Metabolic Variations in Bamboo Shoot Boiled Liquid During Pediococcus pentosaceus B49 Fermentation
Juqing Huang, Meng Sun, Xuefang Guan, Lingyue Zhong, Jie Li, Qi Wang, Shizhong Zhang

TL;DR
This study examines how fermentation with Pediococcus pentosaceus B49 changes the chemical makeup of bamboo shoot boiled liquid, revealing key metabolic shifts that could help create functional food products.
Contribution
The study identifies specific metabolite changes and key metabolic pathways affected by Pediococcus pentosaceus B49 fermentation in bamboo shoot boiled liquid.
Findings
Fermentation significantly alters metabolite profiles, with the most changes occurring in the first 24 hours.
Fermented samples show increased levels of certain flavones and coumarins, while bitter peptides decrease significantly.
Tyrosine and arginine/proline metabolism pathways are most affected by the fermentation process.
Abstract
Bamboo shoot boiled liquid (BSBL), a processing byproduct containing soluble proteins, peptides, amino acids, carbohydrates, and phenolics, is typically discarded, causing resource waste and environmental issues. This study analyzed metabolic changes in BSBL during Pediococcus pentosaceus B49 fermentation. The result of partial least squares discriminant analysis (PLS-DA) revealed significant metabolite profile differences across fermentation times (0 h, 24 h, 48 h, 72 h, 96 h). The most substantial alterations occurred within the first 24 h, followed by stabilization. Compared to unfermented BSBL, fermented samples exhibited significantly elevated signal intensities for 5,7-dimethoxyflavone, cinnamic acid, 3,4-dihydro-2H-1-benzopyran-2-one, 6,8-dimethyl-4-hydroxycoumarin, and 2-hydroxycinnamic acid (p < 0.05), showing upward trends over time. Conversely, (+)-gallocatechin intensity…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
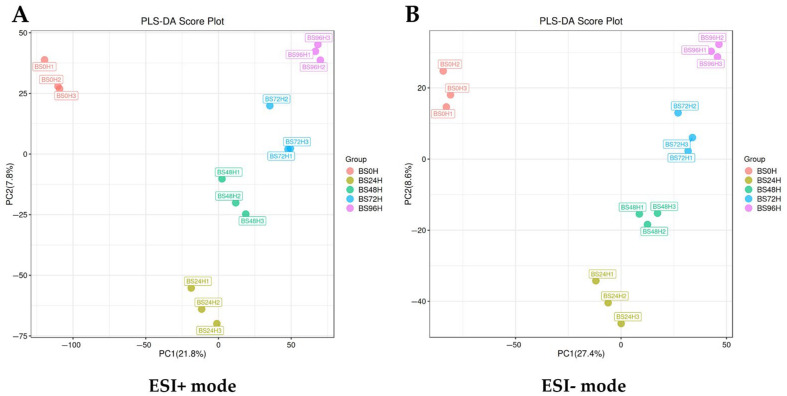 Figure 1
Figure 1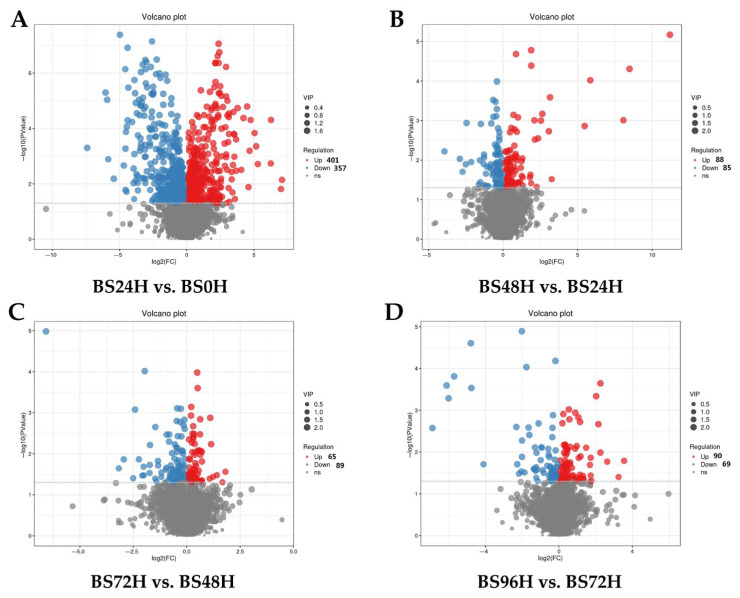 Figure 2
Figure 2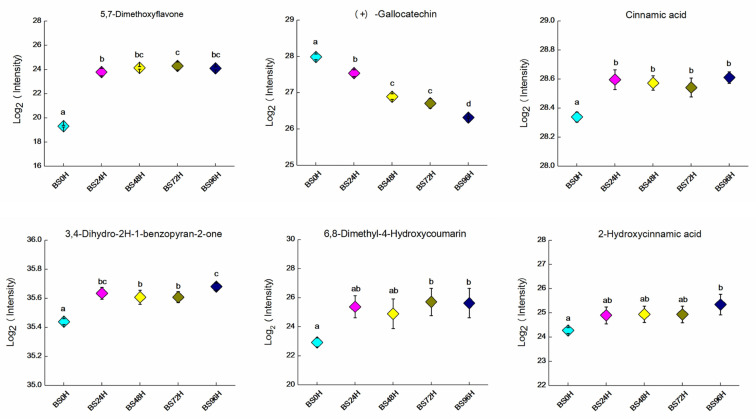 Figure 3
Figure 3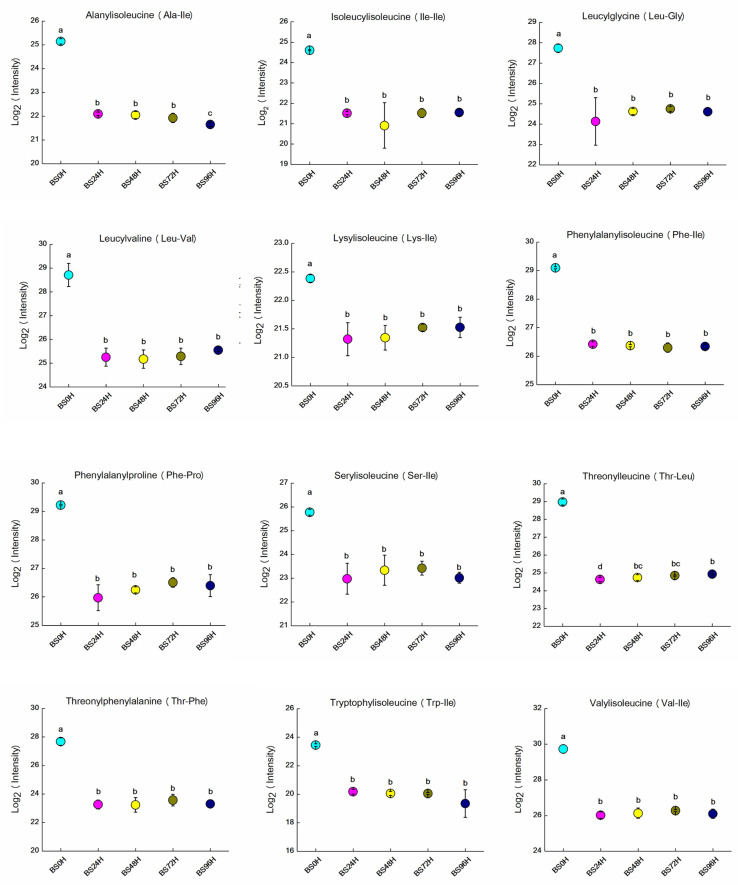 Figure 4
Figure 4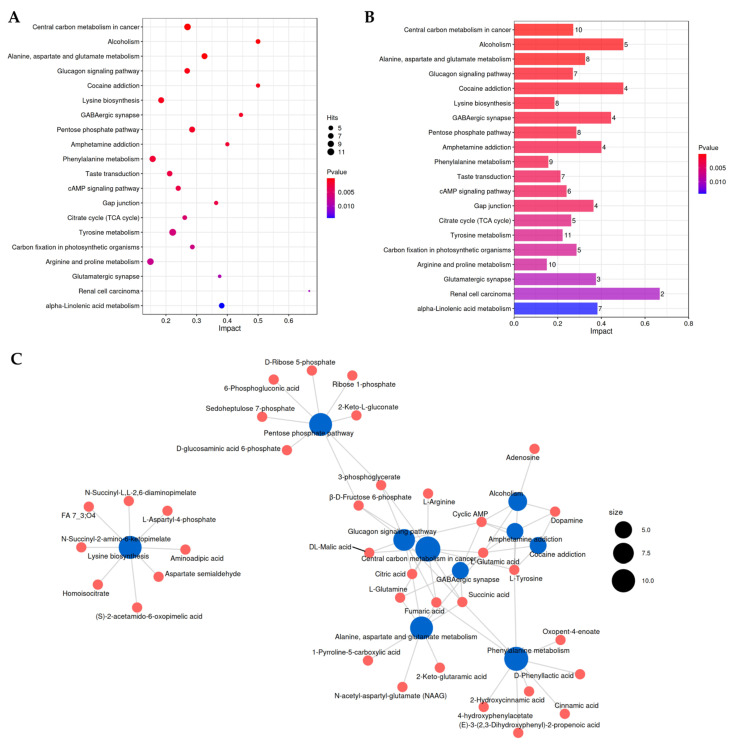 Figure 5
Figure 5- —Fujian Natural Science Foundation
- —Fujian Academy of Agricultural Sciences’ External Cooperation Project
- —Regional Development Project of Fujian Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAdvanced Cellulose Research Studies · Bamboo properties and applications · Biofuel production and bioconversion
1. Introduction
Bamboo shoots, the edible young culms of bamboo plants, are a traditional and nutritious vegetable widely consumed in Asia, particularly in China [1]. Rich in dietary fiber, vitamins (e.g., B1, B2, and C), minerals (e.g., potassium and calcium), and bioactive compounds (e.g., phenolics and flavonoids), bamboo shoots exhibit numerous health benefits, including antioxidant, anti-inflammatory, and hypolipidemic effects [2]. Reflecting its pivotal role in the global bamboo economy, China stands as the world’s largest producer, yielding approximately 1.03 million tons of bamboo shoots annually, with an associated industry value