Evaluation of Metabolic Characteristics Induced by Deoxynivalenol in 3D4/21 Cells
Yu Han, Bo Yu, Wenao Weng, Liangyu Shi, Jing Zhang

TL;DR
This study explores how the mycotoxin deoxynivalenol affects metabolism in pig immune cells, identifying key metabolic changes that could help reduce its harmful effects.
Contribution
The study reveals novel metabolic pathways affected by deoxynivalenol in porcine macrophages, linking them to immunotoxicity through integrated metabolomic and transcriptomic analysis.
Findings
DON reduced cell viability in a concentration- and time-dependent manner in 3D4/21 cells.
127 differential metabolites were identified, primarily affecting purine, glutathione, and arginine–proline metabolism.
Metabolic changes were confirmed to play key roles in DON-induced immunotoxicity through integration with transcriptomic data.
Abstract
Deoxynivalenol (DON) is a common mycotoxin that weakens immune function in pigs. However, its effects on cellular metabolism remain unclear. This study employs porcine alveolar macrophages (3D4/21 cells) to investigate DON-induced metabolic alterations using non-targeted metabolomics. MTT assays showed DON reduced cell viability in a concentration- and time-dependent manner. Metabolomic analysis identified 127 differential metabolites, revealing distinct metabolic profiles between control and DON-treated cells. These changes mainly affected purine metabolism, glutathione