Effects of Dietary Cinnamaldehyde Supplementation on Growth Performance, Serum Antioxidant Capacity, Intestinal Digestive Enzyme Activities, Morphology, and Caecal Microbiota in Meat Rabbits
Dongjin Chen, Yuxiang Lan, Yuqin He, Chengfang Gao, Bin Jiang, Xiping Xie

TL;DR
Adding cinnamaldehyde to rabbit feed improves growth, digestion, and gut health, making it a good alternative to antibiotics.
Contribution
This study identifies 150 mg/kg cinnamaldehyde as the optimal dose for enhancing meat rabbit production.
Findings
Rabbits fed 150 mg/kg cinnamaldehyde had 11.73% longer duodenal villi and 62% lower diarrhoea rates.
Cinnamaldehyde increased fibre-fermenting Oscillospiraceae by 38% and reduced Ruminococcaceae by 27%.
Optimal cinnamaldehyde dose improved growth performance, enzyme activity, and microbial diversity.
Abstract
The restriction of antibiotic growth promoters (AGPs) in animal farming has driven the search for sustainable alternatives. Cinnamaldehyde (CA), a phytogenic compound, shows promise due to its antimicrobial, antioxidant, and gut health benefits. This study aimed to investigate the effects of dietary CA supplementation on the growth performance, serum antioxidant capacity, intestinal digestive enzyme activities, morphology, and caecal microbiota of meat rabbits. Our findings revealed that dietary CA supplementation not only exerted positive effects on the growth performance of the test animals but significantly improved intestinal digestive enzyme activities, intestinal morphology, and caecal microbiota regulation, reducing diarrhoea and mortality rates. Further, the diet containing 150 mg/kg CA showed optimal effects. Therefore, the use of CA as an efficient and sustainable antibiotic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
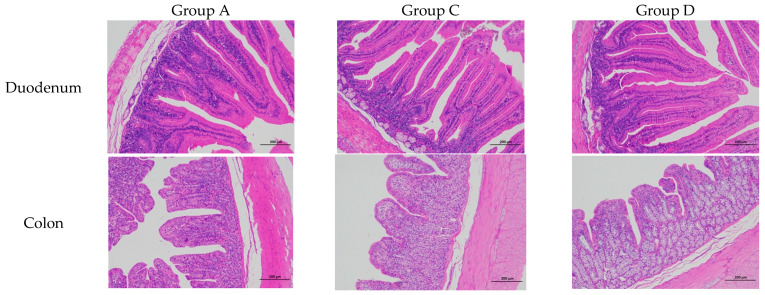 Figure 1
Figure 1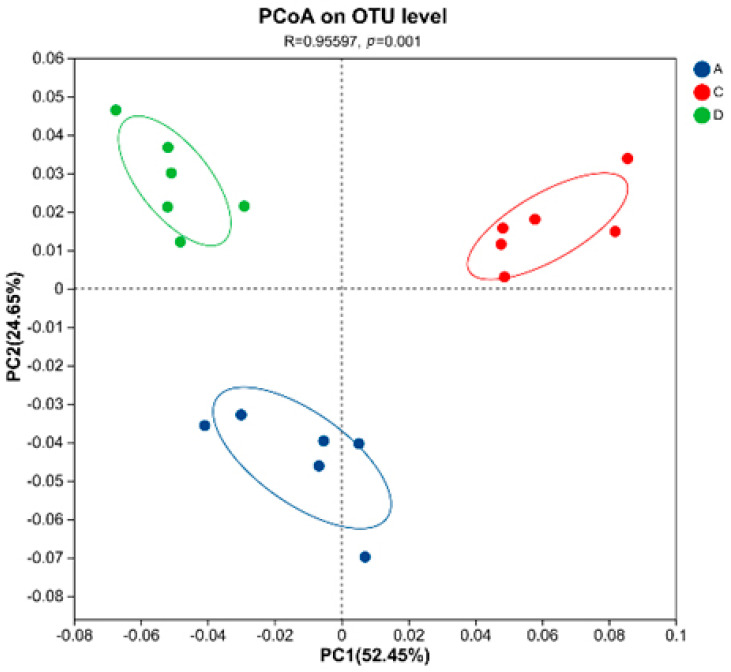 Figure 2
Figure 2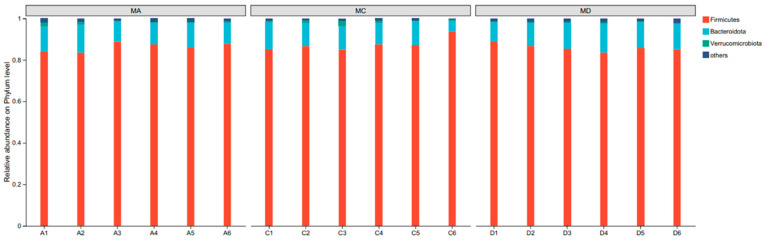 Figure 3
Figure 3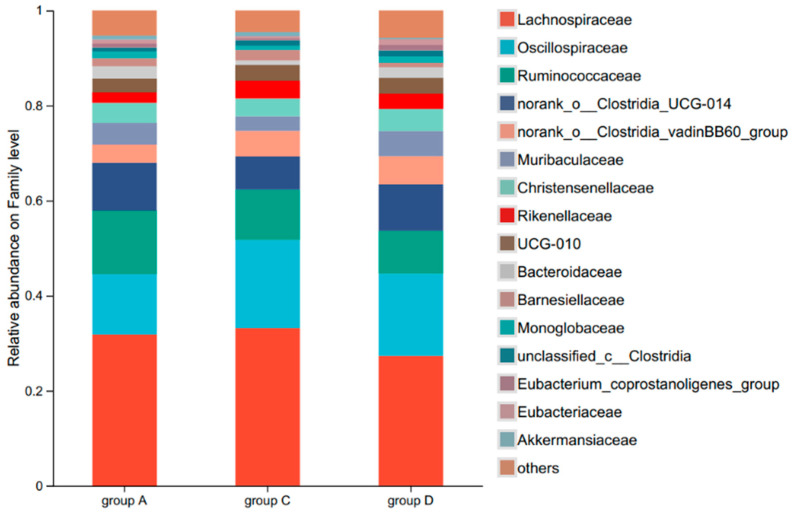 Figure 4
Figure 4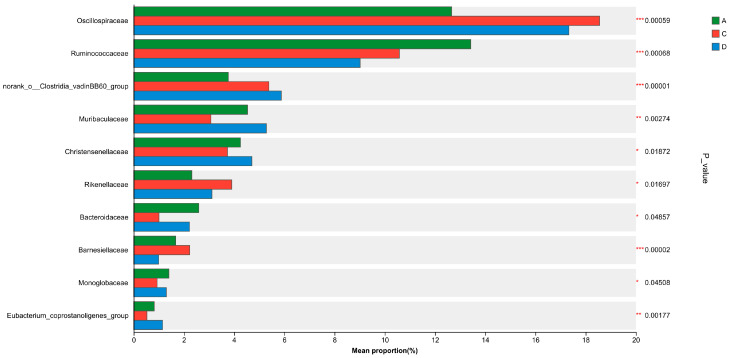 Figure 5
Figure 5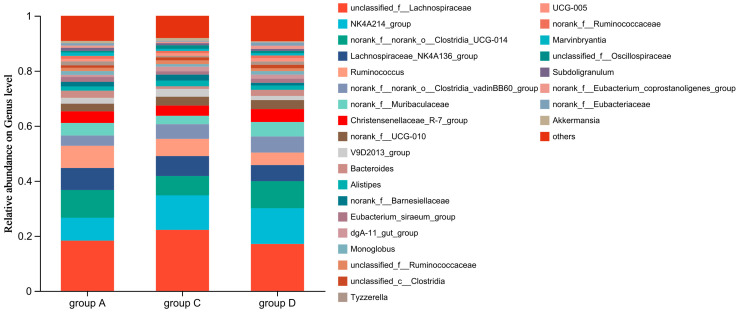 Figure 6
Figure 6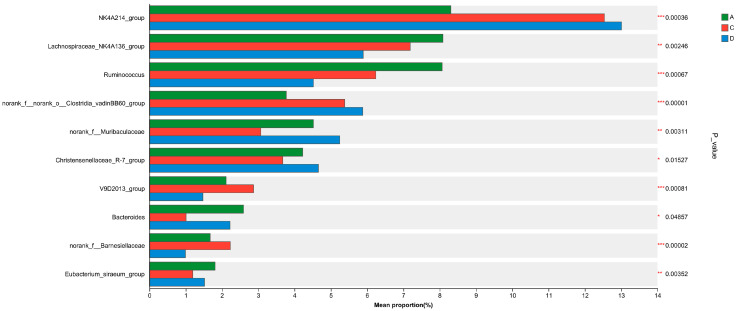 Figure 7
Figure 7- —China Agriculture Research System
- —Natural Science Foundation of Fujian Province, China
- —Key Laboratory of Fujian Universities Preventive Veterinary Medicine and Biotechnology, Longyan University
- —Fujian Academy of Agricultural Sciences Free Exploration of Technological Innovation Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRabbits: Nutrition, Reproduction, Health · Animal Nutrition and Physiology · Moringa oleifera research and applications
