Investigating the Influence of Conventional vs. Ultra-High Dose Rate Proton Irradiation Under Normoxic or Hypoxic Conditions on Multiple Developmental Endpoints in Zebrafish Embryos
Alessia Faggian, Gaia Pucci, Enrico Verroi, Alberto Fasolini, Stefano Lorentini, Sara Citter, Maria Caterina Mione, Marco Calvaruso, Giorgio Russo, Emanuele Scifoni, Giusi Irma Forte, Francesco Tommasino, Alessandra Bisio

TL;DR
This study examines how ultra-high dose rate proton irradiation affects zebrafish embryos under different oxygen conditions, finding that it can reduce toxicity in some cases.
Contribution
The study demonstrates the FLASH effect in zebrafish embryos under both normoxic and hypoxic conditions, highlighting tissue-specific responses.
Findings
Ultra-high dose rate proton irradiation reduces toxicity in zebrafish embryos compared to conventional dose rates.
The protective effect of ultra-high dose rate irradiation is observed in pericardial edema under normoxic conditions and in head and eye size under hypoxic conditions.
The FLASH effect appears to be tissue- and endpoint-dependent, with oxygenation playing a variable role.
Abstract
FLASH radiotherapy is based on the selective sparing of normal tissue side effects, keeping the same anti-tumor effectiveness, when the irradiation is performed at an ultra-high dose rate (i.e., >40 Gy/s). In this study, we explore the response of zebrafish embryos after proton irradiation at conventional and ultra-high dose rates, in terms of multiple developmental endpoints. The role of oxygenation at the time of irradiation is also explored. Overall, our data indicate that the so-called FLASH effect can be observed either under normoxic and hypoxic conditions, being more or less pronounced depending on the specific tissue and endpoint considered. This suggests that the sparing effect can have a significant dependence on the specific tissue, and that oxygenation could play a different role depending on the tissue type. Objectives: To investigate how the FLASH effect modulates…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
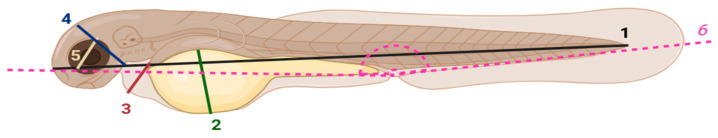 Figure 1
Figure 1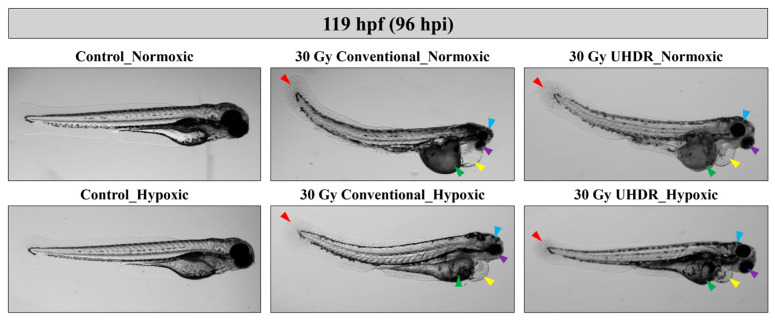 Figure 2
Figure 2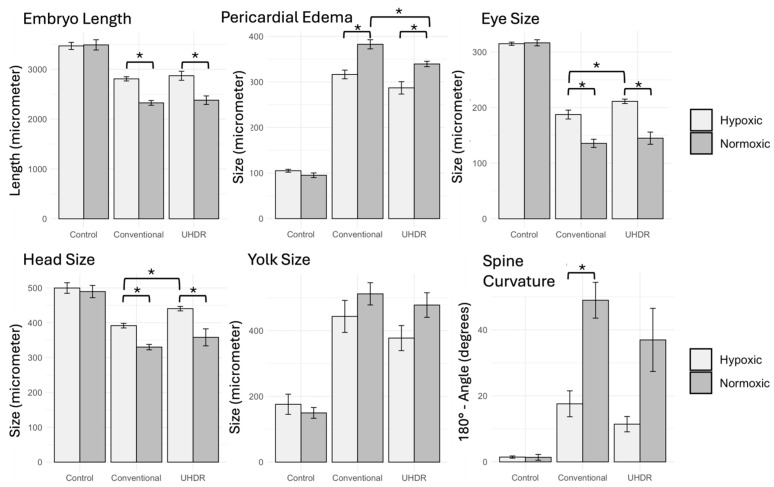 Figure 3
Figure 3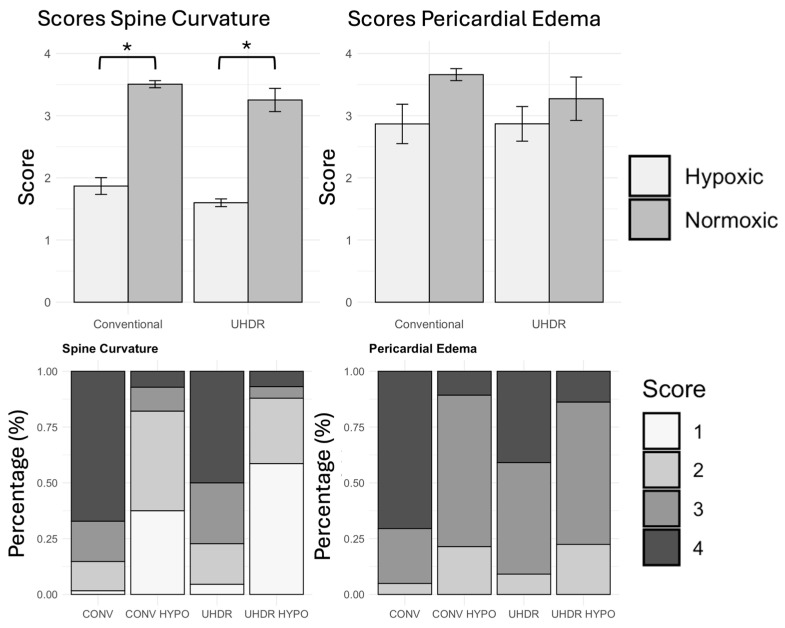 Figure 4
Figure 4- —GIOCONDA (Understanding the radiobiological effects of BNCT and FLASH therapy) project