metabolism, and arginine–proline metabolism. Integration with transcriptomics data confirmed these pathways are important for DON-induced immunotoxicity. The study provides new insights into DON-induced metabolic reprogramming in immune cells and identifies candidate targets for alleviating mycotoxin-driven…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
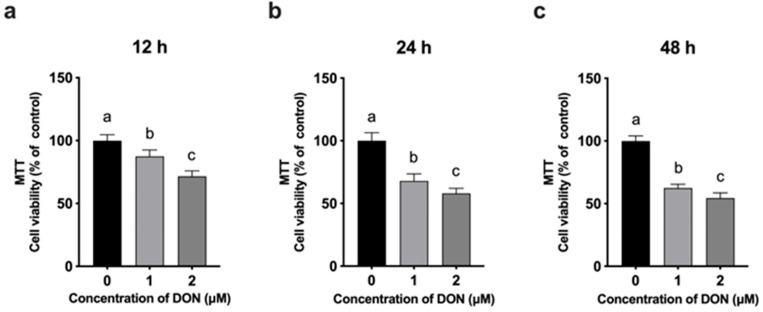 Figure 1
Figure 1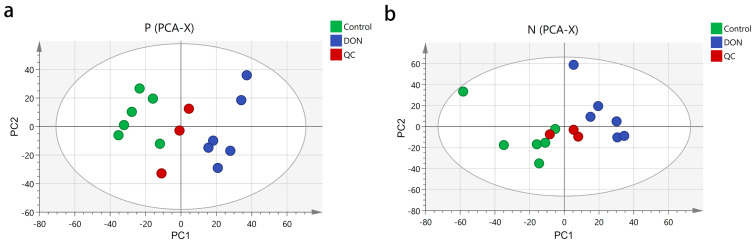 Figure 2
Figure 2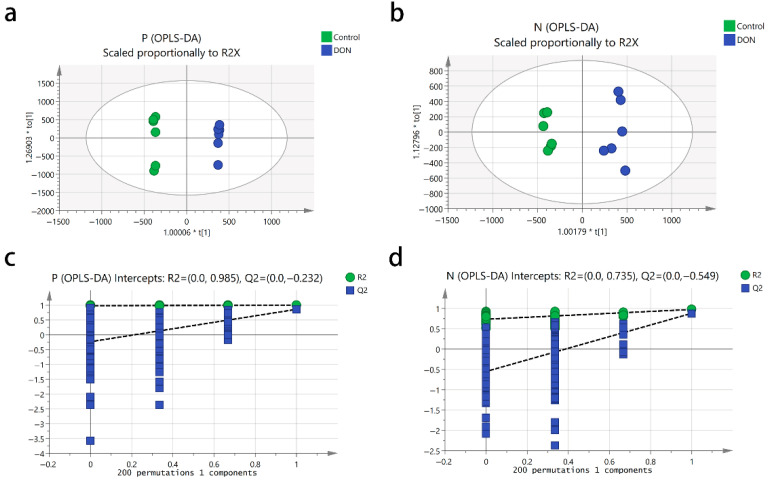 Figure 3
Figure 3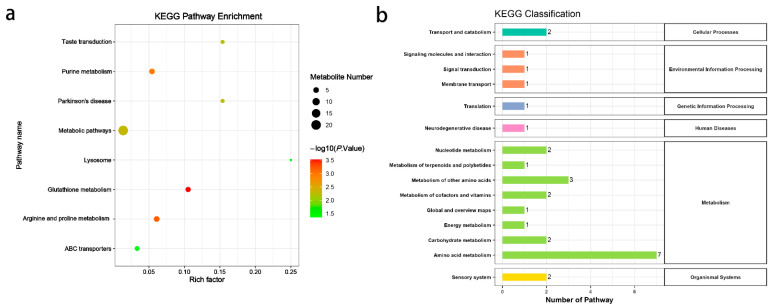 Figure 4
Figure 4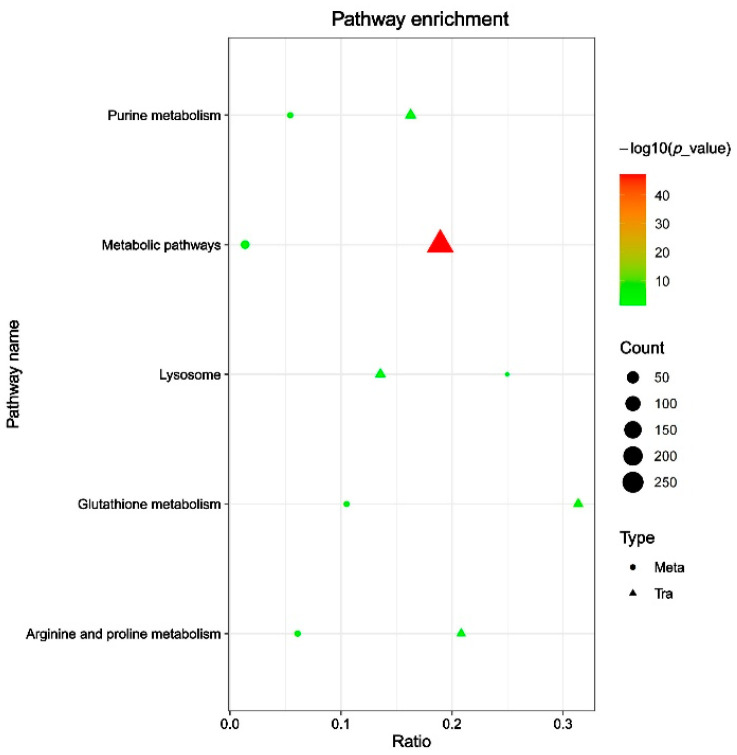 Figure 5
Figure 5- —the National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFood Quality and Safety Studies · Redox biology and oxidative stress · Mycotoxins in Agriculture and Food
1. Introduction
Mycotoxin contamination in feed is a major threat to global livestock health and causes severe economic losses in agriculture [1]. Among these toxins, deoxynivalenol (DON), a type B trichothecene produced by Fusarium species, is one of the most widespread contaminants in cereal crops worldwide [2]. Due to its high stability during storage and food processing, DON exposure through contaminated feed is almost unavoidable in the diet of domestic animals [3]. Even at subclinical levels, DON exposure reduces feed intake, slows growth, and decreases feed conversion efficiency, further harming productivity [4].