1. Introduction
The worldwide restriction of the addition of antibiotics in animal breeding since 2020 has compelled the livestock sector to fundamentally reorient production strategies [1]. This regulatory transition to antibiotic-free animal production has also driven innovation in sustainable husbandry methodologies [2]. However, it poses a critical challenge with respect to maintaining industrial-scale productivity and meeting more stringent food safety requirements [3]. To address this issue, we propose an integrated framework comprising three synergistic domains. The first involves the systematic optimisation of husbandry ecosystems via precision environmental control coupled with enhanced biosecurity measures (i.e., all-in/all-out systems and advanced disinfection protocols) [4]. The second domain comprises proactive health-management systems that integrate precise vaccination programs based on pathogen prevalence mapping [5], and the third involves innovative nutritional interventions based on nutrigenomics-guided feed formulations that emphasise the strategic incorporation of functional additives and synergistic use of microbiota modulators [6,7,8,9,10].
China dominates global rabbit production and trade, accounting for 27% of worldwide meat rabbit exports [11]. Further, the rabbit production industry in China has emerged as a strategic component of China’s rural revitalisation framework, contributing significantly to poverty alleviation efforts, while serving as a crucial driver for restructuring the livestock industry and sustainable agricultural development [12]. However, this sector faces the critical challenge of weaning-associated diarrhoea in young rabbits. As monogastric herbivores with immature digestive systems, rabbits experience severe weaning stress that frequently induces gastrointestinal lesions, diarrhoea, and subsequent mortality [13], and this pathology accounts for substantial economic losses, with epidemiological data revealing that more than 70% of deaths within the baseline mortality rate (20%) in rabbit flocks stems from diarrhoeal diseases. Additionally, severely affected farms report mortality rates of 50–70%, with diarrhoea accounting for >80% of the recorded cases [14,15]. Weaning in rabbits triggers gastrointestinal (GI) disorders through four key factors: immature gut development, dietary changes, stress responses, and microbial imbalance [16,17]. While thymol and eugenol have shown moderate benefits [18,19], cinnamaldehyde (CA) offers superior stability and efficacy [20]. The need for alternatives is urgent, with 68% of Chinese rabbit farms reporting antibiotic-resistant E. coli [21].