- —ASTROFLASH (Assessing Toxicity and Effectiveness of Proton and FLASH Therapy in Zebrafish Models of Brain Cancer) project
- —European Union-NextGenerationEU
- —INFN-CSN5: FRIDA project
- —European Union—Next Generation EU
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRadiation Therapy and Dosimetry · Effects of Radiation Exposure · Advanced Radiotherapy Techniques
1. Introduction
In recent years, a new scenario has been opened in the field of radiotherapy (RT), related to the so-called FLASH effect. This consists of the reduction in normal tissue toxicities while maintaining high anti-tumor efficiency, which arises when a radiation dose is delivered in a very short time (i.e., at an ultra-high dose rate, UHDR) compared to conventional treatments. After the seminal paper by Favaudon and colleagues appeared in 2014 [1], an obvious interest was registered in the RT community. When translated to the clinics, the FLASH effect might provide a unique tool to enlarge the therapeutic window, namely the dosimetric distance between tumor control and normal tissue complication probabilities. The practical implications of this result were soon acknowledged, and extensive research started.
However, all that glitters is not gold. Almost in parallel with the enthusiasm raised around the experimental results, the difficulty to grasp the exact conditions under which the FLASH effect appears and could then be exploited emerged. While independent research groups [2,3], employing different animal models and irradiation approaches, reported results consistent with those obtained by Favaudon et al., there were also a significant number of negative reports, challenging the promising potential of the UHDR.
After about ten years of research on the topic, some results can be considered overall established. First, experimental studies indicate that quite specific irradiation conditions trigger the FLASH effect. Quasi-threshold values have been identified for the dose and dose rates, which are required to be above approximately 8 Gy and 40 Gy/s (i.e., the UHDR), respectively. These values are higher than the 1.8–2 Gy delivered in typical RT treatment at a dose rate of ≅ 0.1 Gy/s. Although studies show that temporal aspects on a smaller scale, associated with the delivery of dose pulses at the micro- or millisecond level, could also play a role, the reported values are typically considered a reference.