Pigs are especially susceptible to DON toxicity owing to their substantial dietary consumption of DON-contaminated cereals (wheat, corn, and barley) [5]. Compared with other domestic animals, pigs have a higher absorption rate of DON (exceeding 70%), mainly due to the short transit time of feed through the gastrointestinal tract and the efficient absorption of DON in the upper small intestine [4,6]. Moreover, only a small amount of DON can be converted to the less toxic metabolite DOM-1, resulting in high bioavailability of the toxin in the pig’s bloodstream [7]. Consequently, pigs are particularly susceptible to DON toxicity, even at low exposure levels. Previous research shows that even low DON levels (3.02 mg/kg) can reduce feed intake and growth in pigs [5], while in vitro studies report that DON as low as 0.8 μM impairs porcine immune cell function [8].
DON has strong effects on the porcine immune system in multiple ways. It directly damages immune organs such as the thymus and spleen, leading to atrophy of the thymic cortex, reduction in lymphocyte populations, and impaired development of germinal centers in the spleen [9,10,11]. In pig models, DON exposure leads to thymic atrophy and increased apoptosis of thymocytes, with significant reductions in cortical lymphocyte density [10,12]. In addition, DON has been reported to impair both T-cell and B-cell function in porcine peripheral blood mononuclear cells (PBMCs) [8]. However, the global metabolic alterations underlying DON-induced immunotoxicity remain poorly characterized.
Metabolomics, a powerful systems biology tool, provides a comprehensive view of how xenobiotic exposure affects cellular metabolism [13]. Non-targeted metabolomic approaches are particularly advantageous for elucidating unexpected pathways altered by toxic insults [14,15]. By mapping these metabolic alterations, researchers can pinpoint specific biomarkers for early detection of xenobiotic exposure, evaluate the severity of its toxic effects, and understand interindividual variability in susceptibility [13,16]. Nevertheless, limited data exist on DON-driven metabolic dysregulation in porcine immune cells, especially in the context of alveolar macrophage functionality.
Since alveolar macrophages play a key role in the first line of defense against pulmonary pathogens, the porcine alveolar macrophage cell line 3D4/21 has emerged as a valuable in vitro model to investigate DON-induced cellular and molecular alterations in pigs [17]. In this study, we employ an in vitro model using 3D4/21 porcine alveolar macrophages exposed to DON in combination with LC-MS/MS-based non-targeted metabolomics. Our work expands our understanding of DON’s immunometabolism toxicity and offers potential therapeutic targets for mitigating mycotoxin-related risks in swine.
2. Materials and Methods
2.1. Cell Culture
The 3D4/21 cells were obtained from the American Type Culture Collection and grown in RPMI 1640 medium (HyClone, Marlborough, MA, USA) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin–streptomycin solution (Gibco, New York, NY, USA). Cells were maintained at 37 °C with 5% CO_2_.
2.2. Cell Viability Assay
The 3D4/21 cells were seeded into 96-well plates at a density of 8 × 10^3^ cells/mL and treated with 0, 1, or 2 μM DON (Sigma-Aldrich, St. Louis, MO, USA) for 12, 24, or 48 h. Cell viability was assessed using the MTT assay (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; Beyotime, Shanghai, China) as previously described [17]. Briefly, 100 μL of fresh medium and 10 μL of freshly prepared MTT solution (5 mg/mL) were added to each well. After incubation, 100 μL of formazan solubilization solution was added to dissolve the crystals. The absorbance was measured at 570 nm using a microplate reader, and cell viability was calculated accordingly.