exceeding USD 21.7 million [3]. Despite their nutritional merits, fresh bamboo shoots are highly perishable, necessitating processing to ensure shelf stability and year-round availability [4]. A substantial portion of the harvest enters the industrial processing stream. Industry reports and studies indicate that approximately 60% of China’s bamboo shoots are processed commercially, primarily into forms such as canned (boiled), dried, marinated, fermented products, and medicinal extracts, while the remaining 40% are consumed fresh [5]. Among these processed forms, boiled bamboo shoots, predominantly packaged in cans for domestic and international markets, represent a major industrial product category. A critical environmental challenge arising from this large-scale processing, particularly the industrial boiling step essential for preservation, is the generation of vast volumes of nutrient-rich wastewater, known as bamboo shoot boiled liquid (BSBL). At the industrial scale, it is estimated that processing one ton of bamboo shoots generates approximately 2 tons of BSBL [6]. Extrapolating from China’s annual processed shoot volume (roughly 0.618 million tons, based on 60% of 1.03 million tons), this translates to an industrial-scale generation exceeding 1.2 million tons of BSBL annually. This represents a significant waste stream distinct from the comparatively negligible quantities produced sporadically at the domestic level. Currently, this nutrient-laden effluent is predominantly treated as wastewater or discarded directly, leading to substantial resource wastage of valuable compounds and posing considerable environmental burdens due to its organic load and potential eutrophication effects [5]. BSBL contains soluble proteins, peptides, free amino acids, carbohydrates, and phenolic compounds leached from bamboo shoots during boiling [6]. We assumed that BSBL possesses potential prebiotic and antioxidant properties, making it a promising substrate for microbial fermentation. However, current utilization of BSBL remains limited. Exploring microbial fermentation to valorize BSBL could enhance its functional properties and enable its application in functional foods or bioactive ingredient production.
Pediococcus pentosaceus, a widely recognized lactic acid bacteria (LAB), is frequently applied in food fermentation thanks to its strong acid production capability and excellent tolerance, which significantly improve the flavor and quality of fermented products [7,8]. However, systematic studies focusing on metabolic changes during the fermentation of bamboo shoots or their by-products (i.e., BSBL) by P. pentosaceus remain insufficient. Employing P. pentosaceus to ferment BSBL presents a promising strategy to valorize this byproduct. The process aligns with circular economy goals by converting waste into high-value ingredients for nutraceuticals or functional beverages. Untargeted metabolomics can elucidate dynamic metabolic changes during fermentation, providing insights into strain-specific pathways and optimizing fermentation conditions for targeted bioactivity enhancement.