From a practical point of view, this implies that the delivery of UHDR treatments would require technical aspects to be taken into account [4]; in this context, clinical protons accelerated with a cyclotron are considered the most promising tool for the near future [5].
Several explanations have been proposed to elucidate the FLASH mechanisms [6,7], but none has been able to provide a comprehensive and unique picture so far. While the initial hypotheses largely relied on a transient induced “local” hypoxia due to the oxygen depletion because of the high dose and dose rates [8], this theory called transient oxygen depletion (TOD) has been largely shown as inconsistent [9,10,11]. It is now indeed established that the origin of the FLASH effect has to be searched in a complex network of biochemical processes taking place at the cellular level during and shortly after irradiation, with oxygenation still playing a role albeit not in the simple way initially suggested, and pointing also to a multiplicity of factors, including reactive oxygen species (ROS), their recombination, iron, and lipid metabolism, which, taken together, might contribute to explaining not only the sparing effect of healthy tissues but also the concurrent iso-effectiveness of the treatment on cancer cells [7,11,12,13,14].
In this study, we used zebrafish embryos to investigate the effects of UHDR proton irradiation at two different oxygenation levels on the development process. Zebrafish share approximately 70% of human genes, and over 80% of known human disease-related genes—including oncogenes and tumor suppressors—have at least one ortholog. Combined with their high throughput and relative ease of handling, zebrafish serve as an effective intermediate model for studying biological responses [15]. This is also the case of the investigation for the mechanistic bases of the FLASH effect, being a purely in vivo effect, but presenting some features that are emerging and can be partially explorable also with in vitro assays [16]. Zebrafish, representing the ideal bridge between the two approaches, were thus previously employed extensively for the study of the FLASH effect [17,18,19,20,21,22,23,24,25]. Different endpoints and irradiation conditions were explored, and contradictory results have been reported so far. While some studies showed a significant FLASH effect on the malformation rate of multiple organs, other works did not observe significant differences, or they did so only for some specific tissues.
One important aspect that we explored is the interplay of the protective effect induced by the dose rate and the level of oxygenation. In fact, while, as above mentioned, the specific impact of oxygen in the FLASH mechanism is far from being understood, beside the disproof of the TOD, significant results reported an increased emergence of the FLASH effect at lower oxygenation conditions, with mice novel object recognition tests [17]. Such an effect was confirmed also by zebrafish experiments, with a larger occurrence in hypoxic conditions [18,19,21]. At the same time, it appeared that reactive oxygen species and radical recombination may be importantly correlated by the observed sparing effect versus dose rate [9].
We present in this work an analysis of developmental and morphological endpoints such as larval length, yolk malabsorption, pericardial edema, head and eye size, and spinal curvature, which we employed to assess the extent of normal tissue sparing under UHDR conditions. The role of hypoxia was also investigated. This approach aims to confirm and extend previous findings in zebrafish while addressing gaps in our understanding of dose-rate and oxygenation dependencies in FLASH RT.
2. Materials and Methods
2.1. Zebrafish Care and Mating
Adult zebrafish (Danio rerio) wild-type AB-strain were housed at the Model Organism Facility, Department of Cellular, Computational and Integrative Biology (CIBIO Department, University of Trento, Italy) and they were kept under standard conditions (14 h light/10 h dark cycle, temperature 27.5 °C, conductivity 500 ± 50 μS, and pH 7.5).
All experiments involving zebrafish were conducted in compliance with the European and Italian regulations (D.Lgs. 26/2014). No ethical approval for the use of fish was required since larvae were used before the fifth day post-fertilization. Adult fish used to generate embryos belonged to the colony of MOF authorized as breeders by the Municipality of Trento (C_L378/S022/103359/03.06.2015).
2.2. Preparation of Zebrafish Samples for Treatments
Three independent experimental replications were performed on different days using embryos from different breeding couples of the same line. The number of embryos for each replicate of the same experimental condition was variable in the range 23–27.