2.3. Metabolite Extraction
After treating 3D4/21 cells with or without DON (2 μM) for 24 h (six biological replicates per group), the cells were collected by centrifugation and stored in liquid nitrogen until further processing. The samples were first removed from liquid nitrogen, and 1 mL of −80 °C pre-cooled methanol was added to the sample containing 1 × 10^7^ cells. The mixture was vortexed for 30 s, then centrifuged at 4 °C and 13,000 rpm for 20 min. 1 mL of −80 °C pre-cooled HPLC-grade water was added to the remaining pellet, followed by one freeze–thaw cycle in liquid nitrogen, vortexed for 30 s, and centrifuged again (4 °C, 13,000 rpm, 20 min). Supernatants were pooled and centrifuged at 4 °C and 13,000 rpm for 20 min to ensure clarity. The clear supernatant was transferred to a fume hood and evaporated to dryness. The dried residue was reconstituted in 100 μL of initial mobile phase, sonicated for 5 min to ensure complete dissolution, and centrifuged at 4 °C and 13,000 rpm for 20 min prior to subsequent analysis.
2.4. Metabolite Identification and Data Analysis
Metabolite identification was performed using the LC-MS/MS by BIOMS Biotechnology (Beijing, China). Chromatographic separation was conducted on an ExionLC system (AB Sciex, Framingham, MA, USA) equipped with a Waters HSS T3 column (100 × 2.1 mm, 1.8 μM, Waters, Milford, MA, USA). Mass spectrometric analysis was carried out using a TripleTOF 5600 + system (AB Sciex) under both positive and negative ion modes with Information Dependent Acquisition (IDA) in high-sensitivity mode and dynamic background subtraction. In negative ion mode, mass spectrometry was performed with the following ion source parameters: sheath gas flow rate of 30 psi, Gas1 and Gas2 flow rates of 55 psi each, ion source temperature of 550 °C, and ion spray voltage of −4500 V. The data acquisition time was 14 min. The TOF MS scan range was set from m/z 100 to 1200, and each MS1 scan was followed by 12 product ion (MS/MS) scans. The MS/MS scan range was m/z 50~1200, with an accumulation time of 0.05 s for each MS/MS scan. The collision energy was set at −40 eV (spread ±20 eV). In positive ion mode, the same parameters were used except that the ion spray voltage was set at +5500 V and the collision energy was adjusted to +40 eV.
The raw data obtained from mass spectrometry detection were imported into Progenesis QI (v3.0) software for data preprocessing and metabolite identification. Metabolites were identified by referencing the Kyoto Encyclopedia of Genes and Genomes (KEGG, http://www.genome.jp/kegg/, accessed on 21 May 2024) and the Human Metabolome Database (HMDB, http://www.hmdb.ca, accessed on 21 May 2024). Quality control (QC) analysis was carried out to ensure the accuracy and reliability of the data. Metabolites with a coefficient of variation (CV) less than 30% in QC samples, a variable importance in the projection (VIP) value greater than 1 and a p-value less than 0.05 were selected as differential metabolites (DMs). To reveal the differences in metabolites among different components, principal component analysis (PCA) was carried out. To remove noise and identify metabolites that contribute most to group separation, orthogonal partial least squares discriminant analysis (OPLS-DA) was performed. The quality of the model was assessed by cross-validation, with R^2^Y and Q^2^ representing the explained variables and predictability of the model, respectively. Metabolic pathway enrichment was conducted using MetaboAnalyst (v5.0) (https://www.metaboanalyst.ca/, accessed on 27 May 2024) and KEGG (https://www.genome.jp/kegg/, accessed on 27 May 2024) to identify potentially dysregulated metabolic pathways.
2.5. Integrative Analysis of Metabolomics and Transcriptomics
Transcriptomic data were obtained from prior analysis of 3D4/21 cells treated with 2 μM DON for 24 h [17]. Differentially expressed genes (DEGs) filtered by log_2_ (fold change) ≥ 1 and FDR < 0.05. For metabolomics, 50 DMs with top VIP scores (VIP > 1) were selected, while 50 DEGs with the lowest FDR values were chosen for integration. Data were standardized (Z-score), and Pearson’s correlation coefficients (PCCs) between each DM-DEG pair were calculated in R (v4.2.1). Pairs with |PCC| ≥ 0.5 and p < 0.05 (adjusted by Benjamini–Hochberg) were considered significant. Pathway enrichment of significant pairs was performed via MetaboAnalyst. Correlation networks were visualized using Cytoscape (v3.9.1), with node sizes reflecting pathway enrichment and edge colors indicating positive/negative correlations.