P. pentosaceus B49 is a strain that was isolated from human colostrum [9]. Our previous study has demonstrated its ability to relieve constipation in mice [9]. However, the effects of this strain fermentation on BSBL have not been investigated. This study aimed to investigate the changes in metabolite profiles during BSBL fermentation by P. pentosaceus B49. We utilizes BSBL as the raw material, with P. pentosaceus B49 as the fermentation strain, employing untargeted metabolomics via ultrahigh performance liquid chromatography (UHPLC-MS/MS) to systematically assess metabolite changes at different fermentation stages (0 h, 12 h, 24 h, 48 h, 72 h). Through multivariate statistical analysis, significant metabolites are identified, and combined with KEGG pathway enrichment analysis, the key metabolic pathways and changes in active substances during fermentation are clarified. The findings will enhance the understanding of metabolic regulation mechanisms in this process, providing a theoretical foundation for developing functional bamboo shoot fermentation products, and offering new pathways for the advanced development and utilization of bamboo shoot resources.
2. Materials and Methods
2.1. Materials and Reagents
Fresh spring shoots of moso bamboo (Phyllostachys edulis) were purchased from local markets. LC-MS-grade methanol was purchased from Fisher Scientific UK Ltd. (Loughborough, UK). 2-Amino-3-(2-chloro-phenyl)-propionic acid was obtained from Shanghai Aladdin Biochemical Technology Co., Ltd. (Shanghai, China). LC-MS-grade acetonitrile was purchased from UK Fisher Scientific Ltd. (Loughborough, UK). Formic acid was obtained from TCI (Shanghai) Development Co., Ltd. (Shanghai, China). Ammonium formate was obtained from Sigma-Aldrich (Shanghai) Trading Co., Ltd. (Shanghai, China). Ultrapure water was generated using a Milli-Q system (Millipore, Bedford, MA, USA).
The P. pentosaceus B49 strain was obtained from the human colostrum according to our previous procedure [9]. It was preserved at the China General Microbiological Culture Collection Center (CGMCC, No. 15957). The strain was cultured in MRS broth at 37 °C for 24 h before fermentation.
2.2. Preparation of BSBL Broth for Fermentation
Fresh spring shoots of moso bamboo (Phyllostachys edulis) were peeled and washed, then the edible portion was sliced into approximately 1 cm × 2 cm × 0.3 cm pieces. The slices were heated in a boiling water bath for 30 min. After cooling, the bamboo shoot boiled liquid (BSBL) was obtained through filtration. Subsequently, the liquid was further concentrated to approximately one-tenth of the fresh weight of the edible portion of bamboo shoots to prepare the BSBL broth, which facilitates enhanced microbial growth. Then, a 5% inoculum of activated P. pentosaceus B49 strain was aseptically added to the BSBL broth within a laminar flow hood. Fermentation broth samples were collected at 0, 24, 48, 72, and 96 h (defined as BS0H, BS24H, BS48H, BS72H and BS96H, respectively), centrifuged (3000 rpm, 4 °C, 15 min), and the supernatants were stored at −20 °C for subsequent untargeted metabolomic analysis.