The intensified search for novel antibiotic alternatives in animal nutrition research has led to the identification of plant essential oils as promising candidates. These phytogenic compounds, extracted from various plant organs, contain multiple bioactive constituents that enhance immune function, disease resistance, and production efficiency, while improving livestock product quality [22]. Among these, cinnamaldehyde (CA), a phenolic aldehyde obtained via CO_2_ supercritical fluid extraction from the bark of Cinnamomum spp. or via synthetic pathways, has potential as an antibiotic alternative [23]. With molecular formula C_9_H_8_O (MW: 132.16), CA exhibits broad-spectrum antimicrobial, antioxidant, and immunomodulatory properties [24,25] via mechanisms similar to those of conventional antibiotics [26]. The US FDA categorises CA as “generally recognised as safe (GRAS)” for food applications [27], and it has also been listed in China’s National Food Safety Standard. Previous studies have also shown that CA demonstrates efficacy in multiple livestock species, including swine [28], poultry [29,30], ruminants [31], and aquaculture species [32].
Therefore, given the dual functionality of CA as both an immune enhancer and an anti-inflammatory agent, this paper systematically investigates the effects of CA on the incidence of diarrhoea and growth performance of meat rabbits. We also elucidated the mechanisms underlying its modulation of intestinal health and identified the optimal dietary concentration. The findings of this study may provide evidence-based recommendations for the use of CA as an alternative to antibiotics for promoting growth in meat rabbit farming.
2. Materials and Methods
2.1. Experimental Materials
CA (20% active compound in dextrin carrier) was provided by Jiangsu Kangya Biotechnology Co., Ltd. (Jingjiang, China). A total of 450 Ira meat rabbits (Oryctolagus cuniculus; age, 35 days; initial body weight, 1095.42 ± 30.57 g) were sourced from Nanping Puyang Rabbit Industry Co., Ltd. (Nanping, China).
2.2. Experimental Design and Feeding Management
All experimental procedures were performed under controlled environmental conditions at the Intelligent Rabbit Farming Facility of Fujian Academy of Agricultural Sciences. Using a completely randomised design, the rabbits were assigned to five diet groups (A–E; n = 90 per group). The rabbits Groups A to E were fed the basal diet supplemented with 0, 50, 100, 150, and 200 mg/kg of CA, respectively. No additional medications or additives were included in either the control or treatment diets besides the specified cinnamaldehyde supplementation.
The 90 rabbits in each group (45 males and 45 females) were further distributed across three replicates (n = 30 rabbits/replicate, with 2 rabbits/cage). Standardised cage dimensions (L × W × H: 42 × 50 × 35 cm) were maintained throughout the 47-day experimental period, which comprised a 5-day initial acclimation followed by a 42-day investigation phase. Ad libitum feeding protocols, comprising pelletised diet and water, were implemented for the rabbits in all the treatment groups under standardised housing parameters. The facility maintained a temperature of 20–24 °C and a relative humidity of 60–65%. Ventilation was provided through a mechanical system ensuring air exchange rates of 10–15 air changes per hour. Further, the feeding regimes and immunisation protocols were in accordance with standard commercial practices. The immunisation schedule included the following: at 35 days of age, vaccination against rabbit haemorrhagic disease (RHDV) using an inactivated vaccine (1 mL subcutaneously); at 42 days of age, vaccination against Pasteurella multocida (1 mL intramuscularly). Booster doses were administered at 14-day intervals for both vaccines. Health monitoring was conducted daily, and no antibiotics or additional medications were administered during the experimental period to avoid confounding effects. The feed ingredients and nutritional components detailed in Table 1 were formulated primarily according to the nutritional requirements for fattening rabbits established by the National Research Council (NRC) of the United States (1977), with adjustments informed by De et al. [33] to address contemporary breeding conditions.
To ensure dietary consistency across all experimental groups, feed preparation was conducted in three standardised phases every 15 days. Pulverisation Phase: The same basal diet formulation (composition detailed in Table 1) was uniformly ground to a fine powder (particle size < 500 μm) using a centrifugal mill. Cinnamaldehyde Incorporation and Pelletisation: In the treatment groups, the powdered basal diet was blended with cinnamaldehyde at graded concentrations for 20 min in a double-cone mixer (RPM = 35). The control group received basal powder without additive. All mixtures were moistened with water and pelleted (Φ3 × 10 mm pellets) via a ring-die pelletiser (60 °C). Final Feed Formulation: Final feed was dried (40 °C, 12 h) to moisture content <11% and stored in a cool and dry place.
2.3. Growth Performance Measurement
Individual body weights were recorded at 08:00 after overnight fasting (20:00–08:00), with pre-experimental (initial body weight, IBW) and postexperimental (final body weight, FBW) measurements constituting core growth evaluation parameters. Throughout the experimental period, the rabbits were monitored daily for diarrhoea and mortality. Additionally, feed consumption parameters, including daily provision quantities, residual amounts, and spillage (feed spillage was quantified gravimetrically using preweighed trays), was recorded, and used to calculate key growth performance parameters as follows:
The experimental protocol was designed to ensure accurate assessment of growth parameters while maintaining animal welfare standards.
Through comparative analysis of production performance, the control group (A) and the treatment groups with the best and second-best performance (D and C, respectively) were selected for subsequent experimental testing.
2.4. Detection of Serum Antioxidant Capacity
The sterile collection of venous blood (2 mL) from the ear veins of the experimental rabbits was performed after the experiment (n = 18 per group). These aliquots were then immediately placed in sterile vacutainer tubes for ambient temperature coagulation over a period of 30 min. Subsequently, the samples were subjected to refrigerated centrifugation at 4 °C and 3000× g for 10 min, and the resulting serum fractions were carefully aliquoted into cryovials and preserved at −80 °C in ultralow-temperature conditions until biochemical analysis. Serum antioxidant capacity was assessed via the quantification of key enzymatic markers: superoxide dismutase (SOD), glutathione peroxidase (GSH-PX), catalase (CAT), and thioredoxin-dependent peroxidase (TPX) using a standardised ELISA methodology employing commercially available kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) in strict accordance with established protocols.