2.6. Statistical Analysis
All experimental data are expressed as mean ± standard deviation (SD). Statistical significance was determined using one-way ANOVA followed by Tukey’s multiple comparisons test, with p < 0.05 considered significant. Different letters (e.g., a, b, and c) indicate statistically significant differences between groups.
3. Results
3.1. Viability of 3D4/21 Cells Following DON Exposure
The cytotoxic effects of DON on 3D4/21 cells were evaluated using an MTT assay after treatment with 0, 1, and 2 μM DON for 12, 24, and 48 h. As shown in Figure 1a, DON exposure for 12 h led to a significant, dose-dependent decrease in cell viability. A similar trend was observed at 24 and 48 h (Figure 1b,c), with higher DON concentrations causing progressively reduced viability compared to the untreated control.
A concentration of 2 μM DON for 24 h was chosen for metabolomic analysis, as it caused significant cytotoxicity without excessive cell death, enabling the detection of biologically relevant metabolic changes. Furthermore, this concentration and time point were consistent with those used in our previous studies [17], ensuring comparability across experiments.
3.2. PCA Principal Component Analysis
To investigate the metabolic characteristics of 3D4/21 cells after DON treatment, we conducted metabolomic profiling. Principal component analysis (PCA) showed distinct clustering patterns between the control and DON-treated groups, indicating significant metabolic perturbations induced by DON exposure. The peak areas of each metabolite in positive and negative ion modes were compared separately using SIMCA-P 14.1 software and subjected to principal component analysis. The three quality control (QC) samples were closely clustered near the center point in both the positive ion mode (Figure 2a) and the negative ion mode (Figure 2b). The samples of the same group were gathered in a relatively concentrated area and could be well distinguished from the other groups, confirming the reliability of the analytical method.
3.3. OPLS-DA Analysis and Iterative Validation
To filter out orthogonal signals and establish the OPLS-DA model, we analyzed the metabolic differences between the control and DON-treated groups. The OPLS-DA score plots demonstrated clear separation between the two groups and tight clustering within each group (Figure 3a,b). In positive ion mode, the model achieved an R^2^Y of 0.99 and a Q^2^ of 0.86 (Figure 3a), while in negative ion mode, the R^2^Y and Q^2^ values were 0.97 and 0.88, respectively (Figure 3b). The OPLS-DA model quality was then validated through 200 iterations of cross-validation (Figure 3c,d). Permutation test results showed regression line intercepts for predictive ability at −0.232 (positive ion mode) and −0.549 (negative ion mode), confirming a well-fitting model without overfitting.
3.4. Metabolic Pathway Analysis
We identified 127 DMs between the control and DON treatment groups, using thresholds of CV < 30%, VIP > 1, and p < 0.05 (Table 1). Among these, 36 metabolites were upregulated and 41 metabolites were downregulated in the positive ion mode (Table S1), while 19 metabolites were upregulated and 26 metabolites were downregulated in the negative ion mode (Table S2).
The heatmap of differential metabolites revealed clear clustering between control and DON-treated groups in both positive (Figure S1) and negative ion modes (Figure S2), indicating that DON treatment significantly altered the metabolic profile of 3D4/21 cells. Furthermore, we ranked 127 differential metabolites by log_2_FC. In positive ion mode, ophthalmic acid, lysyl-Hydroxyproline and γ-Glutamylcysteinylserine were upregulated, while (±)-2-Methylthiazolidine, tyrosyl-Asparagine, valyl-Threonine, L-proline, and racemethionine were downregulated (Table 2). In negative ion mode, nitazoxanide, 5-Hexenyl glucosinolate, 2-Deoxy-6-O-sulfo-2-(sulfoamino)-D-glucopyranose, hypotaurocyamine and nicotinate D-ribonucleoside were upregulated, whereas nicotinic acid mononucleotide, asparagusic acid syn-S-oxide and gemcitabine were downregulated (Table 3).