2.3. Extraction of Metabolites
The metabolites were extracted following a previously described method [10]. Briefly, frozen samples were thawed at 4 °C, vortexed for 1 min to ensure homogeneity, and mixed thoroughly. A precise volume of each sample was transferred into a 2 mL centrifuge tube. A volume of 400 µL of methanol was added to the aliquot, followed by vortexing for 1 min to facilitate metabolite extraction. The mixture was centrifuged at 12,000 rpm and 4 °C for 10 min. The supernatant was carefully transferred to a new 2 mL tube, concentrated under reduced pressure (or nitrogen stream), and dried. The dried residue was reconstituted in 150 µL of 2-chloro-l-phenylalanine (4 ppm) solution (prepared in 80% methanol/water), vortexed to ensure complete dissolution, and filtered through a 0.22 μm membrane. The filtrate was transferred to an LC-MS vial for subsequent analysis.
2.4. UHPLC-MS/MS Detection
The analysis was performed according to established methods [11,12] using a Vanquish UHPLC system (Thermo Fisher Scientific, Waltham, MA, USA) coupled to a Q Exactive Focus mass spectrometer (Thermo Fisher Scientific, Waltham, MA, USA). Chromatographic separation was achieved on an ACQUITY UPLC HSS T3 column (2.1 × 100 mm, 1.8 μm; Waters, Milford, MA, USA) maintained at 40 °C with a flow rate of 0.3 mL/min and injection volume of 2 μL. For positive ion mode (ESI+), mobile phases consisted of 0.1% formic acid in water (A2) and 0.1% formic acid in acetonitrile (B2), while negative ion mode (ESI−) used 5 mM ammonium formate (A3) and acetonitrile (B3). Both modes employed identical gradient programs: 0–1 min at 10% B, 1–5 min linear increase to 98% B, 5–6.5 min hold at 98% B, 6.5–6.6 min rapid return to 10% B, followed by 6.6–8 min re-equilibration. Mass spectrometric detection utilized an ESI ion source operating in full MS-ddMS^2^ mode with alternating spray voltages of +3.50 kV (ESI+) and −2.50 kV (ESI−). Key parameters included: sheath gas at 40 arbitrary units (arb), auxiliary gas at 10 arb, capillary temperature at 325 °C, MS1 scan range m/z 115–1720 with a resolution 70,000 (full width at half maximum, FWHM), and data-dependent MS^2^ scans (top 3 precursors per cycle) at 17,500 resolution with 30 eV normalized collision energy and automatic dynamic exclusion.
2.5. Metabolite Identification and Quantitative Analysis
Raw data were initially converted to the mzXML format using MSConvert from the ProteoWizard software suite (v3.0.8789) [13]. Subsequent data processing was conducted with the R package XCMS (version 3.12.0), which included feature extraction, retention time adjustment, and alignment [14]. Key processing parameters included: ppm set to 15, peakwidth at c(5, 30), mzdiff at 0.01, and centWave as the detection method. Systematic variations were minimized through area normalization. Metabolite identification was achieved by matching accurate mass and MS/MS spectral data against several databases: HMDB (http://www.hmdb.ca, accessed on 20 April 2025), MassBank (https://massbank.eu/MassBank/), KEGG (https://www.genome.jp/kegg/, accessed on 20 April 2025), LipidMaps (http://www.lipidmaps.org, accessed on 20 April 2025), mzCloud (https://www.mzcloud.org, accessed on 20 April 2025), as well as a proprietary metabolite database from Panomix Biomedical Tech Co., Ltd. (Suzhou, China). The molecular weights of metabolites were determined by analyzing the m/z values of precursor ions in the MS data. Molecular formulas were inferred based on ppm tolerance and adduct ion types, followed by database matching. Additionally, MS/MS spectra from the quantitative data table were compared with the characteristic fragment ions and related information for each compound within the databases to confirm metabolite identities.
For multivariate statistical analysis, supervised partial least squares discriminant analysis (PLS-DA) was performed using the ropls package (version 1.22.0) in R (version 3.12.0) [15]. Differential metabolites (DMs) were defined according to criteria including statistical significance (p < 0.05), variable importance in projection (VIP > 1), and specified fold change (FC) thresholds.
2.6. Pathway Analysis
Differential metabolites (DMs) were analyzed for pathway involvement using MetaboAnalyst (Version R 4.0) [16], which integrates robust pathway enrichment and pathway topology analysis. The metabolites identified through metabolomics were subsequently mapped onto KEGG pathways to facilitate the biological interpretation of complex systemic functions. Visualization of these metabolites and their associated pathways was accomplished with the KEGG Mapper tool.
2.7. Statistical Analysis
All experiments were carried out with three independent biological replicates. Data visualization was accomplished using Origin (version 2019) and Microsoft PowerPoint (version 2019). Statistical evaluation of the experimental results was conducted through one-way ANOVA, and subsequent comparisons were made using Duncan’s multiple range test with IBM SPSS Statistics 22. A p-value of less than 0.05 was considered statistically significant.