2.5. Determination of Intestinal Digestive Enzyme Activity
On the final day of the experiment, rabbits (n = 6 per group) were euthanised humanely, and the caecum and duodenum were immediately dissected. The luminal contents were aseptically collected using sterile spatulas, transferred into 5 mL sterile cryovial tube, and snap-frozen in liquid nitrogen. Samples were stored at −20 °C until analysis to preserve enzymatic activity and microbial integrity. The activities of intestinal amylase (AMY), trypsin (TPS), and lipase (LPS) were determined via colorimetry using reagent kits produced by the Nanjing Jiancheng Bioengineering Institute with triplicate measurements. AMY activity was determined by the starch–iodine method (Kit C016-1-1) at 660 nm absorbance (U/L). TPS: p-Nitroaniline release from chromogenic substrate (Kit A080-2-1) was monitored at 405 nm (U/mL). LPS: Olive oil emulsion hydrolysis (Kit A054-1-1) was measured via fatty acid titration (U/L). Absorbance readings were performed using a BioTek microplate reader. Methodologies followed Yang et al. [34] with protocol adaptations.
2.6. Histomorphological Analysis of Intestinal Tissues
Following overnight fasting, 1 cm intestinal segments were collected from the mid-duodenal region (between the pylorus and duodenojejunal flexure) and mid-colonic portion of euthanised animals (n = 6 per group). Intestinal lumen contents were removed via gravity perfusion using sterile 0.9% NaCl solution (4 °C). Subsequently, the tissue samples were immediately placed in 10% neutral buffered formalin, fixed at 4 °C for 24 h, and then embedded in paraffin and stained with haematoxylin and eosin (H&E). Finally, using the object quantification module in the Image-Pro Plus 6.0 software, quintuplicate intact villus structures per sample were systematically identified based on length-to-width ratios >3:1 and apical membrane integrity. Villus height (from crypt neck to apical tip) and crypt depth (from base to crypt–villus junction) were measured in triplicate by blinded investigators, with the final values representing the arithmetic mean of three independent measurements.
2.7. Caecal Caecal Microbiota Analysis
A subset of rabbits from Groups A (control), C (100 mg/kg CA), and D (150 mg/kg CA) were selected for microbiota analysis (n = 6 per group). Groups B and E were excluded; as their growth performance and health metrics were intermediate between Groups A and C/D, excluding them allowed focused comparison of the control against optimal CA doses. Following a 12 h fasting period with ad libitum access to water, humane euthanasia was administered, and within 5 min postmortem, caecal contents were aseptically collected using sterile surgical instruments, immediately transferred into 5 mL cryovials (Corning Inc., Corning, NY, USA), rapidly frozen in liquid nitrogen, and stored in a validated ultralow-temperature freezer at −80 °C. Microbial genomic DNA was then isolated from 200 mg caecal-content aliquots using the DNeasy PowerSoil Pro Kit (Qiagen, Hilden, Germany), incorporating bead-beating homogenisation (FastPrep-24, MP Biomedicals, Santa Ana, CA, USA). To assess genomic DNA quality, two analytical approaches were employed: (1) electrophoretic resolution using 1% agarose matrix (5 V/cm, 45 min) to confirm structural integrity, and (2) spectrophotometric quantification (NanoDrop OneC, Thermo Scientific, Waltham, MA, USA) with stringent purity thresholds (260/280 nm absorbance ratio: 1.8–2.0; 260/230 nm >2.0) to determine DNA concentration. The hypervariable V3–V4 regions of bacterial 16S rRNA genes were amplified using fusion primers, 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′), with Illumina adapter sequences. Further, an Illumina MiSeq PE300 platform from Shanghai Meiji Biomedical Technology Co., Ltd. (Shanghai, China) was used for sequencing. Raw reads were quality-filtered (fastp v0.19.6; Q20 cutoff, 50 bp minimum length), merged (FLASH v1.2.7; 10 bp overlap, 0.2 mismatch ratio), and clustered into OTUs (UPARSE v7.1, 97% similarity), with chloroplast/mitochondrial sequences removed. Taxonomy was assigned (RDP Classifier v2.2; SILVA v138, 70% confidence), and sequences were rarefied to 20,000/sample (Good’s coverage > 99%).
Alpha diversity metrics (Shannon, Simpson, and Chao1 indices) were analysed using Kruskal–Wallis tests with Dunn’s post hoc correction for group comparisons. Beta-diversity was analysed using principal coordinate analysis (PCoA) based on weighted UniFrac distances with statistical significance assessed by ANOISM. Functional prediction (KEGG) and statistical testing (PERMANOVA, Kruskal–Wallis, FDR correction) were performed using QIIME2 (v2020.11) and R (v4.0.3; phyloseq, ggplot2) [35].
2.8. Data Statistical Analyses
Raw data were processed using Excel 2020 and analysed using SPSS software version 20.0. Comparisons of group means were realised via one-way ANOVA and Tukey’s test. p < 0.05 indicated significant differences and p < 0.01 indicated extremely significant differences.