To place zebrafish embryos for irradiation, we used 0.5 mL Eppendorf tubes as support. According to a previously proposed protocol [21], we melted agarose to a concentration of 0.8–1%, and we carefully pipetted it into the conical bottom part of the Eppendorf tubes. Next, once the agarose had cooled and solidified, we gently placed 23–27 embryos at 24 h post-fertilization (hpf) onto the solid agarose layer and distributed them uniformly. The embryos occupied a final thickness of about 1.5 mm. Finally, the entire Eppendorf tube was filled with E3 medium. No methylene blue was used in the E3 medium to avoid interfering with redox reactions induced by irradiation.
Half of the prepared samples were irradiated under normoxic and half under hypoxic conditions. For the hypoxic group, the Eppendorf tubes with zebrafish embryos were placed with open caps inside a controlled hypoxic chamber (Ruskinn^®^ INVIVO2 200, Ruskinn Technology Ltd., Bridgend, UK). The chamber was adjusted to establish and maintain a low-oxygen environment (O_2_: 1%, CO_2_: 0%, T: 28 °C). Embryos were kept in the chamber for 90 min to achieve a stable hypoxic state. Tubes were closed to prevent reoxygenation before irradiation, and lids were sealed with vacuum grease and Teflon.
As verification, and to simulate the real timing of the irradiation experiment, oxygen measurements were taken at specific time points (i.e., 60, 90, and 120 min), after removal from the hypoxia chamber and sealing of the Eppendorf tubes, using the PreSens PM-PSt8 needle of PreSens Precision Sensing system (Regensburg, Germany) (Schematic representation and clos-up photo of the system in Supplement S1). Overall, after incubation in the hypoxic chamber, the measured oxygen level was equal to 1.3% ± 0.5% (range 0.8–1.8%) in the monitored time interval.
2.3. Monitoring of Zebrafish Development
After irradiation, embryos were transferred to 96-well plates, with one embryo per well, to monitor the development individually over time. The plates were maintained at a constant temperature of 28 °C and monitored daily, at consistent time points. Pictures of all embryos were acquired using a Leica^®^ MZ16 stereomicroscope with bright light illumination. Quantitative analyses of developmental malformations were conducted twice by two independent operators, by assessing the following features:
- –Larval length [μm]
- –Yolk absorption [μm]
- –Pericardial edema [μm]
- –Head size [μm]
- –Eye size [μm]
- –Spinal curvature [(180—angle)°]
Measurements were performed manually at 119 hpf using ImageJ software (ImageJ-1.54f (64bit version)), after appropriate calibration, as showed in Figure 1.
A morphological scoring system, originally developed by Brannen et al. [26] and described by Szabò et al. [27], was also adopted for the analysis of two specific endpoints, namely pericardial edema and spine curvature. Images of individual zebrafish larvae at the same time point (119 hpf) were retrospectively assessed, and visible malformations were classified according to the scoring system, as described in Supplement S2.
2.4. Irradiation and Dosimetry
Zebrafish embryo irradiation was performed at the Trento proton beam line (TPBL), which is part of the local proton therapy center (Azienda Provinciale per i Servizi Sanitari, APSS, Trento, Italy). The irradiation facility was extensively described in the past [28,29]; recently it introduced the possibility of performing UHDR proton irradiation. To this purpose, new beam transport optics were tuned for the highest available proton beam energy of 228 MeV, for which the highest transport efficiency can be reached. The proton beam at the irradiation position was then characterized in terms of lateral profile, intensity, and stability. Details on the beam characteristics are reported in Supplement S3 (Figure S2).
The embryos were irradiated in 0.5 mL Eppendorf tubes, upon alignment at irradiation position by means of dedicated lasers. Each tube received an expected dose of 30 Gy, delivered either at a conventional dose rate (0.6 Gy/s) or UHDR (317 Gy/s). The dose delivery was monitored online based on the counting obtained from a previously calibrated integral ionization chamber. Additionally, an EBT-XD gafchromic film (Ashland Inc., Wilmington, DE, US) was irradiated together with each sample and the delivered dose was then reconstructed offline.