3. Results and Discussion
3.1. Overall Differences in Metabolite Profiles
As a supervised learning method that emphasizes inter-group variation, PLS-DA was employed to analyze metabolite differentiation among BS0H, BS24H, BS48H, BS72H, and BS96H. PLS-DA analyses were conducted in both ESI+ and ESI− modes, and the results are presented in Figure 1. The results indicated a clear statistical separation among the metabolite profiles of BS0H, BS24H, BS48H, BS72H, and BS96H in both ESI+ and ESI− modes, demonstrating that the metabolite compositions of the fermented samples differed significantly at various time points.
3.2. Volcano Plot Analysis
Volcano plot analyses were employed to visualize differential expression across sequential time comparisons: BS24H vs. BS0H, BS48H vs. BS24H, BS72H vs. BS48H, and BS96H vs. BS72H. These analytical outputs are systematically presented in Figure 2. The comparative analysis of BS0H vs. BS24H (Figure 2A) identified 401 significantly upregulated metabolites compared to 357 downregulated metabolites, highlighting substantial metabolic alterations in BSBL within the initial 24 h period. The BS48H vs. BS24H comparison (Figure 2B) demonstrated 88 upregulated metabolites versus 85 downregulated counterparts. Subsequent analysis of BS72H vs. BS48H (Figure 2C) revealed 65 upregulated metabolites compared to 89 downregulated compounds. The final comparison between BS96H and BS72H (Figure 2D) indicated 90 upregulated metabolites with 69 metabolites exhibiting significant downregulation (p < 0.05). The findings reveal that the most substantial metabolic alterations in BSBL occurred during the initial 24 h of fermentation, while subsequent changes between 24 and 96 h showed an overall stabilization pattern, indicating distinct metabolic phases during the fermentation process.
3.3. Identification of DMs
To further elucidate the metabolic variations in BSBL during the fermentation process of P. pentosaceus B49, we identified and listed the top 30 most significant and relevant differential metabolites (DMs) from the following comparisons: BS24H vs. BS0H, BS48H vs. BS24H, BS72H vs. BS48H, and BS96H vs. BS72H, with their corresponding log_2_ fold change (log_2_FC) values presented in Table 1 and Table 2.
As shown in Table 1, during the initial 24 h period, the primary upregulated DMs were 5,7-dimethoxyflavone; 1,3-nonanediol; (3Z,6Z)-3,6-nonadienal; medicocarpin; N-succinyl-2-amino-6-ketopimelate; 1-methoxy-2-hydroxyanthracene; 6-hydroxymelatonin; D-phenyllactic acid; N-succinyl-L,L-2,6-diaminopimelate; 10-hydroxydecanoic acid; (E)-3-heptenyl acetate; toluene-cis-dihydrodiol; citramalate; 2-inosose; N-acetyl-serylaspartic acid; ornithine; and β-alanine. Of particular note, 5,7-dimethoxyflavone, a naturally occurring flavonoid, has been reported to demonstrate various biological activities, including antioxidant, antidiabetic, anti-obesity, anti-inflammatory, and anticancer properties [17,18,19]. Medicocarpin has demonstrated strong binding ability with angiotensin-converting enzyme 2, a property closely associated with potential anti-COVID-19 activity [20]. The compound 6-hydroxymelatonin is the primary metabolite of melatonin, which is a major biologically active molecule secreted by the pineal gland [21]. It possesses a variety of functions, including free-radical scavenging and the induction of both protective and reparative cellular mechanisms [21]. D-phenyllactic acid, a metabolite produced by lactic acid bacteria, has been demonstrated to exhibit antimicrobial and immunomodulatory activities [22]. Notably, 1,3-nonanediol and (3Z,6Z)-3,6-nonadienal, and (E)-3-heptenyl acetate were identified as flavor substances generated during BSBL fermentation by P. pentosaceus B49 within the initial 24 h period. The DMs showing downregulation in the BS24H vs. BS0H comparison primarily comprised oligopeptides (Table 1). These oligopeptides included: Ile-Val-Gln, Ile-Val-Gly, Phe-Ile, Ala-Ile, Ile-Ile, Trp-Ile, Val-Val-Ala, Val-Ile-Thr, Thr-Leu, and Ile-Val-Val. These peptides may originate from two distinct pathways: microbial degradation of BSBL proteins by P. pentosaceus B49 metabolites, or alternatively, as native components inherent to the BSBL matrix prior to fermentation. The compound 3-phosphoglycerate serves as a central metabolic intermediate in both glycolysis and the Calvin cycle, while 6-phosphogluconic acid operates within the oxidative pentose phosphate pathway, arising through glucose-6-phosphate oxidation [23]. The metabolic depletion of these compounds likely reflects active carbohydrate metabolism by P. pentosaceus B49 during the initial 24 h fermentation phase.