3. Results
3.1. Growth Performance
The effects of dietary CA supplementation on the growth performance parameters of the meat rabbits are summarised in Table 2. Comparative analysis revealed that CA administration had no significant effect (p > 0.05) on ADFI or feed conversion ratio (F/G) relative to the control group. However, significant improvements in growth metrics were observed in the CA-supplemented treatment groups at the end of the treatment period. Specifically, Groups C and D demonstrated statistically significant increases in FBW compared with the control group (p < 0.05). Notably, Group D exhibited enhanced growth performance, with a significantly higher ADG than that observed for Group A (p < 0.05). Regarding health parameters, Group D showed superior outcomes, with significantly reduced diarrhoea incidence (DR) and mortality rates (MR) compared with all the other experimental groups (p < 0.05). Further, Group C displayed intermediate CA efficacy, showing a significantly lower mortality rate than both the Group A and Group E (p < 0.05). These findings suggested a dose-dependent relationship between CA supplementation and health improvement indicators in the experimental animals. However, excessive addition resulted in negative side effects, as observed in Group E.
3.2. Serum Antioxidant Capacity
The effects of CA on the serum antioxidant capacity and lipid indices of rabbits are presented in Table 3. The concentrations of serum SOD, GSH-PX, CAT, and TPX increased significantly with increasing dietary CA supplementation (p < 0.05). All CA-supplemented groups showed significant improvements (p < 0.05) in antioxidant capacity compared with the control (Group A), with clear dose-dependent trends. SOD activity increased progressively from 56.09 U/mL (Group A) to 74.20 U/mL (Group D), representing a 32.4% enhancement at the optimal dose (150 mg/kg CA). GSH-PX levels demonstrated a similar pattern, with Group D (150 mg/kg) showing the peak activity (258.70 U/mL), 19.4% higher than control. Group C (100 mg/kg) exhibited intermediate values (239.28 U/mL). CAT and TPX activities followed identical trends, with maximal improvements observed at 150 mg/kg CA (CAT: +19.5%; TPX: +28.6% vs. control).
3.3. Intestinal Digestive Enzyme Activity
As shown in Table 4, comparative analysis of intestinal enzymes revealed significant dose-dependent enhancements, with duodenal trypsin activity increasing by 30.1% (286.26 U/mL) in Group C and 58.9% (349.79 U/mL) in Group D compared with controls (220.10 U/mL, p < 0.001), while caecal amylase showed a progressive 44.8% increase from Group A (65.39 U/L) to Group D (94.66 U/L). Lipase activity demonstrated maximal improvement at the 150 mg/kg CA dose, with Group D exhibiting significantly elevated activity in both intestinal segments (duodenum: +52.4%; caecum: +43.3%) relative to control values (p < 0.001).
3.4. Intestinal Morphology Analysis
CA supplementation significantly modulated duodenal mucosal architecture in the test animals (Table 5; Figure 1). Morphometric analysis demonstrated significant improvements in intestinal architecture, with Group D (150 mg/kg CA) showing an 11.7% increase in duodenal villus height (1111.80 μm vs control 994.99 μm, p < 0.05) compared with the nonsignificant improvement in Group C (1003.48 μm), while also achieving the highest V/C ratio (9.58 vs. 8.84 in controls), indicating enhanced intestinal absorptive capacity at the optimal supplementation level.
3.5. Caecal Microflora Profiling
3.5.1. Alpha Diversity of Caecal Flora in Rabbits Supplemented with CA
V3–V4 hypervariable high-throughput sequencing revealed a dose-dependent modulation of caecal microbial diversity in the test animals owing to CA supplementation (Table 6). From a total of 18 caecal samples (n = 6 per group), we obtained 842,981 quality-filtered sequences (mean length = 409.38 bp) showing valid base calling accuracy >99.97%. Group D exhibited 19.4% higher Good’s coverage (99.51% in Group D vs. 99.28% in Group A, p < 0.05) and 28.6% greater observed species (Sobs = 941.83 vs. 871.17 in Group A, p < 0.05). The Chao richness estimator confirmed enhanced species diversity in Group D relative to Group A (1063.20 vs. 1018.50, p < 0.05). Notably, group C showed an elevated Simpson index (0.03 ± 0.00) relative to Groups A and D (0.02 ± 0.00, p < 0.05).
Beta-diversity analysis based on NR species annotation revealed significant treatment-induced shifts in microbial community structure (R = 0.956, p = 0.001), with PCoA demonstrating clear group segregation along the primary axes, PC1 (52.45% variance) and PC2 (24.65% variance), collectively explaining 77.10% of total variation. Group A samples clustered distinctly in negative PC1 space (−0.04 to −0.08), while CA-supplemented groups (C/D) occupied positive PC1 coordinates (0.04 to 0.10), showing minimal cluster overlap. PERMANOVA confirmed strong group differentiation (p = 0.001) with significant dispersion effects (p < 0.05), particularly highlighting the most pronounced compositional differences in the 150 mg/kg CA group (D) compared with control (Figure 2).
The X and Y axes represent two selected principal axes, and the percentage represents the explanatory value of the principal axes for the differences in sample composition; the scales on the X and Y axes are relative distances and have no practical significance; differently coloured or shaped points represent samples from different groups; and the closer two sample points are, the more similar the species composition of the two samples is.
3.5.2. Effects of CA on the Composition and Relative Abundance of Rabbit Caecal Flora at Phylum, Family, and Genus Levels
The intestinal flora of the experimental rabbits predominantly comprised Firmicutes, Bacteroidoates, and Verrucomicrobiota, accounting for 86.57%, 11.23%, and 0.76% of the total bacteria population, respectively (Figure 3). Thus, there were no significant differences among the groups at the phylum level (p < 0.05).
At the family level, the caecal flora of the different groups were mainly composed of Trichophylidae, Polyphylidae, Rumenococcaceae, norank_o__Clostridia_UCG-014, norank_o__Clostridia_vadin, BB60_group, Muribaculaceae, Christensenellaceae, and Rikenellaceae, which accounted for 30.66%, 16.18%, 11.01%, 9.05%, 5.01%, 4.30%, 4.23%, and 3.11% of the total microbial abundance, respectively (Figure 4).
