300 Å solenoid fiber. Unit fibers represent a disruption in normal chromosomal condensation. They have been observed in short-term lymphocyte cultures treated with topoisomerase II inhibitors [15], and their appearance likely reflects incomplete chromatid condensation during metaphase, as result of genotoxic treatment.
- (e)Chromosome fragmentation (C-Frag): C-Frag is a distinct form of cell death during mitosis, characterized by the progressive degradation of chromosomes. It was previously misidentified as chromatin pulverization or premature chromosome condensation. However, unlike apoptosis or mitotic catastrophe, C-Frag arises in response to various cellular stressors, such as gene mutations, infections, drug treatments, and centrosomal dysfunction, and is now recognized as a general stress response mechanism. Some studies suggest that C-Frag may lead to aneuploidy; when it affects a single chromosome, it can trigger genomic chaos, including chromothripsis, ultimately contributing to karyotypic abnormalities [22].
- (f)Micronuclei (MN): These are small nuclei containing one or a few whole chromosomes, chromosomal fragments, or both. They form through various mechanisms, including chromosome displacement during metaphase, slow chromosome separation during transitions from anaphase to telophase, both bipolar and multipolar, fragments from broken chromosome bridges, inheritance of MN from mother cells, separated nuclear fragments during anaphase, extrusion of chromosomes into a mini cell that fuses with a daughter cell, and formation of nuclear buds during the interphase. MN formation is linked to dysfunction in gene networks responsible for DNA damage response, commonly observed in cancers.
It has been shown that the frequency of MN in peripheral blood lymphocytes is elevated after environmental exposure, such as to harmful chemicals and radiation, and its presence is related to genomic instability [15] and pathophysiological conditions such as DNA double-strand breaks, altered DNA repair response, inappropriate DNA replication, treatment of DNA with adduct-forming chemicals, inhibition of microtubule polymerization, and centromere interference [23,24]. It has been suggested that MNs not only indicate genotoxic effects and genomic instability, but also cellular stress response [25] and immune activity followed by DNA damage [26,27,28]. This makes MN valuable biomarkers in genetic toxicology and cancer research [29].
(g)Micronuclei cluster: This is a group of micronuclei of varying sizes that typically form within a single cell, which can be either diploid or polyploid. These clusters can evolve over time through processes of cell fusion and fission, acting as transitional structures during cancer macroevolution (large-scale genomic changes driving cancer progression) [30]. Their formation depends on both the intensity and duration of stress or damaging exposure on cells, which triggers diverse cellular responses, including activation of apoptotic pathways. The formation and evolution of micronuclei clusters may be connected to genomic chaos, reflecting massive genomic rearrangements and instability [23,30]. It has been proposed that MN clusters arise as a consequence of frustrated cell death. These clusters of MN undergo repeated cycles of fusion and fission, reflecting a trial-and-error process in the search for viable genomic configurations. Through these dynamic events, the genomic content is rearranged, leading to the generation of novel, unstable genomes a phenomenon known as genomic chaos. While most MN clusters do not persist, a small subset may survive and initiate macroevolutionary changes that contribute to cancer development [22,23].(h)Abnormal nuclear morphology: It is characterized by markedly altered chromatin condensation patterns within the nucleus and it exhibits a dose-dependent response to chemotherapeutic agents such as doxorubicin. These nuclear changes reflect underlying damage to DNA and disruptions in nuclear organization, often linked to the cytotoxicity of chemotherapy [15].
2.4. Statistical Analysis
Differences in the proportions of UnCAs between HL patients and healthy donors were evaluated using the Chi-squared with Yates’ correction test when comparing two groups, whereas the Kruskal–Wallis and Wilcoxon test were used to compare all groups. Statistical significance was set at p < 0.05.
3. Results
3.1. Patients
Table 1 presents the clinical history of the HL group, including BT, DT, and 1yAT. The average age of the survivors was 25.2 years, with ages at diagnosis ranging from 18 to 33 years.
3.2. Unclassified Chromosomal Aberrations (UnCAs)
3.2.1. UnCAs in Hodgkin’s Lymphoma Patients Versus Healthy Individuals
UnCAs were observed in all analyzed samples, including those from healthy individuals (Table 2). In the healthy group, the percentage of abnormal cells was 8.16. In HL patients, the total percentage of UnCAs was highest in the 1yAT samples, compared to BT and during-treatment, DT, samples. Specifically, the UnCA percentages were 30.62% for 1yAT, 23.92% for BT, and 18.58% for DT. These values were significantly higher than those observed in healthy individuals (p < 0.016; Kruskal–Wallis test) (Table 2, Figure 2).