Homodolicholide is reported to be a kind of steroidal plant-growth regulator [24]. The observed accumulation of homodolicholide in BSBL fermented with P. pentosaceus B49 between 24 and 48 h likely stems from the microbial bioconversion of particular bamboo shoot sterols [25]. Alisol C monoacetate is a kind of triterpenoid that is reported to have an anti-liver fibrosis effect [26]. Zhang et al. demonstrated the anti-fatigue effects of a triterpenoid-rich extract obtained from Chinese bamboo shavings, suggesting that these bioactive compounds are inherently present in bamboo plants [27]. In this study, a significant upregulation of alisol C monoacetate and farnesyl acetate was observed during the 24 to 48 h BSBL fermentation process mediated by the P. pentosaceus B49 strain. These metabolic alterations are likely attributable to the bacterial bioconversion of terpenes constituents naturally occurring in bamboo shoots via the enzymatic activities of the P. pentosaceus B49 strain. The content of 7-methylxanthosine was also significantly increased during the 24~48 h fermentation of BSBL by P. pentosaceus B49, which may be attributed to the microbial methylation of purine compounds present in bamboo shoots. Gentisyl alcohol, a compound exhibiting radical-scavenging activity [28], was observed to be upregulated during the 24 to 48 h BSBL fermentation process mediated by P. pentosaceus B49. Similarly, 6-hydroxymelatonin, a metabolite of melatonin that also displays antioxidant activity [29], was continuously upregulated throughout the same fermentation period mediated by P. pentosaceus B49. Three distinct lipid classes including phosphatidylglyceride (PG) (16:0/16:0) and PG (18:1/18:1) along with triglycerides (TG) (24:2) are supposed to be the lipid components present in bamboo shoots, with their content exhibiting significant changes during the 24 to 48 h BSBL fermentation process mediated through P. pentosaceus B49. Phaseollin and vanylglycol, both classified as phenylpropanoid compounds, demonstrate distinct biological activities. Phaseollin acts as an antifungal phytoalexin [30], while vanylglycol may exhibit an anti-inflammatory property [31]. Both compounds experienced marked increases in signal intensity from 24 to 48 h in B49-fermented BSBL. N-acetylanthranilate and spermine are likely metabolic derivatives of amino acids in bamboo shoots generated by the P. pentosaceus B49 strain, exhibiting significant signal attenuation during the 24–48 h fermentation phase.
By comparing the overall differences in log_2_FC values at various fermentation stages (BS24H vs. BS0H, BS48H vs. BS24H, BS72H vs. BS48H, BS96H vs. BS72H) as shown in Table 1 and Table 2, it becomes evident that the magnitude of changes in metabolite signal intensity in BSBL fermentation broth gradually decreases as the fermentation progresses. Among the detected metabolites, N-demethylvindolidine—an indole alkaloid which is produced by removing a methyl group from the nitrogen atom of the vindolidine molecule [32]—was identified for the first time in BSBL fermentation broth in this study, likely representing a demethylation product of vindolidine metabolized by the P. pentosaceus B49 strain [33]. 5-methylthioadenosine may be a metabolic product of S-adenosylmethionine processed by P. pentosaceus B49, and prior research has highlighted its potential anticancer properties [34]. Val-Ile-Val-Leu-Leu is likely an oligopeptide synthesized by the B49 strain using amino acids from BSBL. Purpuritenin B, a diterpenoid compound predominantly isolated from the seeds of Annona purpurea [35], was also detected in the BSBL fermentation broth. 13-oxo-octadecadienoic acid (13-OxoODE), an oxylipin derived from linoleic acid oxidation, plays a significant role in inflammation and immune regulation [36]. Although direct metabolism of 13-OxoODE by LAB has not been reported, some strains are known to modulate the transformation of unsaturated fatty acids like linoleic acid [37], which may indirectly affect oxylipin levels, including 13-OxoODE, under certain conditions. The signal intensity of alisol C monoacetate increased during the 24–48 h fermentation period but declined from 48 to 96 h, reflecting the dynamic transformation of triterpenoids in bamboo shoots by the P. pentosaceus B49 strain. Benzoylcholine, postulated as a flavor compound in fermented bamboo shoots, exhibited a significant reduction in signal intensity during the 48–72 h fermentation period by P. pentosaceus B49. The decrease in citramalate from 48 to 72 h may be associated with isoleucine synthesis via the threonine deaminase pathway, while the increase from 72 to 96 h could stem from the strain’s utilization of pyruvate and acetyl-CoA for biosynthesis. Manool, a diterpenoid abundant in sage, was detected in BSBL fermentation broth and has demonstrated selective antitumor activity against melanoma cells [38]. 3-hydroxy-3-methylglutaric acid, a derivative of glutaric acid, may be produced through the metabolism of carbohydrates and amino acids by the P. pentosaceus B49 strain, with its signal intensity rising between 72 and 96 h of fermentation. Neoxanthin, a carotenoid widely present in plants and algae [39], was detected in the BSBL fermentation broth, with increasing signal intensity between 72 and 96 h; previous studies have shown its effectiveness in relieving symptoms related to renal failure [40]. Neocembrene, a characteristic diterpene hydrocarbon of R. communis [41], showed increased signal intensity during the 72–96 h fermentation of BSBL by P. pentosaceus B49. Methylphosphatidylcholine is an essential component of cell membranes, contributing to the maintenance of structural stability, mediation of signal transduction, and regulation of membrane fluidity [42]. LAB can decompose or transform lipids present in bamboo shoots, leading to the synthesis or consumption of various phospholipids, including methylphosphatidylcholine. In this study, a significant reduction in the signal intensity of methylphosphatidylcholine (42:9) was observed in samples fermented with BSBL for 72 to 96 h. (13S)-13-hydroperoxy-9Z,11E-octadecadienoic acid, a lipoxygenase product of linoleic acid, exhibited decreased signal intensity from 72 to 96 h of fermentation. Naturally occurring carotenoids in bamboo shoots may be converted into derivatives such as 1-hydroxy-γ-carotene glucoside through microbial activity, and the decrease in its signal intensity between 72 and 96 h may be linked to β-glucosidase activity in the P. pentosaceus B49 strain. Notably, the signal intensity of 6-hydroxymelatonin continuously increased throughout the fermentation process, suggesting that the P. pentosaceus B49 strain may modulate the dynamic balance of melatonin and its metabolites via the tryptophan metabolic pathway.