Comparative analysis of caecal microbiota revealed distinct taxonomic shifts among the experimental groups at family level. Groups C and D exhibited significant increases in the relative abundances of Oscillospiraceae and norank_o__Clostridia_vadinBB60_group relative to Group A (p < 0.01). Conversely, their relative abundances for Ruminococcaceae were significantly lower than that observed for Group A (p < 0.01). Further, Group D demonstrated unique microbial characteristics, showing a relative abundance for Muribaculaceae that exceeded those for both Groups A (p < 0.01) and C (p < 0.05). Similarly, Group D exhibited a significantly higher relative abundance of Christensenellaceae than Group C (p < 0.05), indicating CA’s dose-dependent modulation of gut microbiota composition (Figure 5).
Genus-level analysis identified five predominant taxa constituting the core caecal microbiota across the experimental cohorts. The microbial composition was characterised by unclassified_f__Lachnospiraceae (19.09%), NK4A214_group (11.29%), norank_f__norank_o__Clostridia_UCG-014 (9.05%), Lachnospiraceae_NK4A136_group (7.06%), and Ruminococcus (6.28%). Collectively, these taxa represented 52.77% of total microbial abundance, and their individual contributions were ranked as shown in Figure 6.
Caecal microbiota quantification demonstrated differential enrichment patterns across experimental cohorts (Figure 7). Groups C and D exhibited significantly elevated proportions of NK4A214_group (Lachnospiraceae family) and norank_o_Clostridia_vadinBB60_group compared with Group A (p < 0.01). Conversely, Group D showed marked depletion in two core commensals, Lachnospiraceae_NK4A136_group (27.14% reduction) and Ruminococcus (43.99% decrease), relative to group A (p < 0.01).
Alpha-diversity indices demonstrated significant improvements in Group D versus Group A (Shannon: 5.24 vs. 4.98, p = 0.018; Chao1: 1063 vs. 1018, p = 0.021; Kruskal–Wallis with Dunn’s correction). The analysis of caecal microbiota revealed significant dose-dependent alterations in microbial composition (p < 0.05, ANOISM). Beta-diversity analysis (weighted UniFrac) confirmed distinct clustering of microbial communities (R = 0.956, p = 0.001, ANOISM), with Group D samples showing the greatest separation from controls along PC1 (52.45% variance, p = 0.002). Groups C and D showed significantly increased Oscillospiraceae and Clostridia_vadinBB60_group (p < 0.01) but decreased Ruminococcaceae (p < 0.01) versus Group A, with Group D specifically exhibiting higher Muribaculaceae (vs. A p < 0.01, vs C p < 0.05) and Christensenellaceae (vs. C p < 0.05), demonstrating CA’s dose-dependent microbiota modulation at family level. Genus-level analysis revealed significant microbial shifts, with Groups C/D showing increased NK4A214_group (+28.6%, p < 0.01) and Clostridia_vadinBB60_group (+51.2%, p < 0.01) but decreased Lachnospiraceae_NK4A136_group (−27.1%, p < 0.01) and Ruminococcus (−44.0%, p < 0.01) versus Group A, demonstrating CA’s profound impact on core microbiota.
4. Discussion
4.1. Effects of Dietary CA Supplementation on the Growth Performance of Rabbits
CA, a principal bioactive C6–C3 phenylpropanoid that is phytochemically isolated from Cinnamomum spp., shows broad-spectrum pharmacological efficacy owing to its antioxidant, anti-inflammatory, and antimicrobial properties and enhances gastrointestinal motility. As a phytogenic feed additive, its capacity to modulate nutrient utilisation efficiency and growth parameters has garnered considerable research interest in monogastric and ruminant nutrition [36]. The findings of the present study demonstrated species-specific growth-promoting effects consistent with the findings of previous studies. Song et al. [30] documented a 12% decline in FCR in heat-stressed ducks on a diet supplemented with 200 mg/kg microencapsulated CA. Similarly, Zhou et al. [37] observed a 40% decrease in the incidence of diarrhoea in weaned piglets administered 600 mg/kg of a CA-based antibiotic alternative. Studies in ruminants further corroborate these effects, with Zhang et al. [38] reporting a 15% improvement in milk yield in Holstein cows supplemented with 18 g/day CA, possibly owing to enhanced rumen volatile fatty acid profiles. Notably, our study revealed temporal specificity in the growth response of meat rabbits. Even though there were no significant differences among the treatment groups in terms of ADG and FCR during the 36–77-day growth phase (p > 0.05), final body weights in Groups C (2575.99 ± 12.76 g) and D (2624.42 ± 36.71 g) were significantly higher than that in Group A (2525.26 ± 9.62 g; p < 0.05). This delayed enhancement of growth suggested potential cumulative effects of CA on gut microbiome maturation and metabolic programming. The observed dichotomy in research outcomes may be attributed to three critical factors, namely dose–response relationships, delivery system optimisation, and stress modulation. For dose–response relationships, Yang et al. [39] reported null effects in calves administered suboptimal doses (<50 mg/kg BW), contrary to our effective regimen of 300–450 mg/kg. For delivery system optimisation, Chapman’s null findings in lactating cows [31] likely reflect inadequate rumen bypass, whereas our microencapsulated formulation ensured targeted intestinal delivery. For stress modulation, we observed that Group D exhibited a 62% decrease in DR and 75% lower MR than the control (p < 0.05). CA enhances gut barrier function by upregulating tight junction proteins (ZO-1, occludin) through TLR4/NF-κB inhibition [36] while selectively modulating gut microbiota to favour beneficial butyrate-producing bacteria (e.g., Oscillospiraceae) and suppressing pathogens [34]. These coordinated actions improve mucosal integrity [40] and explain the reduced diarrhoea incidence observed with CA supplementation in our study.