Enrichment analysis revealed that the differential metabolites were involved in eight metabolic pathways, including lysosomes, parkinsonism, taste transduction, purine metabolism, glutathione metabolism, arginine and proline metabolism, and ABC transporter proteins (Figure 4a). These pathways were classified under KEGG categories as follows: cellular processes (transport and catabolism); environmental information processing (signaling molecules and interaction; signal transduction; membrane transport); genetic information processing (translation); human diseases (neurodegenerative disease); metabolism (nucleotide metabolism; metabolism of terpenoids and polyketides; metabolism of other amino acids; metabolism of cofactors and vitamins; global and overview maps; energy metabolism; carbohydrate metabolism; amino acid metabolism); and organismal systems (sensory system) (Figure 4b).
3.5. Integration of Metabolomics Results with Transcriptomics
To provide mechanistic insight into links between gene expression and functional metabolic outcomes, we integrated our previous transcriptomic analysis of DON-induced 3D4/21 cells with current metabolomics data [17]. In the positive ion mode, specific correlation analyses (Figure S3) showed that the metabolite HMDB0039733 exhibited the strongest positive correlation with the gene PDIA4, with a correlation coefficient of 0.9994, while HMDB0039733 displayed the strongest negative correlation with the gene TXNIP, with a coefficient of −0.9989. Additionally, in the positive ion mode (Figure S4), the metabolite HMDB0125517 showed the strongest positive correlation with the gene OTUD1 (0.9977), and the metabolite CSID54753 had the strongest negative correlation with the gene BRCA1 (−0.9986). In addition, pathway enrichment analysis indicated that both differential genes and metabolites co-regulated purine metabolism, metabolic pathways, lysosome, glutathione metabolism, and arginine–proline metabolism (Figure 5).
4. Discussion
Deoxynivalenol (DON) is a well-known threat to porcine health and swine industry productivity because it damages macrophages. However, the global metabolomic perturbations underlying DON-induced immunotoxicity remain poorly understood. In this study, we employ non-targeted metabolomics, cell viability assays, and multi-omics integration to characterize the metabolic disturbances caused by DON in 3D4/21 cells. Our findings extend current knowledge of DON as a potent immunosuppressive mycotoxin in pigs.
The concentration- and time-dependent reduction in 3D4/21 cell viability corroborates our previous evidence of DON’s direct cytotoxic effects in 3D4/21 cells [17]. Consistent with our previous results, DON treatment led to a decrease in cell viability at 2 μM after 24 h exposure. Unexpectedly, a significant decrease also occurred at 1 µM after only 12 h, showing that DON exerts harmful effects even at low concentrations and that 3D4/21 cells are highly sensitive to it. In porcine alveolar macrophages (PAMs), 1 µM DON did not affect viability but did alter immune signaling [18]. Previous studies have shown that DON exerts variable effects depending on both the cell type and concentration. For example, in granulosa cells, low concentrations of DON have been reported to promote cell proliferation, potentially through enhanced IGF-I signaling, while higher concentrations inhibit proliferation, likely due to cytotoxic effects [19,20,21]. These findings underscore that the biological impact of DON is not only dose-dependent but also highly influenced by the cellular context, indicating that different cell types may respond to DON exposure via distinct mechanisms.
In our study, all samples are processed using a standardized methanol–water extraction protocol alongside QC samples. These QC samples clustered tightly in PCA plots, and all metabolites showed a CV below 30%, minimizing the likelihood of systematic errors such as ion suppression. Moreover, PCA and OPLS-DA analyses showed clear separation between control and DON-treated groups, indicating profound metabolic dysregulation [22]. The robust model fit (Q^2^ > 0.85) and absence of overfitting (permutation test results) validate the reliability of our metabolomic data, supporting the biological relevance of identified DMs.