2.5. Statistical Analysis
Concerning radiation-induced malformations, each data point in the plots represents the inverse-variance weighted average calculated for the three replicates, together with the associated standard deviation. Similarly, a mean score value was calculated for each replicate for both endpoints (i.e., pericardial edema and spine curvature). An average value was then computed accounting for the three replicates, together with the standard deviation. Statistical significance was evaluated by means of Student’s t-test. Statistical significance was attributed to p-values < 0.05.
3. Results
Zebrafish survival rate was monitored until 119 hpf, corresponding to about 96 h post irradiation, and no differences were found between control and irradiated samples. The morphological malformations observed on zebrafish larvae are displayed in Figure 2 for representative cases, according to the different treatment conditions. A quantitative analysis of the different endpoints is reported in Figure 3. As expected, data show a radioprotective effect associated with the hypoxic irradiation environment compared to the normoxic one. This is consistently observed for each endpoint both for zebrafish irradiated at conventional and UHDR, which is associated in most cases with statistical significance. In terms of the dose rate parameter, there is a trend of slightly reduced toxicity after exposure to UHDR, which is maintained under both normoxic and hypoxic conditions. However, the difference turns out to be significant for a limited number of cases, i.e., normoxic irradiation for pericardial edema (382 ± 10 μm conventional vs. 339 ± 6 μm UHDR, p = 0.01), hypoxic irradiation for eye (187 ± 8 μm conventional vs. 211 ± 4 μm UHDR, p = 0.03) and head size (391 ± 6 μm conventional vs. 440 ± 6 μm UHDR, p = 0.001).
In terms of relative variation, the pericardial edema is reduced by about 11% at UHDR, while the measured eye and head size are larger by about 13% and 12%, respectively. Notably, for pericardial edema the difference between conventional and UHDR irradiation approaches does not reach significance also for the hypoxic case (Supplement S3 Table S2 for p-values). Control groups under normoxic and hypoxic conditions show comparable developmental outcomes, indicating that the temporary hypoxic incubation does not adversely affect zebrafish development.
Similar results emerge from the analysis of the average toxicity score (i.e., SV mean) for pericardial edema and spinal curvature (Figure 4). A differential response is observed for embryos irradiated under hypoxic or normoxic conditions; however, significant differences are only attributed to spinal curvature. The possible protective role of UHDR here translates into slightly lower average scores attributed to UHDR samples, which nevertheless do not result in statistically significant deviations from conventional ones. When comparing conventional and UHDR, we observe for spinal curvature an average score reduction of approximately 7% and 15% for normoxic and hypoxic irradiation, respectively; for pericardial edema, the average score is comparable for the hypoxic samples, while a reduction of about 10% is associated with UHDR under normoxia.
These results are complemented by the scoring percentage distribution, summarized in Figure 4. On top of the obvious dependence on the oxygenation level, these data show a tendency toward a protective effect of UHDR, which is less evident when looking at the average scores (Figure 4). UHDR appears to be associated with a lower percentage of embryos in the high score categories. This is observed both for pericardial edema and spinal curvature, as well as under normoxic and hypoxic conditions, with the only exception registered for the spinal curvature endpoint for hypoxic irradiation. (Supplement S3 Table S3 for p-values).
4. Discussion
An increasing interest in Zebrafish as an experimental model for radiobiology research has been registered in recent years. This is confirmed by several publications focused on investigating the FLASH effect [17,18,19,20,21,22,30,31], but also by other studies unrelated to UHDR and dedicated to comparing the biological effects of different radiation qualities [26,27]. Although zebrafish possess inherent limitations compared to mammalian models—most notably in terms of tissue complexity and immune system maturation—they offer significant advantages in scalability and experimental throughput. Their optical transparency, external development, and rapid embryogenesis allow for high-resolution phenotypic assessment and facilitate high-content screening approaches that are essential for dose–response characterization and multivariate analyses.