Among the eight types of UnCA, MN clusters were the most common, representing approximately 90% of the nuclear abnormalities in HL patients and healthy individuals (Table 2, Figure 3); there was at least a 2.3-fold increase in the percentage of MN clusters in all HL patients as compared with healthy individuals. The highest fold increase in MN cluster (3.7) was observed in HL samples collected one-year AT. All values were significantly different (p < 0.013; Kruskal–Wallis test) compared to the values of the MN cluster observed in the healthy individual samples.
We classified MN clusters into three subgroups depending on the number of MN observed: 2–4, 5–7, and 8–11 (Figure 4). In all HL samples, the predominant MN clusters were composed of two to four MN, regardless of the time of sampling.
Free chromatin was the second most common type of UnCAs, accounting for 5–8% of all UnCAs observed across all sample groups, including healthy individuals. Although the difference was not statistically significant, a higher frequency of this UnCA was noted in patient samples (Table 2, Figure 3). Overall, UnCAs in the 1yAT samples showed a significantly increased frequency compared to those in healthy individuals, as well as in the BT and DT samples.
All other types of UnCAs were observed at low frequencies and without significantly differences among sampling time or studied group of individuals. An analysis of these low-frequency UnCAs showed that MN presented with the same pattern of free chromatin and MN clusters although without statistical significance. Also, a non-significant increase in abnormal nuclear morphology was noted in the DT patient group and chromosome fragmentation (C-Frag) frequency was more frequent in samples collected 1yAT, without a statistically significant difference (Figure 3).
3.2.2. UnCA vs. Chromosomal Aberrations
The comparison between UnCA and classical chromosomal aberrations, CAs, was based on the number of aberrant cells, as this is the only feasible standardization for both analyses. This approach was necessary because UnCA quantifies the abnormal cells, whereas CA analysis is traditionally reported as the frequency of aberrations per cell, though it can also be expressed as the percentage of aberrant cells. When comparing the total number of cells with UnCA across all study groups, including healthy individuals and HL patients at three different sampling time points, a parallel pattern emerged, mirroring the trends observed in the CA analyses. Consistent with previous CA findings, a lower frequency of CA and chromosomal/nuclear UnCAs was detected in DT versus AT, indicating that the most severely damaged cells may be eliminated by chemotherapy and therefore underrepresented in cytogenetic assessments [7]. Interestingly, the aberrant cells, both with UnCAs and CA showed the highest frequency recorded one year after treatment; both analytical methods corroborated this trend, suggesting a temporal pattern in the occurrence of genomic aberrations and reflecting the dynamic nature of genomic instability across different stages of treatment and recovery in HL patients (Figure 5, Table 3).
4. Discussion
In this study, we investigated the quantity and types of chromosomal/nuclear UnCAs in lymphocytes from HL patients, assessed before, during, and after anticancer treatment, to evaluate whether these underreported alterations are induced by radio/chemotherapy, and to determine whether their occurrence parallels that of classical CAs, which are commonly used to assess the genotoxic effects of such treatments. Our results showed that when comparing the total number of cells with UnCAs across all study groups, including healthy individuals and HL patients at three different sampling times, the emerging pattern mirrors the trends observed with the classical CA analyses. These results suggest that anticancer-therapy-induced stress leads to chromosomal and nuclear alterations, evident in different phases of the cell cycle. Furthermore, chromosomal/nuclear UnCAs, similar to classical CAs, may serve as valuable indicators for detecting chromosomal instability (CIN) resulting from exposure to genotoxic agents. These genotoxic-stress-induced NCCAs lead to genomic heterogeneity in the hematopoietic stem cells, the essential factor for macroevolution toward cancers, and may partially explain the development of secondary new blood cancers in HL survivors.
As regards the basal frequencies of chromosomal/nuclear UnCA, we found a percentage of 8.16 total UnCAs in healthy individuals, revealing that the most represented type is MN clusters, with a mean of percentage of 7.36, and showing extremely low percentages of the other types of UnCAs.