DON drives oxidative stress management and rewires sulfur amino acid and NAD^+^ metabolism in porcine macrophages, metabolic axes closely tied to their inflammatory phenotype [23,24]. In positive ion mode, we observed elevated levels of ophthalmic acid, a sensitive proxy for glutathione depletion and oxidative stress [25], together with γ-glutamylcysteinylserine, a dipeptide that reinforces glutathione biosynthesis under inflammatory challenge [26]. Concomitant declines in racemethionine [27], L-Proline [28], and related dipeptides imply a constrained sulfur and aminoacid supply that supports macrophage methylation and stress-responsive metabolism. In negative ion mode, accumulation of nitazoxanide [29] points to an immunomodulatory signal, whereas the opposite shifts in nicotinate D-ribonucleoside (up) and nicotinic acid mononucleotide (down) reflect remodeling of the NAD salvage network that shapes macrophage effector programs [23]. Increased hypotaurocyamine [30] is consistent with taurine/hypotaurine antioxidative pathways, and the reduction in gemcitabine [31] aligns with lowered exposure to a nucleoside analog known for myeloid immunosuppression. Two high-abundance features were annotated as harzianopyridone, a pyridone alkaloid produced [32], and oxybutynin chloride, a compound with antimuscarinic activity [33]. As there is no evidence of endogenous synthesis of these compounds in macrophages or any mechanistic association with DON or PAMs, their observed increase is likely attributable to background contamination. This finding does not impact our conclusions regarding DON-induced immunotoxicity. These results emphasize the need for further validation to confirm compound identities and determine any residual biological relevance.
Through metabolic pathway analysis, we identified metabolic pathways associated with immune regulation. Glutathione (GSH) metabolism emerged as a critical pathway, with reduced GSH levels potentially compromising antioxidant defenses and exacerbating oxidative stress [34,35]. This aligns with DON’s known ability to induce reactive oxygen species (ROS) production and mitochondrial damage in porcine lymphocytes [10]. Depletion of GSH might disrupt redox balance, amplifying cellular injury and immune dysfunction [36]. Notably, our study reveals that disturbances in arginine and proline metabolism constitute another key pathway affected by DON exposure. Arginine is a critical substrate for both nitric oxide synthase (NOS) and polyamine synthesis—processes essential for macrophage effector functions [37]. Disruption of arginine metabolism by DON may therefore impair the production of immune-related proteins in 3D4/21 cells, potentially reducing their functional capacity. As alveolar macrophages, 3D4/21 cells serve as a primary defense against pulmonary pathogens [38,39]. Consequently, dysfunction in these cells due to altered arginine metabolism could weaken this critical immune barrier in pigs. In addition, perturbations in purine metabolism suggest impaired nucleotide synthesis and energy homeostasis, which may contribute to reduced cell viability and dysfunctional immune responses [40]. Elevated purine metabolites could reflect increased nucleic acid turnover due to cellular stress or apoptosis, which is consistent with extensive apoptosis observed in lymphocytes after DON exposure in pigs [41,42].