We presented the results of a comprehensive study of zebrafish developmental malformations induced by conventional and UHDR irradiation, comparing the results for normoxic and hypoxic conditions. Our study contributes to the ongoing research dedicated to the understanding of the FLASH effect, a context in which zebrafish have been employed obtaining partially contradictory results. Montay-Gruel et al. [17] showed a protective effect of UHDR electrons compared to a conventional dose rate after 8 Gy electron irradiation. Interestingly, when the samples were irradiated in the presence of antioxidants (amifostine or NAC), a sparing effect was observed after conventional exposure but not after UHDR. Beyreuther et al. were the first to report experimental outcomes on embryos irradiated with protons at different dose rates [18]. While those results showed a FLASH effect limited to a single end-point (i.e., pericardial edema), a later publication employing irradiation at controlled oxygenation (i.e., pO_2_ < 10 mmHg, obtained by sealing zebrafish embryos inside the Eppendorf tube and thus consuming oxygen without replacement) indicated a significant radioprotection at a UHDR for embryo length, spine curvature, and pericardial edema, both after electron and proton irradiation [21]. Consequently, a protocol for embryo preparation was proposed to allow irradiation under hypoxia, which we adapted to our experiments that include incubation inside a hypoxic chamber. Interestingly, the authors reported an increase in length of about 3% at a UHDR compared to conventional, which is in line with the about 2.2% increase observed in our study. Such results were further corroborated by the dose response curves published by the same group [19], employing protons at the entrance and mid-SOBP (i.e., spread-out Bragg peak). In contrast, Kacem et al. [22] reported negative results in terms of a missing radioprotective effect after either x-ray, electron or proton irradiation; remarkably, oxygenation levels were not monitored during those experiments, leaving room for the possible influence of the hypoxic environment on the FLASH effect induction. In Jansen et al. 2022 [9], oxygen depletion was measured in the same conditions of the zebrafish irradiations, and it was taken as an alias of the radical recombination rate. Larger dose rates correlated with a lower depletion, a value confirmed by several following studies, thus indicating, in addition to another point of inconsistency for the TOD hypothesis, a possible increase in radical recombination at a UHDR as a source of the protective effect. Additional results were obtained by Saade et al. [25], who adopted the protocol proposed in [21] and therefore irradiated zebrafish embryos at comparable oxygen levels. The authors showed a sparing effect for embryo length and pericardial edema, while no significant effect was associated with spinal curvature. In this case, embryo length was about 6% larger at 30 Gy UHDR compared to conventional, an effect somewhat larger than the 2.2% increase that we reported. Interestingly, the FLASH effect was more pronounced at 30 Gy than at 40 Gy, suggesting a threshold or saturation phenomenon in the biological response. In a separate publication, malformations were observed after helium ion irradiation, obtaining results overall in line with the previous study [23]. The same group recently published partially contradictory results [24], comparing proton irradiation at conventional and extremely high dose rates. In this case, no sparing was observed on morphological endpoints. However, they highlighted a higher level of apoptosis at conventional irradiation compared to UHDR, supported by higher levels of DNA fragmentation detected by the TUNEL test and higher expression levels of genes controlling cell cycle and p53-dependent apoptosis.
Our findings fit in this scenario, as samples subjected to UDHR-hypoxic conditions showed a better sparing of all tissues evaluated for toxic effects, despite not always being supported by statistical significance. Remarkably, the protective effect is observed both for normoxic (pericardial edema) and hypoxic (head and eye size) conditions. Taken together, these results suggest that while UHDR may reduce radiation-induced damage, its protective effects are endpoint-dependent. The role of oxygenation might as well be pronounced to a different extent depending on the tissue involved. The different degrees of response observed for different tissues agree with the recent report by Bogaerts et al. Combining gene expression analysis with immunohistochemistry, they were able to show that, for instance, apoptosis is not uniformly activated but is more pronounced in the tail compared to the head region. This paves the way for further studies exploiting molecular biology techniques to enhance our understanding of the differential response associated with the FLASH effect, possibly including information on oxygenation levels.