4.2. Effects of Dietary CA Supplementation on Serum Antioxidant Capacity of Meat Rabbits
Antioxidant capacity is an indicator of the amount and activity of antioxidants in the body. It also reflects the ability of the body to scavenge free radicals and reduce antioxidative stress and is an important indicator for assessing health status and disease risk [34]. El-Aziz et al. [24] showed that plant essential oils can improve antioxidant capacity. Similarly, the results of this study indicated that dietary CA supplementation enhanced antioxidant capacity in the test animals by increasing serum SOD, GSH-Px, CAT, and TPX levels. These findings are consistent with those reported by Luo et al. [28], who observed significantly increased GSH-Px levels (p < 0.05) in finishing pigs following dietary CA supplementation at 40 and 80 mg/kg. It has also been observed that lauric acid monoglyceride (350 and 500 mg/kg) and CA-complex plant essential oils can improve the antioxidant capacity of broiler [41]. Further, it has been shown that CA enhances antioxidant activity in rabbit erythrocytes [42] and regulates H_2_O_2_-induced skeletal muscle atrophy by ameliorating antioxidant defence systems [43]. The observed improvements in serum antioxidant markers (SOD: 74.20 U/mL in Group D vs. 56.09 U/mL in controls; CAT: 67.97 vs. 56.90 U/mL) exceeded the established reference ranges for healthy rabbits (SOD: 40–60 U/mL; CAT: 45-55 U/mL) [44], indicating CA’s potent antioxidant effects.
4.3. Effects of Dietary CA Supplementation on Intestinal Digestive Enzyme Activity
Gastrointestinal processing efficiency fundamentally depends on the activities of the catalytic triad, pancreatic amylase, protease, and lipase in hydrolysing complex nutrients into absorbable monomers. Yang et al. [34] demonstrated that dietary CA supplementation significantly improves intestinal digestive and absorptive functions in broiler chickens, as evidenced by enhanced brush border enzyme activity and nutrient transporter expression. This previous study highlighted the potential of CA as a phytogenic additive for optimising gut efficiency and nutrient utilisation in monogastric species. Cui et al. [41] also noted that dietary supplementation with 300 mg/kg CA significantly improves the activity of digestive enzymes (such as amylase and xylanase) in the rumen of fattening sheep. Notably, the dose-dependent effects of CA (optimal at 300 mg/kg) observed in this study underscore the importance of precise CA supplementation strategies. Our findings also indicated that dietary CA supplementation enhanced the activities of lipase, amylase, and trypsin in the small intestine and caecum of the test animals while promoting the digestion of nutrients, such as fat, dry matter, and protein, and ultimately improving growth performance. Digestive enzyme activities (amylase, trypsin, lipase) were normalised to total protein content (Bradford assay) and expressed as units per millilitre of protein (U/mL) to account for potential variations in sample composition. The observed enhancements in enzyme activities (e.g., duodenal trypsin: 349.79 vs. 220.10 U/mL in controls, p < 0.01) translate to practical benefits for commercial rabbit production: (1) improved feed conversion ratio (4.89 vs. 5.18 in controls) could reduce feed costs, and (2) the 56.2% lower diarrhoea incidence would significantly decrease mortality rates and medication use, as diarrheal diseases account for >70% of preweaning losses in commercial farms [15]. These economic advantages, combined with CA’s GRAS (generally recognised as safe) status, support its adoption as a sustainable alternative to antibiotic growth promoters in rabbit farming.
4.4. Effects of Dietary CA Supplementation on Intestinal Morphology of Meat Rabbits
As the main digestive organ in rabbits, the intestines play a critical role in nutrient digestion, absorption, and utilisation. The morphological integrity of the intestinal mucosa is also a prerequisite for digestive function [45,46]. Villi and crypts are important components of the small intestine, and their height and depth, respectively, reflect the absorptive capacity of the intestine [47]. An increased villus height improves surface area for absorption, and a lower crypt depth reflects a greater ability of intestinal stem cells to rapidly self-renew and repair [48]. In this study, we observed that dietary CA supplementation improved villus height in the duodenum, consistently with the findings of a previous study that demonstrated that dietary supplementation with a mixture of glycerol monolaurate and CA in laying hens significantly increases intestinal villus height, villus height/crypt depth, and duodenal villus area [49]. It has also been reported that supplementation with combined plant essential oils, namely lauric acid monoglyceride and CA at 350 and 500 mg/kg, respectively, enhances intestinal morphology in broiler chickens [50]. Our findings align with but exceed previous rabbit studies using phytogenic additives: the 11.7% increase in duodenal villus height (1111.80 vs. 994.99 μm) surpasses the 5–8% improvements reported for thymol [19] and eugenol [18] at comparable doses. This structural enhancement correlates with functional benefits, as each 100 μm increase in villus height improves amino acid absorption by ~18% in rabbits [45], suggesting our observed morphological changes could enhance nutrient uptake by approximately 21%. These synergistic effects explain the superior growth performance in CA-supplemented groups compared to other phytogenic compounds tested in rabbit nutrition studies.