Detailed information regarding metabolite identification is provided in the Supplementary Materials. The detailed information for the top 30 significant DMs identified in the comparisons of BS24H vs. BS0H, BS48H vs. BS24H, BS72H vs. BS48H, and BS96H vs. BS72H are presented in Table S1, Table S2, Table S3, and Table S4, respectively. The experimental fragment ions are shown in the identification charts for each DM, where the green line represents the DM and the red line represents the standard from the database.
3.4. Variations in Selected Phenylpropanoids Metabolites
LAB fermentation significantly modulates phenolic profiles and bioactivities in plant-based foods [43]. LAB hydrolyzes phenolic glycosides into bioactive aglycones, thereby enhancing free phenolic content and antioxidant capacity through enzymatic bioconversion [44,45,46]. For instance, Lu et al. reported that fermentation and enzymatic hydrolysis increased the total phenolic content of guava leaf tea and enhanced both antioxidant and α-glucosidase inhibitory activities [44]. Some studies have found that spontaneous fermentation increases the content of certain phenolic acids and flavonoids, thereby improving the antioxidant and anti-inflammatory activities of extracts derived from tomato, grape, and coffee residues [45]. Zhang et al. investigated the influence of Bacillus subtilis fermentation on the composition of phenolic compounds and antioxidant activity in cornmeal [46]. Their results indicated that the fermentation process significantly increased both total phenolic and total flavonoid contents. Therefore, LAB fermentation plays a crucial role in promoting the release of phenolic compounds and enhancing their bioactivities in plant-based food. Phenylpropanoids including flavonoids, coumarins, are important bioactive compounds, with numerous studies confirming their significant antioxidant, antidiabetic, and antiobesity potential [47]. Therefore, this study focused on examining the dynamic changes in selected phenylpropanoid and polyketide metabolites during the BSBL fermentation process, as illustrated in Figure 3.
As depicted in Figure 3, compared with the unfermented samples, the signal intensities of 5,7-dimethoxyflavone, cinnamic acid, 3,4-dihydro-2H-1-benzopyran-2-one, 6,8-dimethyl-4-hydroxycoumarin, and 2-hydroxycinnamic acid in the BSBL fermentation broth were significantly elevated (p < 0.05), and these metabolites generally exhibited an upward trend as fermentation time increased. Conversely, the signal intensity of (+)-gallocatechin decreased gradually during fermentation, which is likely associated with its metabolic utilization by the P. pentosaceus B49 strain.
3.5. Variations in Bitter Peptides
Preliminary experimental results indicate that B49 fermentation significantly reduces the bitterness of BSBL. Among the 20 common amino acids, those most commonly associated with bitterness are leucine, isoleucine, valine, phenylalanine, methionine, tryptophan, lysine, and threonine. Generally, peptides composed of these amino acids also exhibit a bitter taste [48]. In this study, twelve of the most bitter short peptides were identified from the BSBL fermentation sample, namely alanylisoleucine (Ala-Ile), isoleucylisoleucine (Ile-Ile), leucylglycine (Leu-Gly), leucylvaline (Leu-Val), lysylisoleucine (Lys-Ile), phenylalanylisoleucine (Phe-Ile), phenylalanylproline (Phe-Pro), serylisoleucine (Ser-Ile), threonylleucine (Thr-Leu), threonylphenylalanine (Thr-Phe), tryptophylisoleucine (Trp-Ile), and valylisoleucine (Val-Ile). Therefore, this research focused on analyzing the changes in bitter peptides in BSBL during P. pentosaceus B49 fermentation, with the relevant results presented in Figure 4. As shown in Figure 4, the signal intensities of bitter peptides in samples fermented for 24, 48, 72, and 96 h were significantly lower than those in the unfermented samples (all p < 0.05). Furthermore, there were no significant changes in these bitter peptides between 24 and 96 h of fermentation, indicating that the removal of bitter peptides by P. pentosaceus B49 primarily occurred within the first 24 h of fermentation.
3.6. Overall KEGG Pathway Enrichment Analysis
To elucidate the organism-specific metabolic dynamics during P. pentosaceus B49 fermentation of BSBL, KEGG pathway enrichment analysis was performed using the bacterial metabolic pathway database (Pediococcus pentosaceus KEGG organism code: ppe) as the primary reference framework, with supplemental mapping to plant pathways where relevant for bamboo-derived metabolites. This analysis specifically aimed to identify (1) metabolic pathways significantly altered by P. pentosaceus B49 activity across fermentation timepoints (0 h to 96 h), and (2) key bacterial biochemical transformations driving DM accumulation patterns. The DMs identified from pairwise comparisons of fermentation stages were annotated using KEGG Compound database and mapped to pathways via the KEGG Mapper search tool. Enrichment significance (Fisher’s exact test, p < 0.05) was calculated against the P. pentosaceus metabolic background.