In this context, a recent modeling work of our group [14] suggested a tissue-dependent antioxidant rate as the main driver of the differential sparing effect between normal tissue and tumor, through the process of organic radical recombination. Thus, an immediate extension of the present work providing further insights would be the inclusion of specific antioxidant molecules to check how they could modulate the occurrence of the sparing in hypoxic and normoxic conditions.
In summary the obtained results evidence a dose rate-dependent protective effect which appears only at specific endpoints, thus highlighting a potential role of the tissue (chemical) environment for its manifestation. Moreover, we observe the effect at any oxygen concentration, thus pointing out the consistency of FLASH sparing also in normoxic conditions, differently from other works but similarly to what happens, e.g., for in vitro irradiations [32]. Finally, the extent of the UHDR-induced sparing is notably smaller as compared to the hypoxia-induced one, the latter being also present in all the analyzed endpoints, thus remarking the large difference in the two effects, and providing additional, indirect support to the implausibility of the RTOD hypothesis.
5. Conclusions
Our study indicates that, while the UHDR confirms its potential to reduce radiation-induced damage, these protective effects are importantly endpoint-dependent. While the pure effect of hypoxia-induced radioprotection appears instead ubiquitous and larger in size, in the combination of the two the relative role of oxygenation in the UHDR might as well be dependent on the tissue involved. Further developments of the present work will include gene expression studies, aiming at a better understanding of the pathways activated by the different irradiation regimes; the role of radioprotectors, and their interplay with dose rate and oxygenation, will be investigated as well, to shed additional light on the channels associated with UHDR-induced damage reduction.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Favaudon V. Caplier L. Monceau V. Pouzoulet F. Sayarath M. Fouillade C. Poupon M.F. Brito I. HupéP. Bourhis J. Ultrahigh Dose-Rate FLASH Irradiation Increases the Differential Response between Normal and Tumor Tissue in Mice Sci. Transl. Med.20146245 ra 9310.1126/scitranslmed.300897325031268 · doi ↗ · pubmed ↗
- 2Vozenin M.C. Bourhis J. Durante M. Towards Clinical Translation of FLASH Radiotherapy Nat. Rev. Clin. Oncol.20221979180310.1038/s 41571-022-00697-z 36303024 · doi ↗ · pubmed ↗
- 3Esplen N. Mendonca M.S. Bazalova-Carter M. Physics and Biology of Ultrahigh Dose-Rate (FLASH) Radiotherapy: A Topical Review Phys. Med. Biol.20206523 TR 0310.1088/1361-6560/abaa 2832721941 · doi ↗ · pubmed ↗
- 4Zou W. Zhang R. Schüler E. Taylor P.A. Mascia A.E. Diffenderfer E.S. Zhao T. Ayan A.S. Sharma M. Yu S.-J. Framework for Quality Assurance of Ultrahigh Dose Rate Clinical Trials Investigating FLASH Effects and Current Technology Gaps Int. J. Radiat. Oncol. Biol. Phys.20231161202121710.1016/j.ijrobp.2023.04.01837121362 PMC 10526970 · doi ↗ · pubmed ↗
- 5Schwarz M. Traneus E. Safai S. Kolano A. van de Water S. Treatment Planning for Flash Radiotherapy: General Aspects and Applications to Proton Beams Med. Phys.2022492861287410.1002/mp.1557935213040 · doi ↗ · pubmed ↗
- 6Hall E.J. Radiation Dose-Rate: A Factor of Importance in Radiobiology and Radiotherapy Br. J. Radiol.197245819710.1259/0007-1285-45-530-814622835 · doi ↗ · pubmed ↗
- 7Limoli C.L. Vozenin M.-C. Reinventing Radiobiology in the Light of FLASH Radiotherapy Annu. Rev. Cancer Biol.2023712110.1146/annurev-cancerbio-061421-02221739421564 PMC 11486513 · doi ↗ · pubmed ↗
- 8Weiss H. Epp E.R. Heslin J.M. Ling C.C. Santomasso A. Oxygen Depletion in Cells Irradiated at Ultra-High Dose-Rates and at Conventional Dose-Rates Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med.197426172910.1080/095530074145509014607987 · doi ↗ · pubmed ↗