Integrated transcriptomic–metabolomic analysis highlighted joint enrichment of purine, glutathione, and arginine/proline pathways, implying that their interaction drives DON immunotoxicity. In piglet models, DON exposure causes thymic atrophy and lymphocyte apoptosis [43], which may be related to energy metabolism disorders (purine metabolism) and oxidative stress (glutathione metabolism) identified in this study. For example, GSH depletion can overproduce ROS and damage mitochondrial function [44], while purine metabolism disorders reduce ATP supply, leading to energy crisis in immune cells and accelerating apoptosis [45,46]. The metabolite HMDB0039733 (γ-glutamylcysteinylserine) correlated strongly with PDIA4 and inversely with TXNIP, supported by stringent criteria (|PCC| ≥ 0.5, adjusted p < 0.05) and consistent patterns across replicates. PDIA4, a protein disulfide isomerase involved in endoplasmic reticulum (ER) stress responses [47], and TXNIP, a thioredoxin-interacting protein that modulates redox balance [48]. Their strong correlations indicate that PDIA4 upregulation may counteract ER stress induced by DON [17], while TXNIP downregulation could attempt to relieve inhibition of thioredoxin, a key antioxidant enzyme, to compensate for glutathione depletion. OTUD1 and BRCA1 showed strong correlations with specific metabolites under stringent statistical thresholds. Functionally, OTUD1 is involved in deubiquitination [49], while BRCA1 plays a key role in DNA repair [50], supporting their potential involvement in DON-induced toxicity. However, the specific roles of these genes in response to DON require further validation.
5. Conclusions
DON exposure significantly impairs 3D4/21 cell line viability and induces metabolic reprogramming. Key affected pathways include purine metabolism, glutathione metabolism, and arginine–proline metabolism. These disruptions suggest oxidative stress, energy imbalance, and immune dysfunction. Integrated metabolomic and transcriptomic analyses confirm their central role in DON-induced immunotoxicity. The study offers novel insights into DON’s effects on immune cells and identifies potential metabolic targets for mitigation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Magnoli A.P. Poloni V.L. Cavaglieri L. Impact of mycotoxin contamination in the animal feed industry Curr. Opin. Food Sci.2019299910810.1016/j.cofs.2019.08.009 · doi ↗
- 2Gab-Allah M.A. Choi K. Kim B. Type B Trichothecenes in Cereal Grains and Their Products: Recent Advances on Occurrence, Toxicology, Analysis and Post-Harvest Decontamination Strategies Toxins 2023158510.3390/toxins 1502008536828399 PMC 9963506 · doi ↗ · pubmed ↗
- 3Wolf-Hall C.E. Hanna M.A. Bullerman L.B. Stability of Deoxynivalenol in Heat-Treated Foods J. Food Prot.19996296296410.4315/0362-028X-62.8.96210456755 · doi ↗ · pubmed ↗
- 4Jia B. Lin H. Yu S. Liu N. Yu D. Wu A. Mycotoxin deoxynivalenol-induced intestinal flora disorders, dysfunction and organ damage in broilers and pigs J. Hazard. Mater.202345113117210.1016/j.jhazmat.2023.13117236907058 · doi ↗ · pubmed ↗
- 5Serviento A.M. Brossard L. Renaudeau D. An acute challenge with a deoxynivalenol-contaminated diet has short- and long-term effects on performance and feeding behavior in finishing pigs J. Anim. Sci.2018965209522110.1093/jas/sky 37830423126 PMC 6276570 · doi ↗ · pubmed ↗
- 6Reddy K.E. Kim M. Kim K.H. Ji S.Y. Baek Y. Chun J.L. Jung H.J. Choe C. Lee H.J. Kim M. Effect of commercially purified deoxynivalenol and zearalenone mycotoxins on microbial diversity of pig cecum contents Anim. Biosci.20213424325510.5713/ajas.20.013732777887 PMC 7876725 · doi ↗ · pubmed ↗
- 7Panisson J.C. Wellington M.O. Bosompem M.A. Nagl V. Schwartz-Zimmermann H.E. Columbus D.A. Urinary and Serum Concentration of Deoxynivalenol (DON) and DON Metabolites as an Indicator of DON Contamination in Swine Diets Toxins 20231512010.3390/toxins 1502012036828434 PMC 9967145 · doi ↗ · pubmed ↗
- 8Pierron A. Kleber A. Mayer E. Gerner W. Effect of DON and ZEN and their metabolites DOM-1 and HZEN on B cell proliferation and antibody production Front. Immunol.202415133893710.3389/fimmu.2024.133893738449861 PMC 10915041 · doi ↗ · pubmed ↗