Chromosomal/nuclear UnCAs showed a higher level of genomic damage in patients with HL compared to healthy individuals, even prior to the initiation of treatment. This observation suggests that the disease itself, as in other cancers [31,32], may be inherently associated with increased genomic instability in HL patient’s normal lymphocytes and this is independent of Hodgkin’s and Reed–Sternberg cancer cells in HL, which are known to undergo extensive genomic modifications [33,34].
When UnCA levels were analyzed at the DT and 1yAT time points, we found that despite the therapeutic success in eliminating cancer cells, the total number of UnCAs was significantly higher in lymphocytes from HL survivors one year after treatment (1yAT). This finding underscores the persistence of damaged cells with a relative survival advantage (“better fitness”), which may sustain the substantial high burden of treatment-induced genomic damage in the patients’ non-cancerous cells.
In addition, we compared the analysis of UnCAs with that of classical CAs as a response to anticancer therapy in patients with HL. This comparison was motivated by previous findings with classical CAs analysis, which showed a low proportion of genomic damage occurring DT, a period when cells are directly exposed to genotoxic agents [7]. We aimed to investigate whether substantial genomic damage might be detectable at other stages of the cell cycle, beyond mitosis. Interestingly, the percentages of aberrant cells obtained from both analyses revealed parallel trends. In both cases, a lower level of genotoxic damage was observed DT as compared with the levels detected 1yAT. This may be attributed to the high rate of cell death induced by chemotherapy, which eliminates the most severely damaged cells, rendering them undetectable by either method. In contrast, in the 1yAT phase, surviving cells accumulate additional alterations, reflecting CIN, a driver of cancer evolution [13,14,35]; in this context, previous studies have reported that abnormalities in nuclear architecture, such as UnCAs, are associated with cellular dysfunction, impaired molecular maintenance mechanisms, and, ultimately, chromosomal breaks that contribute to genomic instability [36].
Our results indicate that MN clusters were the most frequently observed UnCAs (Table 2, Figure 2, Figure 3 and Figure 4). Notably, these clusters showed the greatest differences in frequency between healthy individuals and HL patients and across the different sampling times. Specifically, HL patients exhibited a 2.3-fold increase in MN clusters compared to healthy individuals. Among the HL patient groups, the highest increase, a 3.7-fold rise, was observed in samples 1yAT (Figure 3). Indeed, these types of UnCAs were less frequently observed DT, and those detected at this time point typically involved smaller clusters containing only 2–4 micronuclei per cluster, while the clusters with 8–11 MN re-appeared 1yAT (Figure 3). This supports the idea that the most severely damaged genomes are eliminated during therapy, and that surviving cells with CIN undergo genomic rearrangement. This process facilitates the emergence of NCCAs, which may become detectable at later time points. Notably, we observed a resurgence of these 8–11 MN clusters 1yAT. It is hypothesized that MN clusters act as intermediary structures for genomic reorganization, promoting the formation of new genomic architectures such as genomic chaos, conducive to macroevolutionary selection [11]. Interestingly, chromoplexy (a hallmark of genomic chaos) was detected in the analysis with M-FISH of the HL survivors at 1yAT [7]. This temporal coincidence with the peak of MN clusters suggests a possible mechanistic link, positioning MN clusters as a possible precursor of chromoplexy, and genomic chaos as a potential outcome of treatment-induced genomic remodeling.
The genomic heterogeneity induced by anticancer agents in lymphocytes, and possibly in all somatic cells of cancer survivors, is of critical importance, as each altered genome constitutes a distinct information system with a unique potential for survival. If any of these genomic variants acquire a selective growth advantage, they may initiate a transition from punctuated macroevolution to gene-mutation-driven microevolution [11]. This process could ultimately lead to the development of secondary malignancies, either hematologic or solid tumors, a phenomenon well documented among survivors of Hodgkin’s lymphoma (HL) [37].
This information underscores the lasting impact of anticancer therapies on genomic stability and highlights the potential utility of UnCAs as markers for assessing treatment-related genomic damage over time. Chromosomal/nuclear UnCAs analysis revealed genomic alterations within interphase nuclei, thereby complementing the damage detected by classical CA. Further research is warranted to investigate this phenomenon, which remains detectable even in samples collected a decade after treatment, such as those from patients treated with MOPP, as reported by Salas et al. and Bilban-Jakopin [38,39]. These findings suggest that UnCAs are genome-level alterations that may be initiate a macroevolution steps toward cancer and could serve as a valuable tool for monitoring the long-term genomic effects of anticancer therapies, potentially useful for both treatment strategies and follow-up care for cancer survivors.