As shown in Figure 5A, the most significantly enriched pathways included core bacterial metabolic processes: alanine, aspartate and glutamate metabolism (ko00250); tyrosine metabolism (ko00350); arginine and proline metabolism (ko00330); pentose phosphate pathway (ko00030); citrate cycle (TCA cycle, ko00020); and lysine biosynthesis (ko00300). While some KEGG pathway designations reference human diseases (e.g., ko05230, ko05033), these represent evolutionarily conserved metabolic modules that function in bacterial systems. Figure 5B quantifies DM associations, with tyrosine metabolism (11 DMs), arginine/proline metabolism (10 DMs), and central carbon metabolism (10 DMs) showing highest involvement. Integration of DM profiles with enriched pathways revealed a bacterial-centric metabolic network (Figure 5C), highlighting the key pathways driving fermentation dynamics, including amino acid metabolism and energy production (pentose phosphate pathway, TCA cycle). This systematic approach clarifies how P. pentosaceus B49 reprograms both microbial and plant-derived metabolism during bamboo shoot fermentation.
4. Conclusions
This research explored metabolic shifts in BSBL during fermentation with P. pentosaceus B49. PLS-DA analyses in ESI+ and ESI− modes demonstrated significant variations in metabolite profiles at distinct time intervals (0 h, 24 h, 48 h, 72 h, 96 h). The most notable changes arose within the first 24 h, whereas subsequent modifications from 24 h to 96 h revealed a trend toward stabilization. Relative to unfermented controls, concentrations of compounds like 5,7-dimethoxyflavone, cinnamic acid, 3,4-dihydro-2H-1-benzopyran-2-one, 6,8-dimethyl-4-hydroxycoumarin, and 2-hydroxycinnamic acid were substantially higher (p < 0.05), with these metabolites generally rising as fermentation progressed. In contrast, (+)-gallocatechin levels steadily diminished. However, bitter peptides such as alanylisoleucine, isoleucylisoleucine, leucylglycine, leucylvaline, lysylisoleucine, and phenylalanylisoleucine registered significantly reduced abundances in fermented samples at 24 h, 48 h, 72 h, and 96 h compared to pre-fermentation states (all p < 0.05). KEGG pathway enrichment analysis identified tyrosine metabolism (ko00350) and arginine-proline metabolism (ko00330) as linked to the highest counts of DM species. These results deepen insights into metabolic control during BSBL fermentation, establishing a theoretical basis for functional bamboo shoot product development.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhang Y. Wu L. Li Y. Yang J. Yang H. Zhao Y. Chen G. Bamboo shoot and its food applications in last decade: An undervalued edible resource from forest to feed future people Trends. Food Sci. Technol.202414610439910.1016/j.tifs.2024.104399 · doi ↗
- 2Jana U.K. Bhardwaj P.K. Jeyaram K. Shukla J.K. Somkuwar B.G. Mukherjee P.K. Bamboo shoots: Comprehensive perspectives on food composition, nutritional value, and therapeutic potential J. Food Compos. Anal.202514010719810.1016/j.jfca.2025.107198 · doi ↗
- 3NSBC Database National Bureau of Statistics of China Available online: http://www.stats.gov.cn/(accessed on 14 January 2021)
- 4Ma T. Mo W. Lv B. Wang W. He H. Jian C. Liu X. Li S. Guo Y. A Review of the Nutritional Composition, Storage Challenges, Processing Technology and Widespread Use of Bamboo Shoots Foods 202413353910.3390/foods 1322353939593955 PMC 11592693 · doi ↗ · pubmed ↗
- 5Lin Z. Chen J. Zhang J. Brooks M.S. Potential for Value-Added Utilization of Bamboo Shoot Processing Waste—Recommendations for a Biorefinery Approach Food Bioproc. Tech.20181190191210.1007/s 11947-018-2088-3 · doi ↗
- 6Liu L. Liu L. Lu B. Chen M. Zhang Y. Evaluation of Bamboo Shoot Peptide Preparation with Angiotensin Converting Enzyme Inhibitory and Antioxidant Abilities from Byproducts of Canned Bamboo Shoots J. Agric. Food Chem.2013615526553310.1021/jf 305064 h 23647018 · doi ↗ · pubmed ↗
- 7Escobar-Sánchez M. Carrasco-Navarro U. Juárez-Castelán C. Lozano-Aguirre Beltrán L. Pérez-Chabela M.L. Ponce-Alquicira E. Probiotic Properties and Proteomic Analysis of Pediococcus pentosaceus 1101 Foods 2023124610.3390/foods 1201004636613263 PMC 9818561 · doi ↗ · pubmed ↗
- 8Meng C. Wang Y. Xie J. Xuan J. Geng J. Liu G. Tu J. Xiao H. Pediococcus pentosaceus JS 35 improved flavor, metabolic profile of fermentation supernatant of mulberry leaf powder and increased its antioxidant capacity Front. Nutr.202512155168910.3389/fnut.2025.155168940104817 PMC 11913688 · doi ↗ · pubmed ↗