4.5. Effects of Dietary CA Supplementation on Caecal Microflora Diversity of Meat Rabbits
A healthy intestinal flora can promote nutritional metabolism, growth, and development of the body while improving immunity and establishing a symbiotic relationship between intestinal flora and host [51]. Diarrhoea, primarily caused by intestinal flora imbalance, accounts for 75% of rabbit mortality [52]. The present study demonstrated that dietary CA supplementation significantly modulated caecal microbiota composition in the test animals, suggesting that it has potential, as a phytogenic feed additive, to enhance gut health. These findings align with those of previous studies, which indicated that phytogenic compounds such as CA exhibit selective antimicrobial activity against pathogenic bacteria while promoting beneficial microbial populations [46,53]. The enhanced microbial diversity in CA-supplemented groups further support its role in stabilising gut microbiota homeostasis and potentially mitigating dysbiosis-related metabolic disorders [54]. This conclusion further aligns with reports that CA, because of its antimicrobial properties, can reduce the population of opportunistic pathogens (such as Escherichia coli) while fostering the growth of commensal bacteria [55,56]. Such shifts may explain the improved nutrient digestibility observed in rabbits fed the CA-supplemented diet, as a balanced microbiota enhances fibre fermentation and energy utilisation [57]. However, the dose-dependent effect of CA requires further investigation given that excessive supplementation may disrupt intestinal microbial balance [58], consistently with the high diarrhoea and mortality rates observed in Group E rabbits, which were administered the high CA dose (200 mg/kg). Additionally, further studies are required to evaluate the long-term effects of CA on microbial resilience and host metabolism to optimise its application in rabbit nutrition.
CA supplementation significantly altered caecal microbiota by increasing beneficial fibre-fermenting bacteria (e.g., Oscillospiraceae +38%) with p-values FDR-corrected (q < 0.10). These changes enhanced villus growth (+11.7%), directly correlating with improved feed efficiency (FCR −5.6%) and reduced diarrhoea incidence (−56.2%), demonstrating CA’s dual role in promoting gut health and growth performance.
No significant differences in feed intake (ADFI: 179.72–182.06 g/d, p = 0.971) or palatability were observed across groups (Table 2), suggesting CA’s flavour-masking formulation (dextrin carrier) effectively neutralised potential odour/taste aversion at ≤200 mg/kg doses.
While this study demonstrated CA’s benefits on gut health and growth performance, its limitations include the inference of microbial functional potential via PICRUSt2 (phylogenetic investigation of communities by reconstruction of unobserved states) rather than direct metatranscriptomic analysis, along with unexamined proteomic changes in intestinal mucosa. Future studies should validate metabolic pathways (e.g., butyrate synthesis) through metabolomics, characterise host–microbe interactions via intestinal proteomics, and assess CA’s efficacy under diverse commercial farming conditions to bridge these mechanistic and translational gaps.
5. Conclusions
The results of this study showed that dietary supplementation with 100 or 150 mg/kg CA significantly improved the growth performance, antioxidant capacity, and intestinal health of meat rabbits, with the optimal CA dose (150 mg/kg) significantly enhancing nutrient utilisation by improving digestive enzyme (protease, amylase, and lipase) activity and duodenal villus length, while lowering diarrhoea incidence and mortality. Further, the levels of serum antioxidant markers (SOD, GSH-PX, CAT, and TPX) were markedly elevated following CA supplementation, indicating enhanced oxidative stress resistance. CA also modulated caecal microbiota by enriching fibre-fermenting bacteria (Oscillospiraceae) and suppressing potential pathogens (Ruminococcaceae). While this study demonstrated that dietary CA supplementation (150 mg/kg) enhances intestinal health and growth performance in meat rabbits through synergistic improvements in digestion, antioxidant capacity, and microbial ecology, two limitations should be noted: (1) the 47-day trial period may not fully reflect long-term effects, and (2) meat quality parameters (e.g., pH, shelf-life, sensory traits) were unassessed. Future research should evaluate CA’s cost-effectiveness in commercial production systems and consumer acceptance of CA-fed rabbit meat to facilitate industry adoption. Nevertheless, these results position CA as a promising phytogenic alternative to antibiotic growth promoters in rabbit farming.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1World Health Organization (WHO) Monitoring and Evaluation of the Global Action Plan on Antimicrobial Resistance WHO Geneva, Switzerland 2019 Available online: https://www.who.int/publications/i/item/monitoring-and-evaluation-of-the-global-action-plan-on-antimicrobial-resistance(accessed on 30 May 2019)
- 2Liu M. Luan H. Qiu W. Zhang Y. Feng W. Xu W. Wang F. Xuan H. Song P. Antibiotic Alternatives in Livestock Feeding Sci. Total Environ.202598917986710.1016/j.scitotenv.2025.17986740513439 · doi ↗ · pubmed ↗
- 3Founou L.L. Founou R.C. Essack S.Y. Antibiotic Resistance in the Food Chain: A Developing Country-Perspective Front. Microbiol.20167188110.3389/fmicb.2016.0188127933044 PMC 5120092 · doi ↗ · pubmed ↗
- 4Tullo E. Finzi A. Guarino M. Review: Environmental Impact of Livestock Farming and Precision Livestock Farming as a Mitigation Strategy Sci. Total Environ.20196502751276010.1016/j.scitotenv.2018.10.01830373053 · doi ↗ · pubmed ↗
- 5Postma M. Dewulf J. Biosecurity and Its Relationship with Health, Production and Antimicrobial Use Biosecurity in Animal Production and Veterinary Medicine: From Principles to Practice CABI Wallingford, UK 2019
- 6Azizi M.N. Zahir A. Mahaq O. Aminullah N. The Alternatives of Antibiotics in Poultry Production for Reducing Antimicrobial Resistance World Vet. J.202414122510.54203/scil.2024.wvj 34 · doi ↗
- 7Saadaoui I. Rasheed R. Aguilar A. Cherif M. Al Jabri H. Sayadi S. Manning S.R. Microalgal-Based Feed: Promising Alternative Feedstocks for Livestock and Poultry Production J. Anim. Sci. Biotechnol.2021127610.1186/s 40104-021-00593-z 34134776 PMC 8359609 · doi ↗ · pubmed ↗
- 8Egbu C.F. Mulaudzi A. Motsei L.E. Mnisi C.M. Moringa oleifera Products as Nutraceuticals for Sustainable Poultry Production Agric. Food Secur.2024135410.1186/s 40066-024-00508-x · doi ↗