5. Conclusions
An analysis of UnCAs across different treatment stages revealed a higher frequency in HL patients than in healthy individuals, even before treatment initiation, suggesting an inherent genomic instability associated with the disease.
The analyses of UnCAs across all study groups, including healthy individuals and HL patients at three different sampling times, revealed a genomic damage pattern that mirrors the trends observed with the classical CA analyses, showing DT with a low percentage of damaged cells as compared with the proportion of cells with cytogenetic abnormalities in the 1yAT samples.
The sustained presence and even the amplification of UnCAs observed 1yAT in blood cells suggest that hematopoietic stem and progenitor cells that survived treatment were able to replenish the hematopoietic compartment, albeit at the cost of carrying sublethal genomic damage. The cell population that survived the stress induced by ABVD chemotherapy and/or radiotherapy 1yAT exhibits pronounced genomic heterogeneity, a well-established driver of cancer progression. This genomic variability may help explain the elevated risk of secondary malignancies, both hematologic and solid tumors, commonly observed in survivors of HL.
This study also highlights the reliability and advantages of UnCAs analysis as a complementary tool to classical cytogenetic methods, providing an accessible and broadly applicable approach for assessing genomic instability.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kaatsch P. Byrne J. Grabow D. Managing a Pan-European Consortium on Late Effects among Long-Term Survivors of Childhood and Adolescent Cancer—The Pancarelife Project Int. J. Environ. Res. Public. Health 202118391810.3390/ijerph 1808391833917926 PMC 8068322 · doi ↗ · pubmed ↗
- 2Connors J.M. Cozen W. Steidl C. Carbone A. Hoppe R.T. Flechtner H.H. Bartlett N.L. Hodgkin Lymphoma Nat. Rev. Dis. Primers 202066110.1038/s 41572-020-0189-632703953 · doi ↗ · pubmed ↗
- 3Sung H. Ferlay J. Siegel R.L. Laversanne M. Soerjomataram I. Jemal A. Bray F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries CA Cancer J. Clin.20217120924910.3322/caac.2166033538338 · doi ↗ · pubmed ↗
- 4Zahnreich S. Schmidberger H. Childhood Cancer: Occurrence, Treatment and Risk of Second Primary Malignancies Cancers 202113260710.3390/cancers 1311260734073340 PMC 8198981 · doi ↗ · pubmed ↗
- 5Farnetani G. Vannucci M. Fino M.G. Cioppi F. Rosta V. Palma M. Tamburrino L. Vinci S. Casamonti E. Degl’Innocenti S. Severe Sperm DNA Fragmentation May Persist for up to 3 Years after Cytotoxic Therapy in Patients Affected by Hodgkin Lymphoma and Non-Hodgkin Lymphoma Hum. Reprod.20243949650310.1093/humrep/dead 26938177083 · doi ↗ · pubmed ↗
- 6Engert A. Younes A. Hodgkin Lymphoma: A Comprehensive Update on Diagnostics and Clinics 2nd ed.Hodgkin Lymphoma: A Comprehensive Overview Springer New York, NY, USA 201510.1007/978-3-319-12505-3 · doi ↗
- 7Ramos S. Navarrete-Meneses P. Molina B. Cervantes-Barragán D.E. Lozano V. Gallardo E. Marchetti F. Frias S. Genomic Chaos in Peripheral Blood Lymphocytes of Hodgkin’s Lymphoma Patients One Year After ABVD Chemotherapy/Radiotherapy Environ. Mol. Mutagen.20185975576810.1002/em.2221630260497 · doi ↗ · pubmed ↗
- 8Frias S. Ramos S. Salas C. Molina B. Sánchez S. Rivera-Luna R. Nonclonal Chromosome Aberrations and Genome Chaos in Somatic and Germ Cells from Patients and Survivors of Hodgkin Lymphoma Genes 2019103710.3390/genes 1001003730634664 PMC 6357137 · doi ↗ · pubmed ↗